TRPM7, Magnesium, and Signaling


Institute of Cardiovascular and Medical Sciences, BHF Glasgow Cardiovascular Centre, University of Glasgow, Glasgow G12 8TA, UK
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(8), 1877; https://doi.org/10.3390/ijms20081877
Submission received: 18 March 2019
/
Revised: 12 April 2019
/
Accepted: 12 April 2019
/
Published: 16 April 2019
(This article belongs to the Special Issue Magnesium in Differentiation and Development)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The transient receptor potential melastatin-subfamily member 7 (TRPM7) is a ubiquitously expressed chanzyme that possesses an ion channel permeable to the divalent cations Mg2+, Ca2+, and Zn2+, and an α-kinase that phosphorylates downstream substrates. TRPM7 and its homologue TRPM6 have been implicated in a variety of cellular functions and is critically associated with intracellular signaling, including receptor tyrosine kinase (RTK)-mediated pathways. Emerging evidence indicates that growth factors, such as EGF and VEGF, signal through their RTKs, which regulate activity of TRPM6 and TRPM7. TRPM6 is primarily an epithelial-associated channel, while TRPM7 is more ubiquitous. In this review we focus on TRPM7 and its association with growth factors, RTKs, and downstream kinase signaling. We also highlight how interplay between TRPM7, Mg2+ and signaling kinases influences cell function in physiological and pathological conditions, such as cancer and preeclampsia.
1. Introduction
The transient receptor potential melastatin 7 (TRPM7) and its close homologue TRPM6 share the unique feature of a cation channel fused to a C-terminal α-kinase domain [1,2,3]. TRPM6 and TRPM7 are present in a tetrameric form and each subunit comprises six transmembrane segments (S1-6), and a channel pore permeable to Mg2+, Ca2+, and Zn2+, located between segments 5 and 6 (Figure 1) [4,5]. TRPM7 is ubiquitously expressed, whereas TRPM6 is mostly expressed in cells from kidneys and intestines. In spite of their similarities, they are not compensated by each other, indicating that they are not redundant. TRPM7 was initially proposed to regulate, and itself to be regulated by, intracellular Mg2+ levels, with Mg-ATP below 1mM strongly activating the channel [6]. The importance of TRPM7 in cellular Mg2+ homeostasis has been investigated in several cell types, including leukocytes, platelets, vascular smooth muscle cells (VSMCs), cardiomyocytes, cardiac fibroblasts, osteoblasts, and tumor cells, in both physiological and pathological conditions [6,7,8,9,10,11,12,13]. Experiments using TRPM7 overexpressing cells demonstrated important mechanisms underlying the role of TRPM7 in Mg2+ homeostasis, and this was confirmed in the recently described crystal structure of mouse TRPM7, where partially hydrated Mg2+ ions occupy the center of the conduction pore [3]. However, the importance of TRPM7 in Mg2+ homeostasis has also been questioned by some studies, since the deletion of TRPM7 in T cells did not affect acute uptake or the maintenance of total cellular Mg2+ [14]. However, this may relate to the cell type studied, because Mg2+ is typically regulated by MagT1 in immune cells.
TRPM6 and TRPM7 have been linked to many signaling pathways. Emerging evidence indicates an important role for these systems in growth factor signaling through receptor tyrosine kinases (RTK). The C-terminal α-kinase domain induces phosphorylation of many downstream targets involved in RTK signaling including annexin A1, calpain II, myosin II, PLCγ2, Src, and SMAD2 [15], and regulates epigenetic modifications [16]. TRPM7 kinase domain may also influence TRPM7 channel function, although the exact mechanisms remain unclear [17,18,19].
In addition to controlling Mg2+ transport across cell membranes, TRPM7, together with other Ca2+ channels, regulates Ca2+ influx in many cell types [20,21]. In angiotensin II (Ang II)-stimulated cardiac fibroblasts, TRPM7 is functionally active and controls both Mg2+ and Ca2+ influx [9]. In neuroblastoma cells, bradykinin-induced activation of TRPM7 mediates Ca2+ influx in a kinase-independent manner [22] and Ca2+ entry in macrophages triggered by lipopolysaccharide (LPS) is controlled by TRPM7, as observed in a cell-specific deletion model using a Cre/Lox system [23]. The kinase domain of TRPM7 also plays an important role in Ca2+ homeostasis by modulating store-operated calcium channels (SOCE) [24,25].
TRPM7 is also permeable to Zn2+ and may be an important regulator of cellular Zn2+ dynamics. This was originally identified in mouse cortical neurons, where activation of TRPM7 channels increased intracellular Zn2+, and TRPM7 knockdown using a short hairpin RNA reduced TRPM7-like current and intracellular Zn2+ concentration [26]. Additionally, a recent study showed that TRPM7 is an intracellular Zn2+ storage vesicle, which sequesters Zn2+ during cytosolic overload, and releases Zn2+ under oxidizing conditions [27]. Interestingly, other channels from the TRPM family are also permeable to Zn2+, such as TRPM2, TRPM3, and TRPM5 [28,29,30].
Receptor tyrosine kinases (RTKs), through which growth factors signal, are membrane-associated receptors [31]. Upon growth factor binding, autophosphorylation of RTKs induces phosphorylation and activation of intracellular non-receptor tyrosine kinases that influence phosphorylation of downstream target proteins. The activation of RTKs is involved in critical signaling pathways and cell functions, including proliferation, differentiation, migration and contraction, processes also dependent on Mg2+ and Ca2+. The RTK family comprises major growth factor receptors, such as epidermal growth factor receptor (EGFR), vascular endothelial growth factor receptor (VEGFR), platelet-derived growth factor receptor (PDGFR), nerve growth factor receptor (NGFR), fibroblast growth factor receptor (FGFR), and insulin-like growth factor receptor (IGFR) [32].
In 2009, Bindels and colleagues demonstrated that EGF activates EGFR signaling and promotes TRPM6 translocation from the cytosol to the membrane in kidney cells, through Src family tyrosine kinases [33]. These effects were abolished by the monoclonal antibody specific to EGFR (cetuximab) [33,34]. Similar effects were observed in mice treated with erlotinib [35] and in mammary epithelial cells treated with tyrphostin AG1478, inhibitors of EGFR tyrosine kinase [36]. These studies were amongst the first to show a relationship between TRPM6, Mg2+ and growth factors and might explain the severe hypomagnesemia observed as a side effect in cancer patients treated with EGFR inhibitors [37]. Most clinical trials reported electrolyte disorders, particularly hypomagnesemia and hypocalcemia, in cancer patients treated with EGFR inhibitors [34]. Whether EGFR inhibitors also influence TRPM7 in cancer patients is unclear.
2. Transient Receptor Potential Melastatin 7 Cation Channel (TRPM7)
2.1. Characteristics and Regulation of TRPM7
Among the more than 300 mammalian ion channels, only TRPM7 and TRPM6 have a C-terminal kinase, which belongs to a specific subfamily of atypical protein kinases (APKs), known as α-kinases, displaying little amino acid sequence similarity to conventional protein kinases [3,38,39]. To date very few α-kinases have been identified. Those that have been described include eukaryotic Elongation Factor 2 Kinase (eEF2K), alpha-kinase 1 (lymphocyte alpha-kinase, LAK or ALPK1), alpha-kinase 2 (heart alpha-kinase, HAK or ALPK2) and alpha-kinase 3 (muscle alpha-kinase, MAK or ALPK3), TRPM6 and TRPM7 [38]. The crystal structure of TRPM7 shows that the N-terminal central catalytic core is structurally similar to the classical protein kinase, whereas the structure of the C-terminal lobe resembles metabolic enzymes with ATP-grasp folds [3,39]. The TRPM7 α-kinase predominantly phosphorylates serine/threonine residues on α-helices. Since the cytoplasmic domain is rich in serine/threonine residues, activation of TRPM7 can induce autophosphorylation as well as phosphorylation of downstream targets (Figure 1) [40,41]. TRPM7-kinase phosphorylates annexin-A1 (Ser5) [42]; myosin IIA (Thr1800, Ser1803 and Ser1808) [43]; eukaryotic elongation factor 2 cognate kinase (eEF2-K) (Ser77) under low Mg2+ concentrations [44]; SMAD2 (Ser465/467) [15]; and phospholipase Cγ2 (PLCγ2) (Ser1164) [45]. TRPM7 is cleaved by caspases at Asp1510, dissociating the kinase from the conducting pore, with the cleaved channel exhibiting enhanced activity, and the kinase domain retaining phosphotransferase activity [46]. TRPM7 cleaved fragments translocate and accumulate in the nucleus, where they phosphorylate specific serines and threonines on histones, affecting gene expression. It seems that different cleaved fragments are expressed in different tissues, implying a distinct role for TRPM7 cleaved fragments in a tissue-specific manner [16]. A similar phenomenon has been described for TRPM6 kinase [47].
2.2. Interactions between TRPM7 Kinase and TRPM7 Channel Domains
Despite intense investigations, there are still controversies regarding the functional significance of the coupling between the α-kinase and the channel domain of TRPM7. Mutations at specific sites that disrupt kinase activity do not affect TRPM7 channel activity and sensitivity to inhibition by divalent cations (Figure 2). In addition, there were no significant differences in channel activity and Ca2+ influx between wild type and TRPM7 kinase mutant cells [48]. These data were confirmed in in vivo experiments, where TRPM7 kinase-dead mutant mice have normal serum Mg2+ levels and normal development [15,19,48]. However, many studies support a link between TRPM7 channel and its C-terminal kinase. In HEK293 cells overexpressing mutant human TRPM7 K1648R and G1799D, activity of the kinase domain and function of the channel domain are altered by changes in sensitivity to intracellular Mg2+ [49]. Interplay between the kinase and channel is supported by studies showing that: (i) cleavage of TRPM7 kinase is associated with increased cation channel activity [46], (ii) Mg2+ nucleotide modulation of TRPM7 channel through the kinase domain [50,51], and (iii) regulation of TRPM7 by cAMP/PKA, which requires a functional kinase domain [52]. Additionally, an in vivo model carrying a truncated kinase domain showed reduced intracellular Mg2+, hypomagnesemia [17] and increased sensitivity to Ca2+ in mast cells [53]. Therefore, it seems that the catalytic activity of the kinase is not essential for channel gating, but somehow it modulates and “fine tunes” channel activity and sensitivity to Mg2+ nucleotides.
2.3. Relationship between TRPM6 and TRPM7
In the plasma membrane, TRPM7 functions as a homodimer, but in some conditions it also heterodimerizes with TRPM6. In some cell-based systems, TRPM6 seems to require TRPM7 for full activation and in the complexed state has been termed TRPM6/TRPM7 [60]. The crucial role of TRPM7 in the complex formation was demonstrated in molecular studies, where TRPM6 deficiency in trophoblast stem cells results in a reduction in TRPM6/7 currents, while deficiency in TRPM7 completely abolished the TRPM6/7 currents [53]. TRPM6 and TRPM7 possess several phosphorylation sites and can be phosphorylated independently of autophosphorylation, TRPM7 can also be transphosphorylated by TRPM6 (Figure 3) [61]. This cross activation was demonstrated in co-transfection experiments, which showed that TRPM6 expression was able to transphosphorylate serine and threonine motifs in TRPM7-K1646R kinase dead mutant. Transphosphorylation of TRPM6 by TRPM7 is very weak. These phosphorylated residues are present in the N-terminal, channel, and C terminus, including the kinase domain of the TRPM7-K1646R [61]. These phenomena have been demonstrated in cell models but the (patho) physiological significance in vivo remains to be determined. TRPM7 seems to interact with other Mg2+ channels. In DT40 B cells and colon carcinoma cells, TRPM7 deficiency was associated with increased expression of MagT1 [62,63,64].
2.4. TRPM7 and Cell Function
The indispensable role of TRPM7 in cellular biology is likely linked to its important role in the regulation of homeostasis of divalent cations Mg2+, Zn2+, and Ca2+. Mg2+ and Zn2+ are catalytic and structural cofactors of numerous enzymes and are major regulators of signaling molecules, DNA stability, cell cycle and transcription factors [65,66]. Deficiency of Mg2+ and Zn2+ suppress cell cycle progression leading to growth failure [67,68]. Mg2+ supplementation and overexpression of Mg2+ transporters rescued growth impairment caused by TRPM7 deficiency [17,62]. Ca2+, which is also influenced by TRPM7, is critically involved in controlling cell function including cell proliferation, contraction, secretion, migration and differentiation [69,70,71]. Associated with many of the Ca2+- and Mg2+- regulated signaling pathways is the activation of tyrosine kinases [35,72,73,74,75]. In vascular cells, TRPM7 appears to be the most important cation channel involved in controlling [Mg2+]i. TRPM7 is regulated by vasoactive factors, such as AngII, aldosterone, endothelin-1 and bradykinin [76,77,78,79,80]. Ang II regulates vascular TRPM7 acutely by inducing phosphorylation and chronically by increasing expression at the mRNA and protein levels [76,79]. Also, siRNA downregulation of vascular TRPM7 caused a reduction in [Mg2+]i and attenuated Ang II-mediated VSMC growth [7]. VSMCs from hypertensive rats exhibit reduced TRPM7 expression and decreased TRPM7 activation, as assessed by translocation of annexin-1 (TRPM7 kinase target) to the membrane [79]. Findings from cell-based studies and animal models indicate that TRPM7 activation and increased Mg2+ influx are vasoprotective as they protect against vascular calcification, oxidative stress and fibrosis [81].
3. Receptor Tyrosine Kinase Signaling and Mg2+
3.1. Characteristics and Regulation of Receptor Tyrosine Kinases
Protein kinases induce phosphorylation by catalyzing the transfer of phosphate from adenosine triphosphate (ATP) to serine, threonine and tyrosine residues on protein substrates and are key enzymes in the regulation of intracellular signaling pathways [82,83]. These ATP- and phosphate-dependent processes have an obligatory requirement for Mg2+. Data from the human genome revealed that of the proteins phosphorylated on tyrosine residues, ≈58 are receptor tyrosine kinases and ≈32 are non-receptor tyrosine kinases [32,84]. All RTKs have a similar molecular architecture, comprising three major domains, including an extracellular ligand-binding domain, an intracellular tyrosine kinase domain and a transmembrane domain [31]. RTKs are typically activated by growth factors [85,86,87,88].
Magnesium is a crucial divalent cation required for the activity of protein kinases, including RTKs (VEGFR, EGFR, FGFR, PDGFR) and non-receptor tyrosine kinases (Src, Abl, Jak, FAK, SOCS) [89]. In cell-based studies, high Mg2+ concentration causes increased tyrosine kinase activity. Data obtained with crystallography experiments revealed that two Mg2+ molecules are required for enzyme activity and phosphoryl transfer: one bound to ATP (Mg-ATP) situated between the small and large lobes of the kinase domain, bound to β and γ-phosphates and to the aspartate of the DFG (Asp-Phe-Gly), which are the first residues to be activated in protein kinases; and in high [Mg2+]i conditions, another Mg2+ binds to α and γ-phosphates and to the asparagine amide nitrogen within the catalytic loop [90]. The importance of Mg2+ in the regulation of kinase activity occurs in four steps: (i) Mg-ATP binds to the enzyme; (ii) the kinase binds to the protein substrate and catalyzes the transfer of the phosphoryl group; (iii) phosphorylated protein and Mg2+ are released; and (iv) Mg2+-ADP is released and the catalytic cycle is completed [91] (Figure 4).
3.2. Growth Factors and Receptor Tyrosine Kinase Signaling
Following binding of growth factors to their specific RTKs, cytoplasmic proteins containing Src homology region 2 (SH2) or phosphotyrosine-binding (PTB) domains are recruited to the cell membrane. These recruited proteins either have intrinsic enzymatic activity, such as Src and PLCγ, or serve as docking proteins that function as “assembly platforms” and recruit additional enzymes [32,75,85,92]. Activated RTKs are able to trigger a wide range of downstream signaling pathways, including RAS/RAF/MEK/MAPK, PLCγ/PKC, PI3K/AKT/mTOR, and JAK/STAT (Figure 5) [93].
Activated RTKs bind to the adaptive protein growth factor receptor-bound 2(Grb2) and recruit Son of Sevenless homolog protein (SOS), which interacts with its downstream target Ras, a small GTP binding protein, and transforms it to the active conformation by exchanging GDP for GTP [94]. Ras recruits the serine/threonine protein kinase, Raf, to the membrane, where it becomes activated by phosphorylation. Once activated, Raf, a MAP3K, phosphorylates MAP2Ks, MEK1 and MEK2, at specific serine residues, and activated MEK1/2 in turn catalyzes phosphorylation of ERK1/2 at threonine and tyrosine residues [95]. Activated ERK1/2 phosphorylates many downstream target proteins and transcription factors involved in cell function [32,96].
The phosphoinositol-3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) signaling pathway is also initiated by binding of growth factors to RTKs. Activated RTKs recruit the PI3K to the plasma membrane where the PI3K subunit p110 catalyzes phosphorylation of phosphatidylinositol 4,5-bisphosphate (PIP2) to phosphatidylinositol 3,4,5-triphosphate (PIP3). PIP3 then provides docking sites for signaling proteins, including AKT and 3-phosphoinositide-dependent kinase 1 (PDK1) [97,98]. Activated AKT phosphorylates many other downstream proteins, such as glycogen synthase kinase 3 (GSK3), the forkhead family of transcription factors (FOXOs) and mTOR. Multiple components of the PI3K/AKT/mTOR pathway activated by RKTs play a pivotal role in the regulation of cell growth, proliferation and differentiation [93,98,99,100].
Another signaling system linked to RTKs is PLCγ/protein kinase C (PKC). Upon growth factor stimulation, the phosphorylated tyrosine residues of RTKs interact with SH2 domains of PLCγ and lead to its activation. PLCγ then hydrolyses PIP2 into two second messengers, inositol 1,4,5-trisphosphate (IP3) and diacylglycerol (DAG). IP3 binds to its receptor on the endoplasmic reticulum (ER) surface, whereas DAG mediates activation of PKC [101,102]. This process has a key role in regulating intracellular Ca2+ through multiple Ca2+ channels [103,104,105]. PLCγ/PKC is also involved in the regulation of cell proliferation, contraction and migration [106,107,108].
The Janus kinase/signal transducer and activator of transcription (JAK/STAT) and the Src family kinases (SFKs) are additional signaling pathways associated with RTK activation. Src family members, including Src, Fyn and Yes, are recruited on RTKs, such as EGFR, FGFR and IGFR, and transmit mitogenic signals to regulate DNA synthesis, cell survival, cell adhesion, motility and growth [32]. Once activated, STATs enter the nucleus and bind to specific regulatory sequences in target genes regulating transcription [109].
4. Cross-Talk between TRPM6/7, Receptor Tyrosine Kinases and Signaling Kinases
4.1. Regulation of TRPM6/7 by Receptor Tyrosine Kinases
Upon ligand binding, VEGFR and EGFR undergo receptor dimerization and tyrosine phosphorylation leading to recruitment of effector proteins and activation of downstream cascades. These processes are highly regulated and are dependent on intracellular Mg2+. Increased RTK activation and abnormal growth factor-mediated tyrosine kinase signaling is associated with uncontrolled cell proliferation in cancer [110]. Accordingly, inhibitors of VEGFR and EGFR tyrosine kinases have been used to treat several cancers. However unexpectedly these drugs have been associated with unwanted secondary effects, including hypertension, cardiovascular and renal toxicity, and electrolyte disorders [110,111,112,113].
One of the most significant electrolyte abnormalities associated with EGFR inhibitors in the treatment of cancer is renal magnesium wasting and hypomagnesemia. In extreme cases, patients need intravenous magnesium treatment and withdrawal of the EGFR inhibitor. This has major impact on the effectiveness of cancer treatment [110,111,112,113,114]. Molecular mechanisms underlying these side effects have been attributed to abnormal function of TRPM6, phenomena that were first described in patients carrying a mutation in the EGF gene and who have severe hypomagnesemia and cognitive disability. EGF binding to EGFR promotes TRPM6 translocation from cytosol to membrane in kidney cells [33]. This process was blocked by the EGFR monoclonal antibody, cetuximab [33,34]. Hence, EGF-EGFR signaling involves TRPM6, which promotes Mg2+ transport into cells. Inhibition of these processes with EGFR inhibitors decreases TRPM6 activity and reduces Mg2+ influx into cells, promoting Mg2+ excretion in the kidney with resultant Mg2+ wasting and hypomagnesemia.
Emerging evidence suggests that TRPM7 may also be regulated by growth factor signaling through EGFR, as well as VEGFR [115,116,117]. Studies in transfected chinese hamster ovary (CHO) cells showed inhibition of TRPM7 channel activity by EGF through mechanisms involving PLCγ and PIP2 [117]. However, Gao et al. found that in pulmonary cancer cell lines, EGF, through its receptor, enhanced the cell membrane protein expression and currents of TRPM7, a process associated with cell migration [118]. The difference in models and cell lines may explain the conflicting data obtained in these two studies. In hippocampal neurons, nerve growth factor (NGF) signals through receptor tyrosine kinase TrkA, reducing the outward rectifying TRPM7-like current, effects blocked by inhibitors of TrkA and PLC [119]. TrkA activation by NGF prevented upregulation of TRPM7 expression through PI3K in hippocampal neurons subjected to ischemia-reperfusion and oxygen-glucose deprivation [120]. Moreover, PDGF stimulation was shown to increase TRPM7 expression in HSC-T6 hepatic stellate cells. The non-specific TRPM7 inhibitor 2-aminoethyl diphenyl borinate (2-APB) diminished PDGF-mediated activation of p-AKT and p-ERK, further suggesting a regulatory role for TRPM7 upstream of AKT and ERK [121].
RTKs induce phosphorylation of downstream kinases that influence TRPM7 activity (Figure 6). However these processes seem to be related to changes in the intracellular concentration of Mg2+, because when [Mg2+]i is low, PLC activation is associated with TRPM7 inhibition, whereas in normal [Mg2+]i conditions, activation of PLC increases TRPM7 currents [122]. In prostate cells, TRPM7 expression was mediated by Ca2+-dependent activation of ERK, since ERK inhibition reduced TRPM7 expression [123]. Ang II, which transactivates TKRs, increased TRPM7 expression through the AT1 receptor and ERK [124]. Bradykinin, another vasoactive peptide, regulates TPRM7 and its downstream target annexin-1 through PLC and c-Src dependent pathways, which have an important role in VSMC Mg2+ homeostasis and cell migration and invasion [78]. The inflammatory mediator interleukin-18 (IL-18) activated TRPM7 currents and upregulated TRPM7 expression in an ERK1/2-dependent manner, processes that influence osteogenic differentiation of VSMCs [125]. In HEK293 cells, interleukin-6 (IL-6) inhibited TRPM7 currents through JAK2-STAT3 signaling and appeared to be independent of the TPRM7 α-kinase domain. Regulation of TRPM7 by IL-6 signaling may result from JAK2-STAT3 mediated phosphorylation of TRPM7 [126].
4.2. Regulation of Signaling Kinases by TRPM7
While kinases regulate TRPM7, these kinases themselves may be targets of TRPM7. MAP kinases are regulated by TRPM7. In human endothelial cells, downregulation of TRPM7 using a siRNA approach was associated with increased ERK1/2 phosphorylation with no effects on p38MAPK or JNK activation [127]. On the other hand, in HEK293 cells overexpressing TRPM7, ERK1/2 phosphorylation was reduced, while phosphorylation of p38MAPK and JNK was increased [128]. In a rat hepatic stellate cell line (HSC-T6), upregulation of TRPM7 promoted PDGF-induced activation of ERK1/2 and AKT pathways [121]. In mouse cortical astrocytes and in rat VSMCs, TRPM7 siRNA was associated with reduced ERK1/2 phosphorylation [129,130]. Additionally, TRPM7 siRNA reduced activation of Src, p38 MAPK, ERK1/2 and JNK in a breast cancer cell line [131,132].
A regulatory role of TRPM7 in MAPK signaling has also been observed using pharmacological approaches in cancer tissue and cell lines. In glioblastoma cells, naltriben, a pharmacological activator of TRPM7 channel, enhanced ERK1/2 phosphorylation and proliferation, with no effect on the PI3K/AKT pathway (150). Carvacrol, a non-specific inhibitor of TRPM7, suppressed phosphorylation of ERK1/2 and AKT, reducing cell proliferation [133,134,135]. Thus, there is accumulating evidence supporting a role for TRPM7 in the regulation of MAPK signaling.
The PI3K/AKT pathway is also affected by TRPM7. In mouse chondrocytes, overexpression of TRPM7 stimulated the AKT pathway, while TRPM7 silencing inhibited AKT signaling [136]. In human osteoblasts, Mg2+-induced increase in mRNA expression of chemotaxis-related genes was attenuated by TRPM7 siRNA and PI3K inhibition, suggesting a link between TRPM7 and PI3K activation mediated by Mg2+.
STAT3 is also modulated by TRPM7. TRPM7 siRNA reduced activation of STAT3 (Tyr705) by affecting phosphorylation of the upstream protein JAK2, a non-receptor tyrosine kinase [137]. Reduced TRPM7 expression was associated with inhibition of Notch activation (Notch1, JAG1, Hey2, and Survivin), involved in angiogenesis and tumor growth mediated by VEGFR and EGFR [138,139]. In breast cancer cells, silencing TRPM7 reduced EGF-induced STAT3 activation and decreased phosphorylation levels of Src, with reduced TRPM7-mediated migration and invasion [131,132].
5. TRPM7and Kinase Signaling—Pathophysiological Implications
TRPM7-regulated Mg2+ has versatile biological functions, contributing to all vital cellular processes, including stability of tertiary structures of DNA and RNA, energy metabolism, enzyme activity, signaling, cell cycle progression and cell differentiation [140,141]. The significance of TRPM7 in development and cell viability was demonstrated in mice with global deletion of TRPM7. TRPM7 knockout mice are embryonic lethal and cardiac-targeted knockout of TRPM7 causes impaired embryonic development of the heart [14,142]. The regulatory role of TRPM7 in cell differentiation has been highlighted in various cell types. In mesenchymal stromal cells, TRPM7 mediates shear stress and silencing TRPM7 accelerates osteogenic differentiation [143]. TRPM7 was also shown to mediate differentiation in hepatic cells, lung fibroblasts, dental pulp stem cells and T cells [14,144,145,146]. Some of these studies investigated underlying mechanisms of the involvement of TRPM7 in cell differentiation. In addition to the contribution of Ca2+ and Mg2+, an interaction between TRPM7 and RTK downstream effectors, including PI3K, AKT and ERK1/2, have been implicated [81,125,145,147,148,149].
Developmental experiments using zebrafish models of TRPM7 mutants showed abnormal pancreas development, which was associated with increased expression of Suppressor of cytokine signaling (SOCS)-3a, the zebrafish homologue to the human SOCS3, which is part of a classical negative feedback system that influences cytokine signal transduction by reducing activation of JAK-STAT3. Mg2+ supplementation partially attenuated the phenotype [150]. Using genetic and pharmacologic inhibitors, 2-APB or TRPM7-siRNA, it was further shown that TRPM7 modulates effects of PDGF-BB on cell proliferation by regulating activity of cell cycle proteins, including cyclin D1, PCNA and CDK4, through mechanisms that involve tyrosine kinases, ERK1/2 and PI3K [121].
The effects of TRPM7 in pathologic cell differentiation has been intensively investigated in cancer cells, including pancreas, ovary, breast and adenocarcinoma of lungs and prostate [118]. Expression of TRPM7 in these cancers was associated with increased expression of proliferative markers. Epithelial-mesenchymal transition is involved in fibrosis and cancer metastasis and involves EGF activation, effects attenuated by TRPM7 siRNA by mechanisms involving ERK1/2 and STAT3 [132].
Cancer stem cells are cells found within tumors and hematological cancers that have features of normal stem cells, particularly the ability to self-renew and differentiate into multiple cell types [151]. They are rare immortal cells within tumors and are able to induce growth of new tumors and metastases. They are found in numerous tumors and have been considered attractive targets for anti-cancer therapy [152]. Many pathways have been implicated in the regulation of stem cell self-renewal and oncogenesis, including signaling through Notch, Wnt and sonic hedgehog (Shh) [151,152]. In epithelial tumors, epithelial-mesenchymal transition (EMT) seems to be important in cancer stem cell function. Recent evidence indicates that cancer stem cell regulation involves TRPM7-dependent pathways. In lung cancer cells, TRPM7 expression was increased and associated with enhanced SOX2, KLF4, and CD133, Hsp90α, uPA, and MMP2 [153]. TRPM7-silencing inhibited epithelial-to-mesenchymal transition (EMT), suppressed stemness markers and phenotypes, and attenuated activation of the Hsp90α/uPA/MMP2 axis. These effects were ameliorated by waixenicin A, a bioactive extract of soft coral, which downregulated TRPM7 and oncogenic markers. Hence, waixenicin A was suggested as an anticancer therapy, by inhibiting lung cancer stem cells through TRPM7 inhibition [153]. In glioma tumorigenesis cancer stem cells, TRPM7 activates JAK2/STAT3- aldehyde dehydrogenase1 (ALDH1) and Notch signaling pathways, contributing to the regulation of cell proliferation, migration and invasion [137]. TRPM7 has also been implicated in epithelial-mesenchymal transition ovarian cancer stem cells, through calcium-related PI3K/AKT oncogenic signaling [154]. In mesenchymal stromal cells (MSCs), TRPM7 senses mechanical stimuli, such as intermittent fluid shear stress and membrane tension, and regulates Ca2+ influx and phosphorylation of signaling kinases, Smad1/5 and p38 MAPK, whereas TRPM7 knockdown decreases MSC proliferation and viability, and induces cell death [143]. Additionally, Mg2+ supplementation promoted osteogenic differentiation through the activation of Notch signaling in MSCs, which was decreased by 2-APB [143,155,156,157]. In human dental pulp stem cells, silencing TRPM7 inhibited proliferation, migration, and osteogenic differentiation, supporting a role for TRPM7 in the dental pulp repair process [146]. Together, these studies suggest that dysregulation of TRPM7 is important in cancer stem cell regulation and that TRPM7 may be a therapeutic target for oncogenesis [158].
Interactions between RTKs, such as VEGFR and EGFR, TRPM6/7 and Mg2+, have also been implicated in preeclampsia. In a study examining placentas from women with preeclampsia compared to placentas from normotensive women, pre-eclamptic placentas had reduced expression of TRPM6 and TRPM7 [159], which was linked to altered cellular Mg2+ homeostasis. These phenomena were associated with decreased placental VEGF expression in preeclampsia versus normotensive pregnancies, effects normalized by treatment with MgSO4 [160]. Together these findings suggest an important relationship between VEGF, TRPM6/7 and Mg2+ in preeclampsia. This is clinically important because it provides a mechanistic basis for the therapeutic use of MgSO4 as the drug of choice for preeclampsia [161].
6. Conclusions and Perspectives
TRPM7 and TRPM6 are unique proteins characterized by their channel and kinase domains, which differentially regulate cell signaling and function. TRPM6 is mostly expressed in epithelial cells of the kidney and gastrointestinal system, whereas TRPM7 is ubiquitously expressed. In the kidney, TRPM6 and cellular Mg2+ homeostasis are regulated by EGF signaling through EGFR. Inhibition of EGFR signaling impairs TRPM6 function leading to renal Mg2+ wasting and consequent hypomagnesemia in cancer patients treated with EGFR inhibitors [161]. It is still unclear if similar processes influence TRPM7. Experimental studies, mainly in cell models, indicate that VEGFR and EGFR influence TRPM7 activity and Ca2+ and Mg2+ mobilization. Changes in TRPM7 activity and altered Mg2+ homeostasis have a significant effect on tyrosine kinase signaling. Hence, RTKs influence TRPM7, which in turn may influence tyrosine kinase signaling. Most studies have focused on EGFR signaling, but growing evidence indicates that VEGFR and PDGFR also signal through TRPM7. The putative cross-talk between TRPM7 and RTKs has clinical relevance, because inhibitors of RTKs, including VEGFR and EGFR, are increasingly being used to treat cancer and inflammatory diseases. Hence, knowing mechanisms whereby these drugs cause hypomagnesemia is important so that unwanted side effects can be managed in a mechanism-specific manner. The field of TRPM6/7 and RTKs is still immature and further studies are required to unravel the complex interplay between these systems.
Funding
R.M.T. is supported by a British Heart Foundation Award (CH/12/429762). A.C.M. is supported through a Walton Fellowship, University of Glasgow. Z.Z. is funded by China Scholarship Council (201708060309).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Levitan, I.B.; Cibulsky, S.M. Biochemistry. TRP ion channels--two proteins in one. Science 2001, 293, 1270–1271. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Li, Z.; Li, J.; Hulse, R.E.; Santa-Cruz, A.; Valinsky, W.C.; Abiria, S.A.; Krapivinsky, G.; Zhang, J.; Clapham, D.E. Structure of the mammalian TRPM7, a magnesium channel required during embryonic development. Proc. Natl. Acad. Sci. USA 2018, 115, E8201–E8210. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Hong, C.; Kim, B.J.; So, I. The Pathophysiologic Roles of TRPM7 Channel. Korean J. Physiol. Pharmacol. 2014, 18, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadolni, W.; Zierler, S. The Channel-Kinase TRPM7 as Novel Regulator of Immune System Homeostasis. Cells 2018, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Nadler, M.J.; Hermosura, M.C.; Inabe, K.; Perraud, A.L.; Zhu, Q.; Stokes, A.J.; Kurosaki, T.; Kinet, J.P.; Penner, R.; Scharenberg, A.M.; et al. LTRPC7 is a Mg.ATP-regulated divalent cation channel required for cell viability. Nature 2001, 411, 590–595. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yao, G.; Savoia, C.; Touyz, R.M. Transient receptor potential melastatin 7 ion channels regulate magnesium homeostasis in vascular smooth muscle cells: Role of angiotensin II. Circ. Res. 2005, 96, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Gwanyanya, A.; Amuzescu, B.; Zakharov, S.I.; Macianskiene, R.; Sipido, K.R.; Bolotina, V.M.; Vereecke, J.; Mubagwa, K. Magnesium-inhibited, TRPM6/7-like channel in cardiac myocytes: Permeation of divalent cations and pH-mediated regulation. J. Physiol. 2004, 559, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Chen, S.; Xiao, C.; Jia, Y.; Guo, J.; Jiang, J.; Liu, P. TRPM7 is involved in angiotensin II induced cardiac fibrosis development by mediating calcium and magnesium influx. Cell Calcium 2014, 55, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Castiglioni, S.; Cazzaniga, A.; Trapani, V.; Cappadone, C.; Farruggia, G.; Merolle, L.; Wolf, F.I.; Iotti, S.; Maier, J.A. Magnesium homeostasis in colon carcinoma LoVo cells sensitive or resistant to doxorubicin. Sci. Rep. 2015, 5, 16538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stritt, S.; Nurden, P.; Favier, R.; Favier, M.; Ferioli, S.; Gotru, S.K.; van Eeuwijk, J.M.; Schulze, H.; Nurden, A.T.; Lambert, M.P.; et al. Defects in TRPM7 channel function deregulate thrombopoiesis through altered cellular Mg(2+) homeostasis and cytoskeletal architecture. Nat. Commun. 2016, 7, 11097. [Google Scholar] [CrossRef]
- Tashiro, M.; Inoue, H.; Konishi, M. Physiological pathway of magnesium influx in rat ventricular myocytes. Biophys. J. 2014, 107, 2049–2058. [Google Scholar] [CrossRef]
- Abed, E.; Moreau, R. Importance of melastatin-like transient receptor potential 7 and cations (magnesium, calcium) in human osteoblast-like cell proliferation. Cell Prolif. 2007, 40, 849–865. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Desai, B.N.; Navarro, B.; Donovan, A.; Andrews, N.C.; Clapham, D.E. Deletion of Trpm7 disrupts embryonic development and thymopoiesis without altering Mg2+ homeostasis. Science 2008, 322, 756–760. [Google Scholar] [CrossRef]
- Romagnani, A.; Vettore, V.; Rezzonico-Jost, T.; Hampe, S.; Rottoli, E.; Nadolni, W.; Perotti, M.; Meier, M.A.; Hermanns, C.; Geiger, S.; et al. TRPM7 kinase activity is essential for T cell colonization and alloreactivity in the gut. Nat. Commun. 2017, 8, 1917. [Google Scholar] [CrossRef] [Green Version]
- Krapivinsky, G.; Krapivinsky, L.; Manasian, Y.; Clapham, D.E. The TRPM7 chanzyme is cleaved to release a chromatin-modifying kinase. Cell 2014, 157, 1061–1072. [Google Scholar] [CrossRef]
- Ryazanova, L.V.; Rondon, L.J.; Zierler, S.; Hu, Z.; Galli, J.; Yamaguchi, T.P.; Mazur, A.; Fleig, A.; Ryazanov, A.G. TRPM7 is essential for Mg(2+) homeostasis in mammals. Nat. Commun. 2010, 1, 109. [Google Scholar] [CrossRef]
- Antunes, T.T.; Callera, G.E.; He, Y.; Yogi, A.; Ryazanov, A.G.; Ryazanova, L.V.; Zhai, A.; Stewart, D.J.; Shrier, A.; Touyz, R.M. Transient Receptor Potential Melastatin 7 Cation Channel Kinase: New Player in Angiotensin II-Induced Hypertension. Hypertension 2016, 67, 763–773. [Google Scholar] [CrossRef]
- Kaitsuka, T.; Katagiri, C.; Beesetty, P.; Nakamura, K.; Hourani, S.; Tomizawa, K.; Kozak, J.A.; Matsushita, M. Inactivation of TRPM7 kinase activity does not impair its channel function in mice. Sci. Rep. 2014, 4, 5718. [Google Scholar] [CrossRef]
- Krebs, J.; Agellon, L.B.; Michalak, M. Ca(2+) homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef]
- Massullo, P.; Sumoza-Toledo, A.; Bhagat, H.; Partida-Sanchez, S. TRPM channels, calcium and redox sensors during innate immune responses. Semin. Cell Dev. Biol. 2006, 17, 654–666. [Google Scholar] [CrossRef]
- Clark, K.; Langeslag, M.; van Leeuwen, B.; Ran, L.; Ryazanov, A.G.; Figdor, C.G.; Moolenaar, W.H.; Jalink, K.; van Leeuwen, F.N. TRPM7, a novel regulator of actomyosin contractility and cell adhesion. EMBO J. 2006, 25, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Schappe, M.S.; Szteyn, K.; Stremska, M.E.; Mendu, S.K.; Downs, T.K.; Seegren, P.V.; Mahoney, M.A.; Dixit, S.; Krupa, J.K.; Stipes, E.J.; et al. Chanzyme TRPM7 Mediates the Ca(2+) Influx Essential for Lipopolysaccharide-Induced Toll-Like Receptor 4 Endocytosis and Macrophage Activation. Immunity 2018, 48, 59–74. [Google Scholar] [CrossRef]
- Faouzi, M.; Kilch, T.; Horgen, F.D.; Fleig, A.; Penner, R. The TRPM7 channel kinase regulates store-operated calcium entry. J. Physiol. 2017, 595, 3165–3180. [Google Scholar] [CrossRef]
- Beesetty, P.; Wieczerzak, K.B.; Gibson, J.N.; Kaitsuka, T.; Luu, C.T.; Matsushita, M.; Kozak, J.A. Inactivation of TRPM7 kinase in mice results in enlarged spleens, reduced T-cell proliferation and diminished store-operated calcium entry. Sci. Rep. 2018, 8, 3023. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Branigan, D.; Xiong, Z.G. Zinc-induced neurotoxicity mediated by transient receptor potential melastatin 7 channels. J. Biol. Chem. 2010, 285, 7430–7439. [Google Scholar] [CrossRef]
- Abiria, S.A.; Krapivinsky, G.; Sah, R.; Santa-Cruz, A.G.; Chaudhuri, D.; Zhang, J.; Adstamongkonkul, P.; DeCaen, P.G.; Clapham, D.E. TRPM7 senses oxidative stress to release Zn(2+) from unique intracellular vesicles. Proc. Natl. Acad. Sci. USA 2017, 114, E6079–E6088. [Google Scholar] [CrossRef]
- Yu, P.; Wang, Q.; Zhang, L.H.; Lee, H.C.; Zhang, L.; Yue, J. A cell permeable NPE caged ADP-ribose for studying TRPM2. PLoS ONE 2012, 7, e51028. [Google Scholar] [CrossRef]
- Wagner, T.F.; Drews, A.; Loch, S.; Mohr, F.; Philipp, S.E.; Lambert, S.; Oberwinkler, J. TRPM3 channels provide a regulated influx pathway for zinc in pancreatic beta cells. Pflugers Arch. 2010, 460, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Tominaga, M. Extracellular zinc ion regulates transient receptor potential melastatin 5 (TRPM5) channel activation through its interaction with a pore loop domain. J. Biol. Chem. 2013, 288, 25950–25955. [Google Scholar] [CrossRef] [PubMed]
- Butti, R.; Das, S.; Gunasekaran, V.P.; Yadav, A.S.; Kumar, D.; Kundu, G.C. Receptor tyrosine kinases (RTKs) in breast cancer: Signaling, therapeutic implications and challenges. Mol. Cancer 2018, 17, 34. [Google Scholar] [CrossRef]
- Segaliny, A.I.; Tellez-Gabriel, M.; Heymann, M.F.; Heymann, D. Receptor tyrosine kinases: Characterisation, mechanism of action and therapeutic interests for bone cancers. J. Bone Oncol. 2015, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Thebault, S.; Alexander, R.T.; Tiel Groenestege, W.M.; Hoenderop, J.G.; Bindels, R.J. EGF increases TRPM6 activity and surface expression. J. Am. Soc. Nephrol. 2009, 20, 78–85. [Google Scholar] [CrossRef]
- Groenestege, W.M.; Thebault, S.; van der Wijst, J.; van den Berg, D.; Janssen, R.; Tejpar, S.; van den Heuvel, L.P.; van Cutsem, E.; Hoenderop, J.G.; Knoers, N.V.; et al. Impaired basolateral sorting of pro-EGF causes isolated recessive renal hypomagnesemia. J. Clin. Investig. 2007, 117, 2260–2267. [Google Scholar] [CrossRef] [Green Version]
- Dimke, H.; van der Wijst, J.; Alexander, T.R.; Meijer, I.M.; Mulder, G.M.; van Goor, H.; Tejpar, S.; Hoenderop, J.G.; Bindels, R.J. Effects of the EGFR Inhibitor Erlotinib on Magnesium Handling. J. Am. Soc. Nephrol. 2010, 21, 1309–1316. [Google Scholar] [CrossRef] [Green Version]
- Trapani, V.; Arduini, D.; Luongo, F.; Wolf, F.I. EGF stimulates Mg(2+) influx in mammary epithelial cells. Biochem. Biophys. Res. Commun. 2014, 454, 572–575. [Google Scholar] [CrossRef]
- Hofheinz, R.D.; Segaert, S.; Safont, M.J.; Demonty, G.; Prenen, H. Management of adverse events during treatment of gastrointestinal cancers with epidermal growth factor inhibitors. Crit. Rev. Oncol. Hematol. 2017, 114, 102–113. [Google Scholar] [CrossRef]
- Middelbeek, J.; Clark, K.; Venselaar, H.; Huynen, M.A.; van Leeuwen, F.N. The alpha-kinase family: An exceptional branch on the protein kinase tree. Cell Mol. Life Sci. 2010, 67, 875–890. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Matsushita, M.; Nairn, A.C.; Kuriyan, J. Crystal structure of the atypical protein kinase domain of a TRP channel with phosphotransferase activity. Mol. Cell 2001, 7, 1047–1057. [Google Scholar] [CrossRef]
- Kim, T.Y.; Shin, S.K.; Song, M.Y.; Lee, J.E.; Park, K.S. Identification of the phosphorylation sites on intact TRPM7 channels from mammalian cells. Biochem. Biophys. Res. Commun. 2012, 417, 1030–1034. [Google Scholar] [CrossRef]
- Clark, K.; Middelbeek, J.; Morrice, N.A.; Figdor, C.G.; Lasonder, E.; van Leeuwen, F.N. Massive autophosphorylation of the Ser/Thr-rich domain controls protein kinase activity of TRPM6 and TRPM7. PLoS ONE 2008, 3, e1876. [Google Scholar] [CrossRef] [PubMed]
- Ryazanova, L.V.; Dorovkov, M.V.; Ansari, A.; Ryazanov, A.G. Characterization of the protein kinase activity of TRPM7/ChaK1, a protein kinase fused to the transient receptor potential ion channel. J. Biol. Chem. 2004, 279, 3708–3716. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Middelbeek, J.; Lasonder, E.; Dulyaninova, N.G.; Morrice, N.A.; Ryazanov, A.G.; Bresnick, A.R.; Figdor, C.G.; van Leeuwen, F.N. TRPM7 regulates myosin IIA filament stability and protein localization by heavy chain phosphorylation. J. Mol. Biol. 2008, 378, 790–803. [Google Scholar] [CrossRef]
- Perraud, A.L.; Zhao, X.; Ryazanov, A.G.; Schmitz, C. The channel-kinase TRPM7 regulates phosphorylation of the translational factor eEF2 via eEF2-k. Cell Signal 2011, 23, 586–593. [Google Scholar] [CrossRef]
- Deason-Towne, F.; Perraud, A.L.; Schmitz, C. Identification of Ser/Thr phosphorylation sites in the C2-domain of phospholipase C gamma2 (PLCgamma2) using TRPM7-kinase. Cell Signal 2012, 24, 2070–2075. [Google Scholar] [CrossRef]
- Desai, B.N.; Krapivinsky, G.; Navarro, B.; Krapivinsky, L.; Carter, B.C.; Febvay, S.; Delling, M.; Penumaka, A.; Ramsey, I.S.; Manasian, Y.; et al. Cleavage of TRPM7 releases the kinase domain from the ion channel and regulates its participation in Fas-induced apoptosis. Dev. Cell 2012, 22, 1149–1162. [Google Scholar] [CrossRef] [Green Version]
- Krapivinsky, G.; Krapivinsky, L.; Renthal, N.E.; Santa-Cruz, A.; Manasian, Y.; Clapham, D.E. Histone phosphorylation by TRPM6’s cleaved kinase attenuates adjacent arginine methylation to regulate gene expression. Proc. Natl. Acad. Sci. USA 2017, 114, E7092–E7100. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, M.; Kozak, J.A.; Shimizu, Y.; McLachlin, D.T.; Yamaguchi, H.; Wei, F.Y.; Tomizawa, K.; Matsui, H.; Chait, B.T.; Cahalan, M.D.; et al. Channel function is dissociated from the intrinsic kinase activity and autophosphorylation of TRPM7/ChaK1. J. Biol. Chem. 2005, 280, 20793–20803. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, C.; Perraud, A.L.; Johnson, C.O.; Inabe, K.; Smith, M.K.; Penner, R.; Kurosaki, T.; Fleig, A.; Scharenberg, A.M. Regulation of vertebrate cellular Mg2+ homeostasis by TRPM7. Cell 2003, 114, 191–200. [Google Scholar] [CrossRef]
- Demeuse, P.; Penner, R.; Fleig, A. TRPM7 channel is regulated by magnesium nucleotides via its kinase domain. J. Gen. Physiol. 2006, 127, 421–434. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Z.; Lis, A.; Penner, R.; Fleig, A. TRPM7 is regulated by halides through its kinase domain. Cell Mol. Life Sci. 2013, 70, 2757–2771. [Google Scholar] [CrossRef] [Green Version]
- Takezawa, R.; Schmitz, C.; Demeuse, P.; Scharenberg, A.M.; Penner, R.; Fleig, A. Receptor-mediated regulation of the TRPM7 channel through its endogenous protein kinase domain. Proc. Natl. Acad. Sci. USA 2004, 101, 6009–6014. [Google Scholar] [CrossRef] [Green Version]
- Chubanov, V.; Ferioli, S.; Wisnowsky, A.; Simmons, D.G.; Leitzinger, C.; Einer, C.; Jonas, W.; Shymkiv, Y.; Bartsch, H.; Braun, A.; et al. Epithelial magnesium transport by TRPM6 is essential for prenatal development and adult survival. Elife 2016, 5, e20914. [Google Scholar] [CrossRef]
- Li, M.; Du, J.; Jiang, J.; Ratzan, W.; Su, L.T.; Runnels, L.W.; Yue, L. Molecular determinants of Mg2+ and Ca2+ permeability and pH sensitivity in TRPM6 and TRPM7. J. Biol. Chem. 2007, 282, 25817–25830. [Google Scholar] [CrossRef]
- Krapivinsky, G.; Mochida, S.; Krapivinsky, L.; Cibulsky, S.M.; Clapham, D.E. The TRPM7 ion channel functions in cholinergic synaptic vesicles and affects transmitter release. Neuron 2006, 52, 485–496. [Google Scholar] [CrossRef]
- Hermosura, M.C.; Nayakanti, H.; Dorovkov, M.V.; Calderon, F.R.; Ryazanov, A.G.; Haymer, D.S.; Garruto, R.M. A TRPM7 variant shows altered sensitivity to magnesium that may contribute to the pathogenesis of two Guamanian neurodegenerative disorders. Proc. Natl. Acad. Sci. USA 2005, 102, 11510–11515. [Google Scholar] [CrossRef]
- Crawley, S.W.; Cote, G.P. Identification of dimer interactions required for the catalytic activity of the TRPM7 alpha-kinase domain. Biochem. J. 2009, 420, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Su, L.T.; Agapito, M.A.; Li, M.; Simonson, W.T.; Huttenlocher, A.; Habas, R.; Yue, L.; Runnels, L.W. TRPM7 regulates cell adhesion by controlling the calcium-dependent protease calpain. J. Biol. Chem. 2006, 281, 11260–11270. [Google Scholar] [CrossRef]
- Ryazanova, L.V.; Hu, Z.; Suzuki, S.; Chubanov, V.; Fleig, A.; Ryazanov, A.G. Elucidating the role of the TRPM7 alpha-kinase: TRPM7 kinase inactivation leads to magnesium deprivation resistance phenotype in mice. Sci. Rep. 2014, 4, 7599. [Google Scholar] [CrossRef] [Green Version]
- Chubanov, V.; Waldegger, S.; Mederos y Schnitzler, M.; Vitzthum, H.; Sassen, M.C.; Seyberth, H.W.; Konrad, M.; Gudermann, T. Disruption of TRPM6/TRPM7 complex formation by a mutation in the TRPM6 gene causes hypomagnesemia with secondary hypocalcemia. Proc. Natl. Acad. Sci. USA 2004, 101, 2894–2899. [Google Scholar] [CrossRef] [Green Version]
- Cai, N.; Bai, Z.; Nanda, V.; Runnels, L.W. Mass Spectrometric Analysis of TRPM6 and TRPM7 Phosphorylation Reveals Regulatory Mechanisms of the Channel-Kinases. Sci. Rep. 2017, 7, 42739. [Google Scholar] [CrossRef] [Green Version]
- Deason-Towne, F.; Perraud, A.L.; Schmitz, C. The Mg2+ transporter MagT1 partially rescues cell growth and Mg2+ uptake in cells lacking the channel-kinase TRPM7. FEBS Lett. 2011, 585, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, A.; Moscheni, C.; Trapani, V.; Wolf, F.I.; Farruggia, G.; Sargenti, A.; Iotti, S.; Maier, J.A.; Castiglioni, S. The different expression of TRPM7 and MagT1 impacts on the proliferation of colon carcinoma cells sensitive or resistant to doxorubicin. Sci. Rep. 2017, 7, 40538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goytain, A.; Quamme, G.A. Identification and characterization of a novel mammalian Mg2+ transporter with channel-like properties. BMC Genom. 2005, 6, 48. [Google Scholar] [CrossRef]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef]
- Long, S.; Romani, A.M. Role of Cellular Magnesium in Human Diseases. Austin J. Nutr. Food Sci. 2014, 2, 1051. [Google Scholar]
- Ikari, A.; Sawada, H.; Sanada, A.; Tonegawa, C.; Yamazaki, Y.; Sugatani, J. Magnesium deficiency suppresses cell cycle progression mediated by increase in transcriptional activity of p21(Cip1) and p27(Kip1) in renal epithelial NRK-52E cells. J. Cell Biochem. 2011, 112, 3563–3572. [Google Scholar] [CrossRef]
- MacDonald, R.S. The role of zinc in growth and cell proliferation. J. Nutr. 2000, 130, 1500S–1508S. [Google Scholar] [CrossRef]
- Capiod, T. Cell proliferation, calcium influx and calcium channels. Biochimie 2011, 93, 2075–2079. [Google Scholar] [CrossRef]
- Resende, R.R.; Andrade, L.M.; Oliveira, A.G.; Guimaraes, E.S.; Guatimosim, S.; Leite, M.F. Nucleoplasmic calcium signaling and cell proliferation: Calcium signaling in the nucleus. Cell Commun. Signal 2013, 11, 14. [Google Scholar] [CrossRef]
- Ni, H.M.; Baty, C.J.; Li, N.; Ding, W.X.; Gao, W.; Li, M.; Chen, X.; Ma, J.; Michalopoulos, G.K.; Yin, X.M. Bid agonist regulates murine hepatocyte proliferation by controlling endoplasmic reticulum calcium homeostasis. Hepatology 2010, 52, 338–348. [Google Scholar] [CrossRef]
- Munaron, L. Calcium signalling and control of cell proliferation by tyrosine kinase receptors (review). Int J. Mol. Med. 2002, 10, 671–676. [Google Scholar] [CrossRef]
- Paxton, R.; Ye, L. Regulation of heart insulin receptor tyrosine kinase activity by magnesium and spermine. Mol. Cell Biochem. 2005, 277, 7–17. [Google Scholar] [CrossRef]
- Grubbs, R.D. Effect of epidermal growth factor on magnesium homeostasis in BC3H1 myocytes. Am. J. Physiol. 1991, 260, C1158–C1164. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed]
- Valinsky, W.C.; Jolly, A.; Miquel, P.; Touyz, R.M.; Shrier, A. Aldosterone Upregulates Transient Receptor Potential Melastatin 7 (TRPM7). J. Biol. Chem. 2016, 291, 20163–20172. [Google Scholar] [CrossRef]
- Yogi, A.; Callera, G.E.; O’Connor, S.E.; He, Y.; Correa, J.W.; Tostes, R.C.; Mazur, A.; Touyz, R.M. Dysregulation of renal transient receptor potential melastatin 6/7 but not paracellin-1 in aldosterone-induced hypertension and kidney damage in a model of hereditary hypomagnesemia. J. Hypertens. 2011, 29, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Callera, G.E.; He, Y.; Yogi, A.; Montezano, A.C.; Paravicini, T.; Yao, G.; Touyz, R.M. Regulation of the novel Mg2+ transporter transient receptor potential melastatin 7 (TRPM7) cation channel by bradykinin in vascular smooth muscle cells. J. Hypertens. 2009, 27, 155–166. [Google Scholar] [CrossRef]
- Touyz, R.M.; He, Y.; Montezano, A.C.; Yao, G.; Chubanov, V.; Gudermann, T.; Callera, G.E. Differential regulation of transient receptor potential melastatin 6 and 7 cation channels by ANG II in vascular smooth muscle cells from spontaneously hypertensive rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R73–R78. [Google Scholar] [CrossRef] [Green Version]
- Yogi, A.; Callera, G.E.; Antunes, T.T.; Tostes, R.C.; Touyz, R.M. Transient receptor potential melastatin 7 (TRPM7) cation channels, magnesium and the vascular system in hypertension. Circ. J. Off. J. Jpn. Circ. Soc. 2011, 75, 237–245. [Google Scholar] [CrossRef]
- Montezano, A.C.; Zimmerman, D.; Yusuf, H.; Burger, D.; Chignalia, A.Z.; Wadhera, V.; van Leeuwen, F.N.; Touyz, R.M. Vascular smooth muscle cell differentiation to an osteogenic phenotype involves TRPM7 modulation by magnesium. Hypertension 2010, 56, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Yoon, W.; Shim, J.H.; Jung, H.; Lim, J.H.; Choi, H.J.; Seo, M.; Lee, T.H.; Min, S.H. Role of Protein Kinases and Their Inhibitors in Radiation Response of Tumor Cells. Curr. Pharm. Des. 2017, 23, 4259–4280. [Google Scholar] [CrossRef]
- Schwartz, P.A.; Murray, B.W. Protein kinase biochemistry and drug discovery. Bioor.g Chem. 2011, 39, 192–210. [Google Scholar] [CrossRef]
- Hubbard, S.R.; Miller, W.T. Receptor tyrosine kinases: Mechanisms of activation and signaling. Curr. Opin. Cell Biol. 2007, 19, 117–123. [Google Scholar] [CrossRef]
- Du, Z.; Lovly, C.M. Mechanisms of receptor tyrosine kinase activation in cancer. Mol. Cancer 2018, 17, 58. [Google Scholar] [CrossRef]
- Lee, J.; Miyazaki, M.; Romeo, G.R.; Shoelson, S.E. Insulin receptor activation with transmembrane domain ligands. J. Biol. Chem. 2014, 289, 19769–19777. [Google Scholar] [CrossRef]
- Niu, X.L.; Peters, K.G.; Kontos, C.D. Deletion of the carboxyl terminus of Tie2 enhances kinase activity, signaling, and function. Evidence for an autoinhibitory mechanism. J. Biol. Chem. 2002, 277, 31768–31773. [Google Scholar] [CrossRef]
- Hubbard, S.R. Autoinhibitory mechanisms in receptor tyrosine kinases. Front. Biosci. 2002, 7, d330–d340. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. Src protein-tyrosine kinase structure, mechanism, and small molecule inhibitors. Pharmacol. Res. 2015, 94, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Knighton, D.R.; Ten Eyck, L.F.; Karlsson, R.; Xuong, N.; Taylor, S.S.; Sowadski, J.M. Crystal structure of the catalytic subunit of cAMP-dependent protein kinase complexed with magnesium-ATP and peptide inhibitor. Biochemistry 1993, 32, 2154–2161. [Google Scholar] [CrossRef]
- Bastidas, A.C.; Deal, M.S.; Steichen, J.M.; Guo, Y.; Wu, J.; Taylor, S.S. Phosphoryl transfer by protein kinase A is captured in a crystal lattice. J. Am. Chem. Soc. 2013, 135, 4788–4798. [Google Scholar] [CrossRef] [Green Version]
- Hadari, Y.R.; Gotoh, N.; Kouhara, H.; Lax, I.; Schlessinger, J. Critical role for the docking-protein FRS2 alpha in FGF receptor-mediated signal transduction pathways. Proc. Natl. Acad. Sci. USA 2001, 98, 8578–8583. [Google Scholar] [CrossRef]
- Fraser, J.; Cabodevilla, A.G.; Simpson, J.; Gammoh, N. Interplay of autophagy, receptor tyrosine kinase signalling and endocytic trafficking. Essays Biochem. 2017, 61, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Arkun, Y.; Yasemi, M. Dynamics and control of the ERK signaling pathway: Sensitivity, bistability, and oscillations. PLoS ONE 2018, 13, e0195513. [Google Scholar] [CrossRef]
- Soares-Silva, M.; Diniz, F.F.; Gomes, G.N.; Bahia, D. The Mitogen-Activated Protein Kinase (MAPK) Pathway: Role in Immune Evasion by Trypanosomatids. Front. Microbiol. 2016, 7, 183. [Google Scholar] [CrossRef]
- Annenkov, A. Receptor tyrosine kinase (RTK) signalling in the control of neural stem and progenitor cell (NSPC) development. Mol. Neurobiol. 2014, 49, 440–471. [Google Scholar] [CrossRef]
- Papadimitrakopoulou, V. Development of PI3K/AKT/mTOR pathway inhibitors and their application in personalized therapy for non-small-cell lung cancer. J. Thorac. Oncol. 2012, 7, 1315–1326. [Google Scholar] [CrossRef]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef]
- Paquette, M.; El-Houjeiri, L.; Pause, A. mTOR Pathways in Cancer and Autophagy. Cancers (Basel) 2018, 10, 18. [Google Scholar] [CrossRef]
- Chang, L.; Graham, P.H.; Ni, J.; Hao, J.; Bucci, J.; Cozzi, P.J.; Li, Y. Targeting PI3K/Akt/mTOR signaling pathway in the treatment of prostate cancer radioresistance. Crit. Rev. Oncol. Hematol. 2015, 96, 507–517. [Google Scholar] [CrossRef]
- Rhee, S.G. Regulation of phosphoinositide-specific phospholipase C. Annu. Rev. Biochem. 2001, 70, 281–312. [Google Scholar] [CrossRef]
- Yang, Y.R.; Follo, M.Y.; Cocco, L.; Suh, P.G. The physiological roles of primary phospholipase C. Adv. Biol. Regul. 2013, 53, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Putney, J.W.; Tomita, T. Phospholipase C signaling and calcium influx. Adv. Biol. Regul. 2012, 52, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Mikoshiba, K. IP3 receptor/Ca2+ channel: From discovery to new signaling concepts. J. Neurochem. 2007, 102, 1426–1446. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Inanobe, S.; Kanemaru, K.; Nakamura, Y. Phospholipase C is a key enzyme regulating intracellular calcium and modulating the phosphoinositide balance. Prog. Lipid Res. 2010, 49, 429–437. [Google Scholar] [CrossRef]
- Zhu, M.; Leung, C.Y.; Shahbazi, M.N.; Zernicka-Goetz, M. Actomyosin polarisation through PLC-PKC triggers symmetry breaking of the mouse embryo. Nat. Commun. 2017, 8, 921. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zheng, D.; Cui, H.; Liu, S.; Zhang, L.; Liu, C. Roles and mechanisms of TRPC3 and the PLCgamma/PKC/CPI-17 signaling pathway in regulating parturition. Mol. Med. Rep. 2018, 17, 898–910. [Google Scholar]
- Yu, M.; Chen, Y.; Zeng, H.; Zheng, Y.; Fu, G.; Zhu, W.; Broeckel, U.; Aggarwal, P.; Turner, A.; Neale, G.; et al. PLCgamma-dependent mTOR signalling controls IL-7-mediated early B cell development. Nat. Commun. 2017, 8, 1457. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.A. The Jak/STAT pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011205. [Google Scholar] [CrossRef]
- Izzedine, H.; Perazella, M.A. Adverse kidney effects of epidermal growth factor receptor inhibitors. Nephrol. Dial. Transpl. 2017, 32, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.S.; Khankin, E.V.; Karumanchi, S.A.; Humphreys, B.D. Hypertension induced by vascular endothelial growth factor signaling pathway inhibition: Mechanisms and potential use as a biomarker. Semin. Nephrol. 2010, 30, 591–601. [Google Scholar] [CrossRef]
- Mouhayar, E.; Salahudeen, A. Hypertension in cancer patients. Tex. Heart Inst. J. 2011, 38, 263–265. [Google Scholar]
- Fakih, M.; Vincent, M. Adverse events associated with anti-EGFR therapies for the treatment of metastatic colorectal cancer. Curr. Oncol. 2010, 17 (Suppl. 1), S18–S30. [Google Scholar] [CrossRef]
- Cosmai, L.; Gallieni, M.; Porta, C. Renal toxicity of anticancer agents targeting HER2 and EGFR. J. Nephrol. 2015, 28, 647–657. [Google Scholar] [CrossRef]
- Hong, B.Z.; Kang, H.S.; So, J.N.; Kim, H.N.; Park, S.A.; Kim, S.J.; Kim, K.R.; Kwak, Y.G. Vascular endothelial growth factor increases the intracellular magnesium. Biochem. Biophys. Res. Commun. 2006, 347, 496–501. [Google Scholar] [CrossRef]
- Jalkanen, R.; Pronicka, E.; Tyynismaa, H.; Hanauer, A.; Walder, R.; Alitalo, T. Genetic background of HSH in three Polish families and a patient with an X;9 translocation. Eur. J. Hum. Genet. 2006, 14, 55–62. [Google Scholar] [CrossRef]
- Runnels, L.W.; Yue, L.; Clapham, D.E. The TRPM7 channel is inactivated by PIP(2) hydrolysis. Nat. Cell Biol. 2002, 4, 329–336. [Google Scholar] [CrossRef]
- Gao, H.; Chen, X.; Du, X.; Guan, B.; Liu, Y.; Zhang, H. EGF enhances the migration of cancer cells by up-regulation of TRPM7. Cell Calcium 2011, 50, 559–568. [Google Scholar] [CrossRef]
- Tian, S.L.; Jiang, H.; Zeng, Y.; Li, L.L.; Shi, J. NGF-induced reduction of an outward-rectifying TRPM7-like current in rat CA1 hippocampal neurons. Neurosci. Lett. 2007, 419, 93–98. [Google Scholar] [CrossRef]
- Jiang, H.; Tian, S.L.; Zeng, Y.; Li, L.L.; Shi, J. TrkA pathway(s) is involved in regulation of TRPM7 expression in hippocampal neurons subjected to ischemic-reperfusion and oxygen-glucose deprivation. Brain Res. Bull. 2008, 76, 124–130. [Google Scholar] [CrossRef]
- Fang, L.; Zhan, S.; Huang, C.; Cheng, X.; Lv, X.; Si, H.; Li, J. TRPM7 channel regulates PDGF-BB-induced proliferation of hepatic stellate cells via PI3K and ERK pathways. Toxicol. Appl. Pharmacol. 2013, 272, 713–725. [Google Scholar] [CrossRef]
- Langeslag, M.; Clark, K.; Moolenaar, W.H.; van Leeuwen, F.N.; Jalink, K. Activation of TRPM7 channels by phospholipase C-coupled receptor agonists. J. Biol. Chem. 2007, 282, 232–239. [Google Scholar] [CrossRef]
- Sun, Y.; Sukumaran, P.; Varma, A.; Derry, S.; Sahmoun, A.E.; Singh, B.B. Cholesterol-induced activation of TRPM7 regulates cell proliferation, migration, and viability of human prostate cells. Biochim. Biophys. Acta 2014, 1843, 1839–1850. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, M.; Fan, X.H.; Chen, J.H.; Guan, Y.Y.; Tang, Y.B. Upregulation of TRPM7 channels by angiotensin II triggers phenotypic switching of vascular smooth muscle cells of ascending aorta. Circ. Res. 2012, 111, 1137–1146. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Feng, W.; Chen, R.; Chen, J.; Touyz, R.M.; Wang, J.; Huang, H. Interleukin-18 Enhances Vascular Calcification and Osteogenic Differentiation of Vascular Smooth Muscle Cells Through TRPM7 Activation. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1933–1943. [Google Scholar] [CrossRef]
- Liu, A.; Zhao, F.; Wang, J.; Zhao, Y.; Luo, Z.; Gao, Y.; Shi, J. Regulation of TRPM7 Function by IL-6 through the JAK2-STAT3 Signaling Pathway. PLoS ONE 2016, 11, e0152120. [Google Scholar] [CrossRef]
- Inoue, K.; Xiong, Z.G. Silencing TRPM7 promotes growth/proliferation and nitric oxide production of vascular endothelial cells via the ERK pathway. Cardiovasc. Res. 2009, 83, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Su, L.T.; Chen, H.C.; Gonzalez-Pagan, O.; Overton, J.D.; Xie, J.; Yue, L.; Runnels, L.W. TRPM7 activates m-calpain by stress-dependent stimulation of p38 MAPK and c-Jun N-terminal kinase. J. Mol. Biol. 2010, 396, 858–869. [Google Scholar] [CrossRef]
- Zeng, Z.; Leng, T.; Feng, X.; Sun, H.; Inoue, K.; Zhu, L.; Xiong, Z.G. Silencing TRPM7 in mouse cortical astrocytes impairs cell proliferation and migration via ERK and JNK signaling pathways. PLoS ONE 2015, 10, e0119912. [Google Scholar] [CrossRef]
- Lin, J.; Zhou, S.; Zhao, T.; Ju, T.; Zhang, L. TRPM7 channel regulates ox-LDL-induced proliferation and migration of vascular smooth muscle cells via MEK-ERK pathways. FEBS Lett. 2016, 590, 520–532. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Cai, C.; Wu, J.; Cai, S.; Ye, C.; Chen, H.; Yang, Z.; Zeng, H.; Shen, Q.; Zou, F. TRPM7 mediates breast cancer cell migration and invasion through the MAPK pathway. Cancer Lett. 2013, 333, 96–102. [Google Scholar] [CrossRef]
- Davis, F.M.; Azimi, I.; Faville, R.A.; Peters, A.A.; Jalink, K.; Putney, J.W., Jr.; Goodhill, G.J.; Thompson, E.W.; Roberts-Thomson, S.J.; Monteith, G.R. Induction of epithelial-mesenchymal transition (EMT) in breast cancer cells is calcium signal dependent. Oncogene 2014, 33, 2307–2316. [Google Scholar] [CrossRef]
- Wong, R.; Turlova, E.; Feng, Z.P.; Rutka, J.T.; Sun, H.S. Activation of TRPM7 by naltriben enhances migration and invasion of glioblastoma cells. Oncotarget 2017, 8, 11239–11248. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.L.; Barszczyk, A.; Turlova, E.; Deurloo, M.; Liu, B.; Yang, B.B.; Rutka, J.T.; Feng, Z.P.; Sun, H.S. Inhibition of TRPM7 by carvacrol suppresses glioblastoma cell proliferation, migration and invasion. Oncotarget 2015, 6, 16321–16340. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Wu, J.Y.; Lu, M.H.; Shi, Z.; Na, N.; Di, J.M. Carvacrol Alleviates Prostate Cancer Cell Proliferation, Migration, and Invasion through Regulation of PI3K/Akt and MAPK Signaling Pathways. Oxid. Med. Cell. Longev. 2016, 2016, 1469693. [Google Scholar] [CrossRef]
- Lu, D.; Qu, J.; Sun, L.; Li, Q.; Ling, H.; Yang, N.; Ma, T.; Wang, Q.; Li, M.; Zhang, K.; et al. Ca2+/Mg2+ homeostasisrelated TRPM7 channel mediates chondrocyte hypertrophy via regulation of the PI3KAkt signaling pathway. Mol. Med. Rep. 2017, 16, 5699–5705. [Google Scholar] [CrossRef]
- Liu, M.; Inoue, K.; Leng, T.; Guo, S.; Xiong, Z.G. TRPM7 channels regulate glioma stem cell through STAT3 and Notch signaling pathways. Cell Signal 2014, 26, 2773–2781. [Google Scholar] [CrossRef] [Green Version]
- Saltarella, I.; Frassanito, M.A.; Lamanuzzi, A.; Brevi, A.; Leone, P.; Desantis, V.; Di Marzo, L.; Bellone, M.; Derudas, D.; Ribatti, D.; et al. Homotypic and Heterotypic Activation of the Notch Pathway in Multiple Myeloma-Enhanced Angiogenesis: A Novel Therapeutic Target? Neoplasia 2019, 21, 93–105. [Google Scholar] [CrossRef]
- Wang, W.M.; Zhao, Z.L.; Ma, S.R.; Yu, G.T.; Liu, B.; Zhang, L.; Zhang, W.F.; Kulkarni, A.B.; Sun, Z.J.; Zhao, Y.F. Epidermal growth factor receptor inhibition reduces angiogenesis via hypoxia-inducible factor-1alpha and Notch1 in head neck squamous cell carcinoma. PLoS ONE 2015, 10, e0119723. [Google Scholar]
- Romani, A.M. Cellular magnesium homeostasis. Arch. Biochem. Biophys. 2011, 512, 1–23. [Google Scholar] [CrossRef]
- Al Alawi, A.M.; Majoni, S.W.; Falhammar, H. Magnesium and Human Health: Perspectives and Research Directions. Int. J. Endocrinol. 2018, 2018, 9041694. [Google Scholar] [CrossRef]
- Jin, J.; Wu, L.J.; Jun, J.; Cheng, X.; Xu, H.; Andrews, N.C.; Clapham, D.E. The channel kinase, TRPM7, is required for early embryonic development. Proc. Natl. Acad. Sci. USA 2012, 109, E225–E233. [Google Scholar] [CrossRef]
- Liu, Y.S.; Liu, Y.A.; Huang, C.J.; Yen, M.H.; Tseng, C.T.; Chien, S.; Lee, O.K. Mechanosensitive TRPM7 mediates shear stress and modulates osteogenic differentiation of mesenchymal stromal cells through Osterix pathway. Sci. Rep. 2015, 5, 16522. [Google Scholar] [CrossRef] [Green Version]
- Ogunrinde, A.; Pereira, R.D.; Beaton, N.; Lam, D.H.; Whetstone, C.; Hill, C.E. Hepatocellular differentiation status is characterized by distinct subnuclear localization and form of the chanzyme TRPM7. Differentiation 2017, 96, 15–25. [Google Scholar] [CrossRef]
- Yu, M.; Huang, C.; Huang, Y.; Wu, X.; Li, X.; Li, J. Inhibition of TRPM7 channels prevents proliferation and differentiation of human lung fibroblasts. Inflamm. Res. 2013, 62, 961–970. [Google Scholar] [CrossRef]
- Cui, L.; Xu, S.M.; Ma, D.D.; Wu, B.L. The effect of TRPM7 suppression on the proliferation, migration and osteogenic differentiation of human dental pulp stem cells. Int. Endod. J. 2014, 47, 583–593. [Google Scholar] [CrossRef]
- Zhang, X.; Zu, H.; Zhao, D.; Yang, K.; Tian, S.; Yu, X.; Lu, F.; Liu, B.; Yu, X.; Wang, B.; et al. Ion channel functional protein kinase TRPM7 regulates Mg ions to promote the osteoinduction of human osteoblast via PI3K pathway: In vitro simulation of the bone-repairing effect of Mg-based alloy implant. Acta Biomater. 2017, 63, 369–382. [Google Scholar] [CrossRef]
- Takahashi, K.; Umebayashi, C.; Numata, T.; Honda, A.; Ichikawa, J.; Hu, Y.; Yamaura, K.; Inoue, R. TRPM7-mediated spontaneous Ca(2+) entry regulates the proliferation and differentiation of human leukemia cell line K562. Physiol. Rep. 2018, 6, e13796. [Google Scholar] [CrossRef]
- Yang, M.; Fang, J.; Liu, Q.; Wang, Y.; Zhang, Z. Role of ROS-TRPM7-ERK1/2 axis in high concentration glucose-mediated proliferation and phenotype switching of rat aortic vascular smooth muscle cells. Biochem. Biophys. Res. Commun. 2017, 494, 526–533. [Google Scholar] [CrossRef]
- Yee, N.S.; Zhou, W.; Liang, I.C. Transient receptor potential ion channel Trpm7 regulates exocrine pancreatic epithelial proliferation by Mg2+-sensitive Socs3a signaling in development and cancer. Dis. Model. Mech. 2011, 4, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef]
- Liu, K.; Xu, S.H.; Chen, Z.; Zeng, Q.X.; Li, Z.J.; Chen, Z.M. TRPM7 overexpression enhances the cancer stem cell-like and metastatic phenotypes of lung cancer through modulation of the Hsp90alpha/uPA/MMP2 signaling pathway. BMC Cancer 2018, 18, 1167. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wu, N.; Wang, Y.; Zhang, X.; Xia, B.; Tang, J.; Cai, J.; Zhao, Z.; Liao, Q.; Wang, J. TRPM7 promotes the epithelial-mesenchymal transition in ovarian cancer through the calcium-related PI3K / AKT oncogenic signaling. J. Exp. Clin. Cancer Res. 2019, 38, 106. [Google Scholar] [CrossRef]
- Xiao, E.; Yang, H.Q.; Gan, Y.H.; Duan, D.H.; He, L.H.; Guo, Y.; Wang, S.Q.; Zhang, Y. Brief reports: TRPM7 Senses mechanical stimulation inducing osteogenesis in human bone marrow mesenchymal stem cells. Stem Cells 2015, 33, 615–621. [Google Scholar] [CrossRef]
- Cheng, H.; Feng, J.M.; Figueiredo, M.L.; Zhang, H.; Nelson, P.L.; Marigo, V.; Beck, A. Transient receptor potential melastatin type 7 channel is critical for the survival of bone marrow derived mesenchymal stem cells. Stem Cells Dev. 2010, 19, 1393–1403. [Google Scholar] [CrossRef]
- Diaz-Tocados, J.M.; Herencia, C.; Martinez-Moreno, J.M.; Montes de Oca, A.; Rodriguez-Ortiz, M.E.; Vergara, N.; Blanco, A.; Steppan, S.; Almaden, Y.; Rodriguez, M.; et al. Magnesium Chloride promotes Osteogenesis through Notch signaling activation and expansion of Mesenchymal Stem Cells. Sci. Rep. 2017, 7, 7839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, N.S. Role of TRPM7 in Cancer: Potential as Molecular Biomarker and Therapeutic Target. Pharmaceuticals (Basel) 2017, 10, 39. [Google Scholar] [CrossRef]
- Yang, H.; Kim, T.H.; Lee, G.S.; Hong, E.J.; Jeung, E.B. Comparing the expression patterns of placental magnesium/phosphorus-transporting channels between healthy and preeclamptic pregnancies. Mol. Reprod. Dev. 2014, 81, 851–860. [Google Scholar] [CrossRef]
- Weintraub, A.Y.; Amash, A.; Eshkoli, T.; Piltcher Haber, E.; Bronfenmacher, B.; Sheiner, E.; Holcberg, G.; Huleihel, M. The effects of magnesium sulfate on placental vascular endothelial growth factor expression in preeclampsia. Hypertens. Pregnancy 2013, 32, 178–188. [Google Scholar] [CrossRef]
- Costa, A.; Tejpar, S.; Prenen, H.; Van Cutsem, E. Hypomagnesaemia and targeted anti-epidermal growth factor receptor (EGFR) agents. Target. Oncol. 2011, 6, 227–233. [Google Scholar] [CrossRef]
Figure 1.
Schematic structure of the TRPM7 channel and kinase. The general structure of the TRPM7 has four Melastatin Homologous Regions (MHR) in the N-terminal domain, and six transmembrane segments (aa 756–1095). The pore (aa 1039–1056) is located between the S5 and S6. The C terminal domain contains the transient receptor potential (TRP) region (aa 1109–1128), common to TRP family members, followed by the coiled coil (CC) domain connecting loop (aa 1198–1250); Serine- and threonine-rich domains (aa 1380–1596), and the α-kinase domain (aa 1597–1821).
Figure 1.
Schematic structure of the TRPM7 channel and kinase. The general structure of the TRPM7 has four Melastatin Homologous Regions (MHR) in the N-terminal domain, and six transmembrane segments (aa 756–1095). The pore (aa 1039–1056) is located between the S5 and S6. The C terminal domain contains the transient receptor potential (TRP) region (aa 1109–1128), common to TRP family members, followed by the coiled coil (CC) domain connecting loop (aa 1198–1250); Serine- and threonine-rich domains (aa 1380–1596), and the α-kinase domain (aa 1597–1821).

Figure 2.
Schematic overview of TRPM7 mutations. Mutations that have been identified are highlighted. Numbers indicate the single amino acids or regions that affect TRPM7 channel or kinase activity [17,19,46,48,49,54,55,56,57,58,59].

Figure 3.
Transphosphorylation of TRPM7 by TRPM6. TRPM6 is able to induce phosphorylation of TRPM7 in the indicated residues. Red stars indicate the location of the residues. Residues in blue are also residues of autophosphorylation.
Figure 3.
Transphosphorylation of TRPM7 by TRPM6. TRPM6 is able to induce phosphorylation of TRPM7 in the indicated residues. Red stars indicate the location of the residues. Residues in blue are also residues of autophosphorylation.

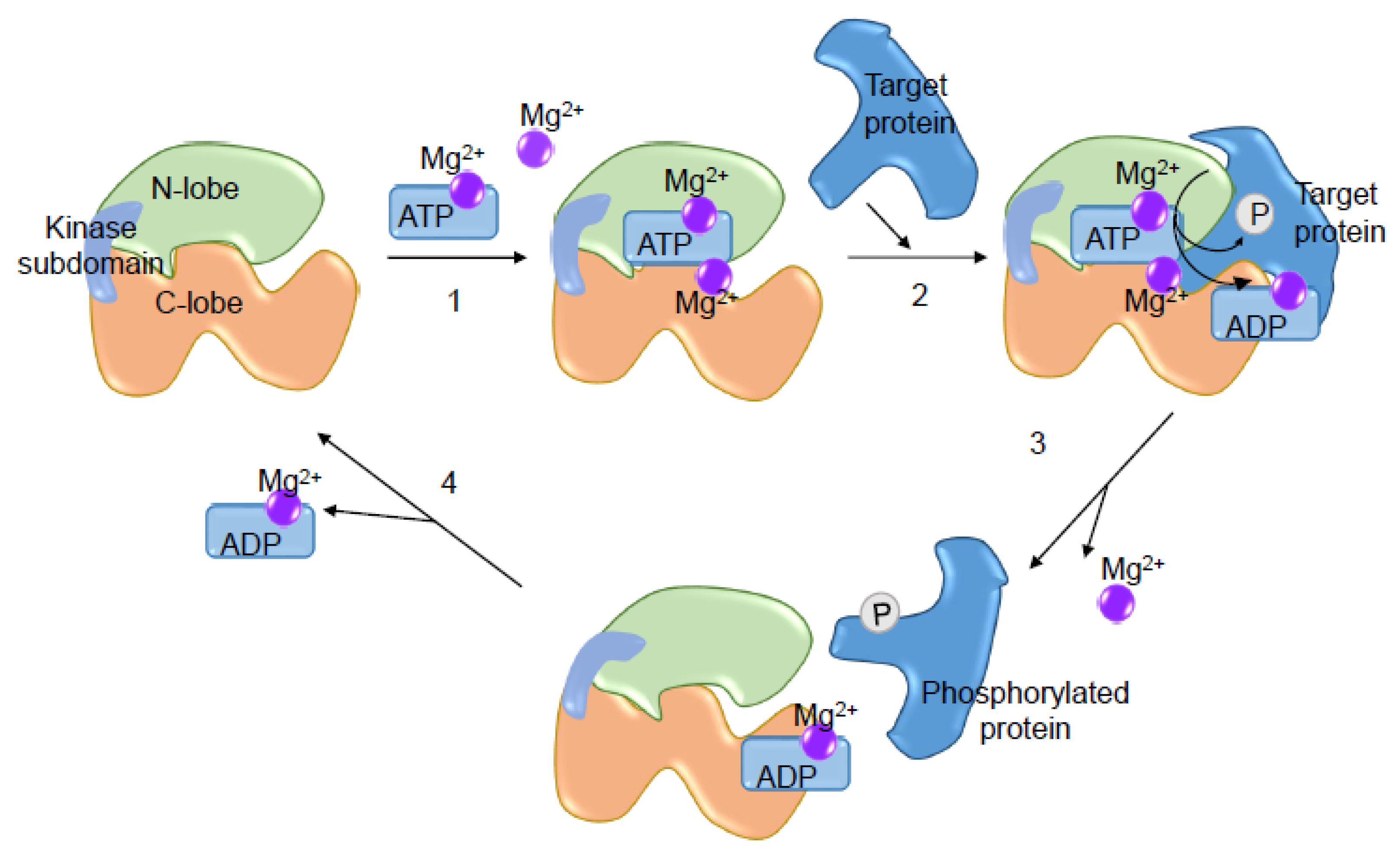
Figure 4.
Importance of Mg2+ for the kinase catalytic activity. Two Mg2+ molecules are required for enzymatic activity and phosphoryl transfer: one bound to ATP (Mg-ATP) situated between the small (N-lobe) and large lobe (C-Lobe) of the kinase domain, and another Mg2+ situated in the catalytic loop. (1) Mg-ATP is the first to bind to the enzyme followed by Mg2+; (2) the kinase binds to the protein substrate and catalyzes the transfer the phosphoryl group; (3) phosphorylated protein and Mg2+ are released; and (4) Mg-ADP is released and the catalytic cycle is finalized.
Figure 4.
Importance of Mg2+ for the kinase catalytic activity. Two Mg2+ molecules are required for enzymatic activity and phosphoryl transfer: one bound to ATP (Mg-ATP) situated between the small (N-lobe) and large lobe (C-Lobe) of the kinase domain, and another Mg2+ situated in the catalytic loop. (1) Mg-ATP is the first to bind to the enzyme followed by Mg2+; (2) the kinase binds to the protein substrate and catalyzes the transfer the phosphoryl group; (3) phosphorylated protein and Mg2+ are released; and (4) Mg-ADP is released and the catalytic cycle is finalized.

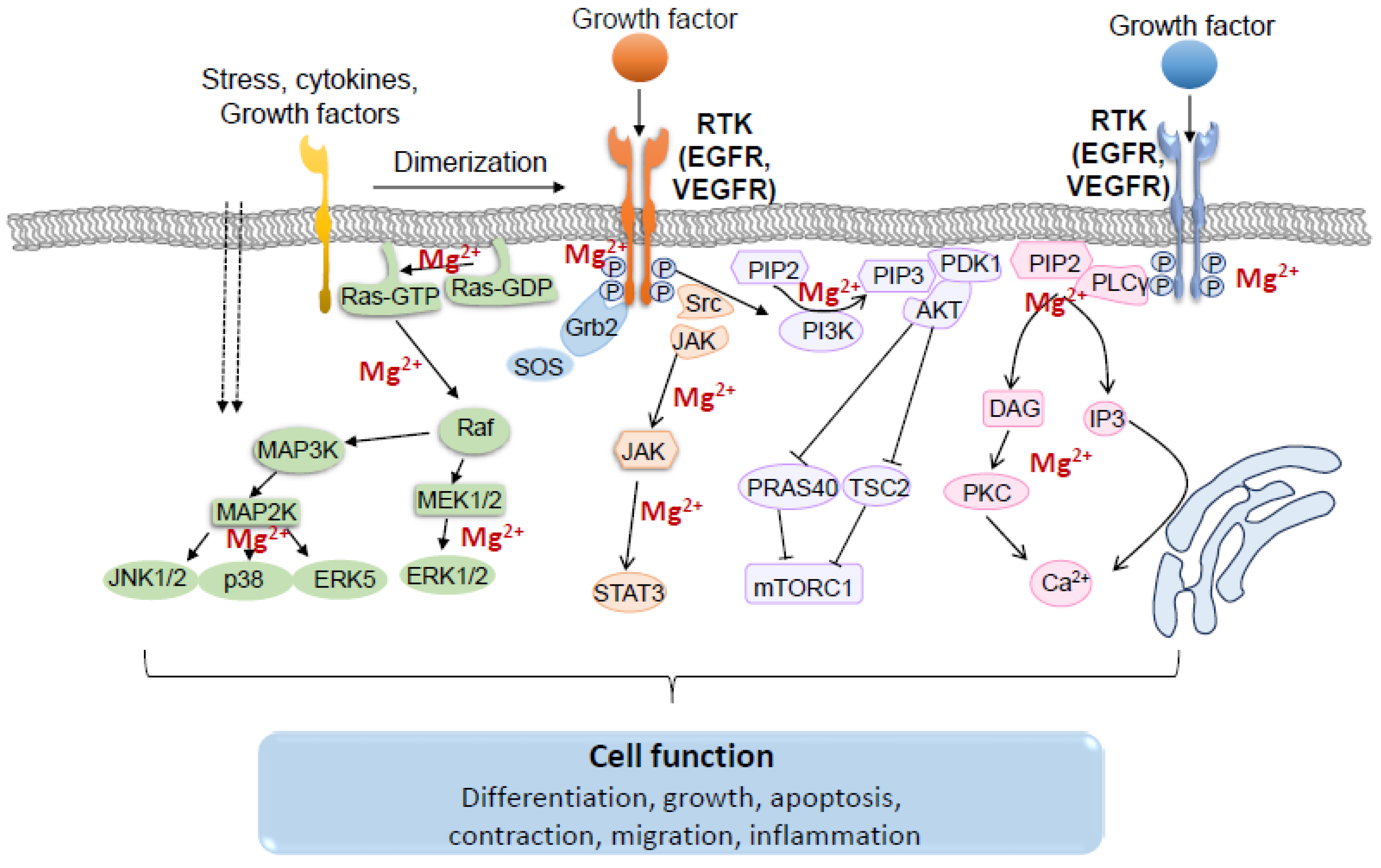
Figure 5.
Schematic representation of main signaling pathways activated by growth factor signaling through receptor tyrosine kinases (RTK) and the importance of Mg2+. Ligand-induced dimerization triggers transphosphorylation of tyrosine residues located in the receptor chain, resulting in activation of RTKs. Intracellular signaling cascades activated by RTKs include the MAPK pathway, the PI3K/AKT/mTOR pathway, the JAK/STAT pathway, the PLCγ/PKC pathway and the Src pathway. The functional response of these signals plays an essential role in the regulation of many physiological processes. Dotted arrows indicate activation through the cell membrane; Solid arrows indicate activation of target protein; T indicates inhibition.
Figure 5.
Schematic representation of main signaling pathways activated by growth factor signaling through receptor tyrosine kinases (RTK) and the importance of Mg2+. Ligand-induced dimerization triggers transphosphorylation of tyrosine residues located in the receptor chain, resulting in activation of RTKs. Intracellular signaling cascades activated by RTKs include the MAPK pathway, the PI3K/AKT/mTOR pathway, the JAK/STAT pathway, the PLCγ/PKC pathway and the Src pathway. The functional response of these signals plays an essential role in the regulation of many physiological processes. Dotted arrows indicate activation through the cell membrane; Solid arrows indicate activation of target protein; T indicates inhibition.

Figure 6.
Cross-talk between Receptor Tyrosine Kinases (RTK) downstream signaling and TRPM7. TRPM7 has the dual properties of acting as an ion channel mainly permeable to Zn2+, Mg2+ and Ca2+, and as a cytoplasmic kinase, which phosphorylates annexin-1, myosin IIA heavy chain, eEF2, SMAD2, and PLCγ2. Through either its channel or kinase, TRPM7 participates in RTK downstream signaling pathways. On the other hand, ligand induced activation of RTKs, such as EGFR, regulates the activity of TRPM7 or TRPM6. Signaling by bradykinin, Ang II, and IL-18 also influence TRPM7 activity. Dotted arrows indicate activation through the cell membrane; Solid arrows indicate activation of target protein; T indicate inhibition.
Figure 6.
Cross-talk between Receptor Tyrosine Kinases (RTK) downstream signaling and TRPM7. TRPM7 has the dual properties of acting as an ion channel mainly permeable to Zn2+, Mg2+ and Ca2+, and as a cytoplasmic kinase, which phosphorylates annexin-1, myosin IIA heavy chain, eEF2, SMAD2, and PLCγ2. Through either its channel or kinase, TRPM7 participates in RTK downstream signaling pathways. On the other hand, ligand induced activation of RTKs, such as EGFR, regulates the activity of TRPM7 or TRPM6. Signaling by bradykinin, Ang II, and IL-18 also influence TRPM7 activity. Dotted arrows indicate activation through the cell membrane; Solid arrows indicate activation of target protein; T indicate inhibition.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zou, Z.-G.; Rios, F.J.; Montezano, A.C.; Touyz, R.M. TRPM7, Magnesium, and Signaling. Int. J. Mol. Sci. 2019, 20, 1877. https://doi.org/10.3390/ijms20081877
AMA Style
Zou Z-G, Rios FJ, Montezano AC, Touyz RM. TRPM7, Magnesium, and Signaling. International Journal of Molecular Sciences. 2019; 20(8):1877. https://doi.org/10.3390/ijms20081877
Chicago/Turabian StyleZou, Zhi-Guo, Francisco J. Rios, Augusto C. Montezano, and Rhian M. Touyz. 2019. "TRPM7, Magnesium, and Signaling" International Journal of Molecular Sciences 20, no. 8: 1877. https://doi.org/10.3390/ijms20081877
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.