SAMHD1 Enhances Chikungunya and Zika Virus Replication in Human Skin Fibroblasts

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
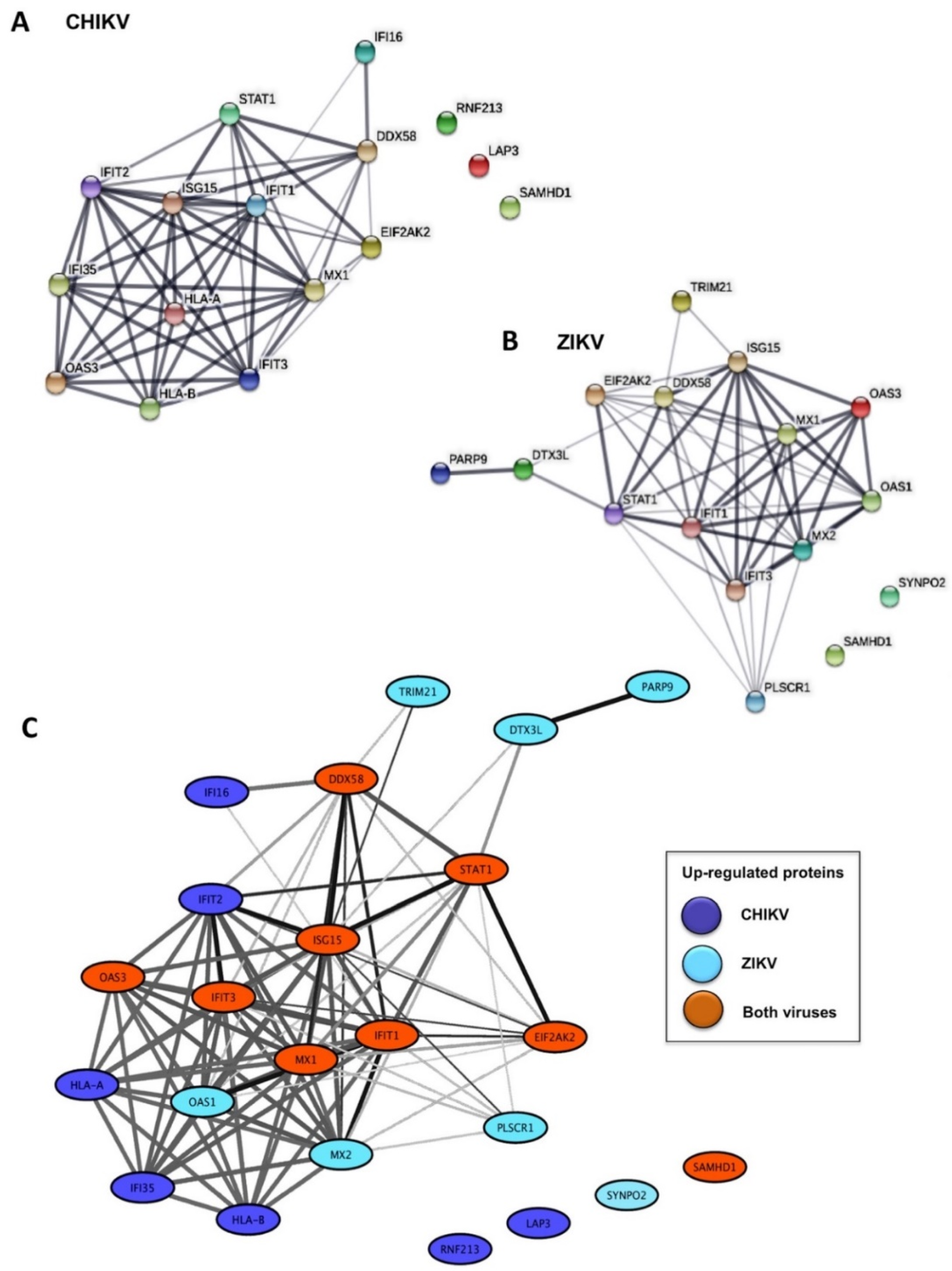
2.1. Identification of Differentially Expressed Proteins in Infected HFF1 Cells
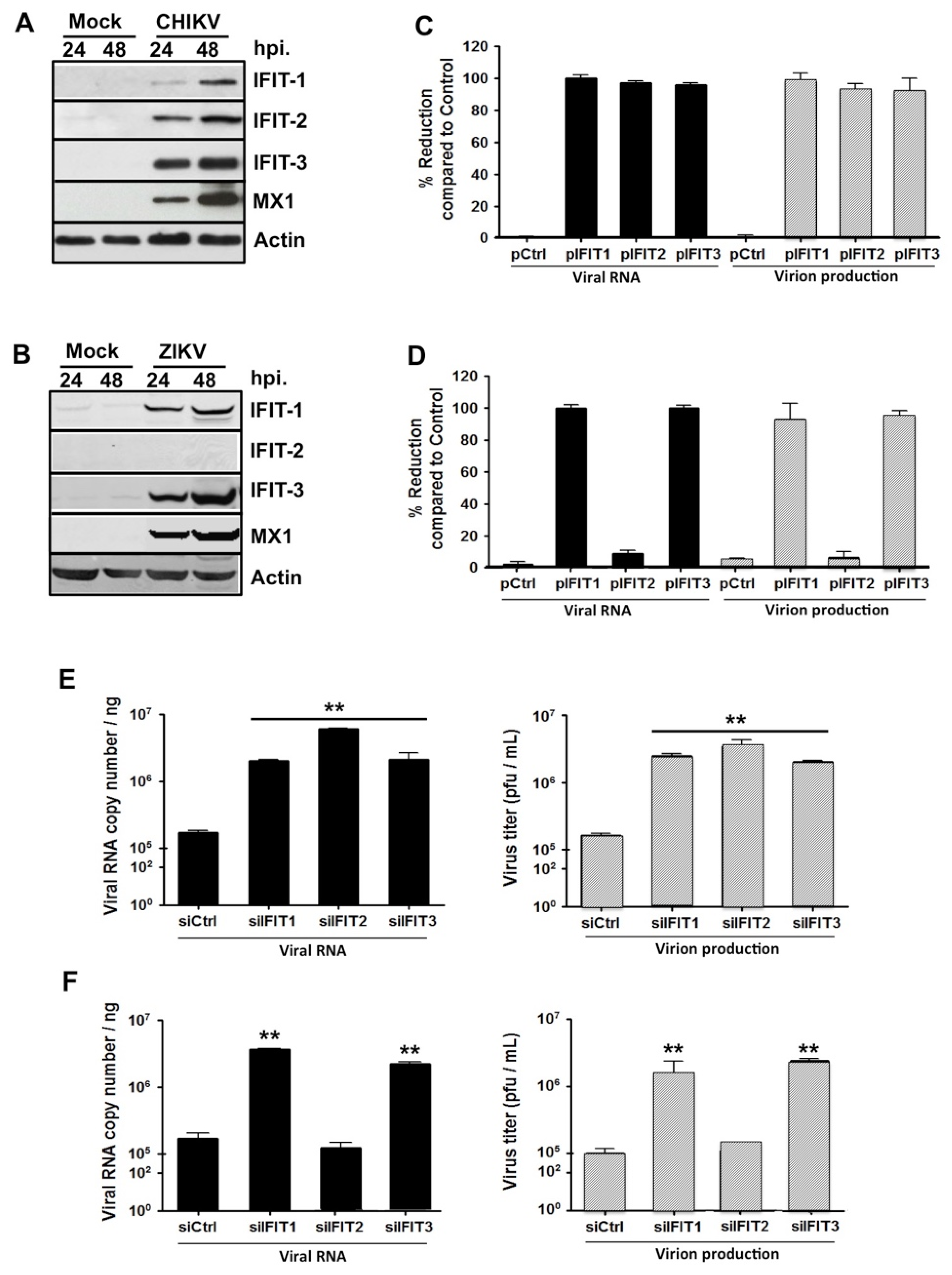
2.2. Expression of IFIT Proteins Affects CHIKV and ZIKV Replication in HFF1 Cells
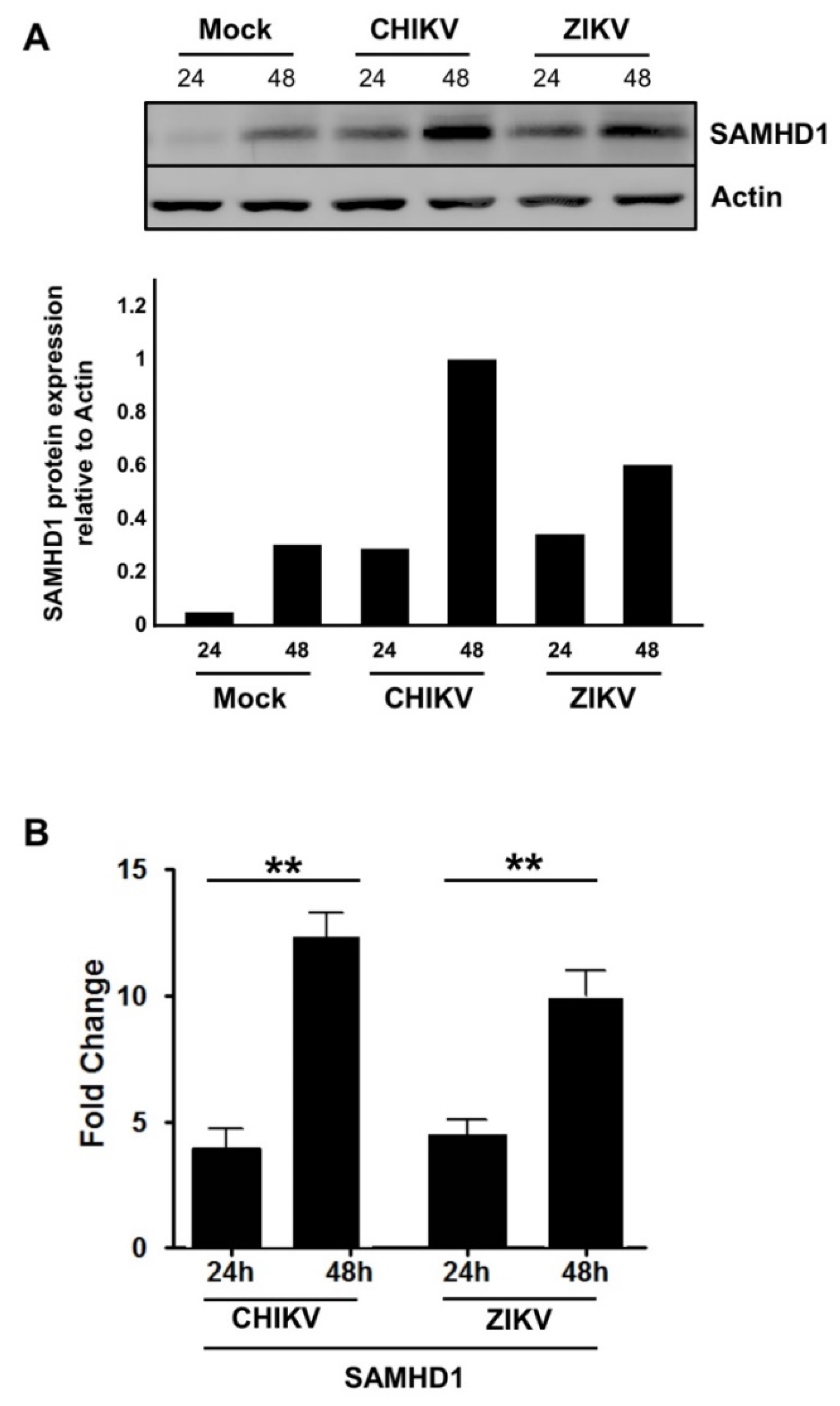
2.3. SAMHD1 Expression Is Induced in Human Skin Fibroblasts Following CHIKV or ZIKV Infection
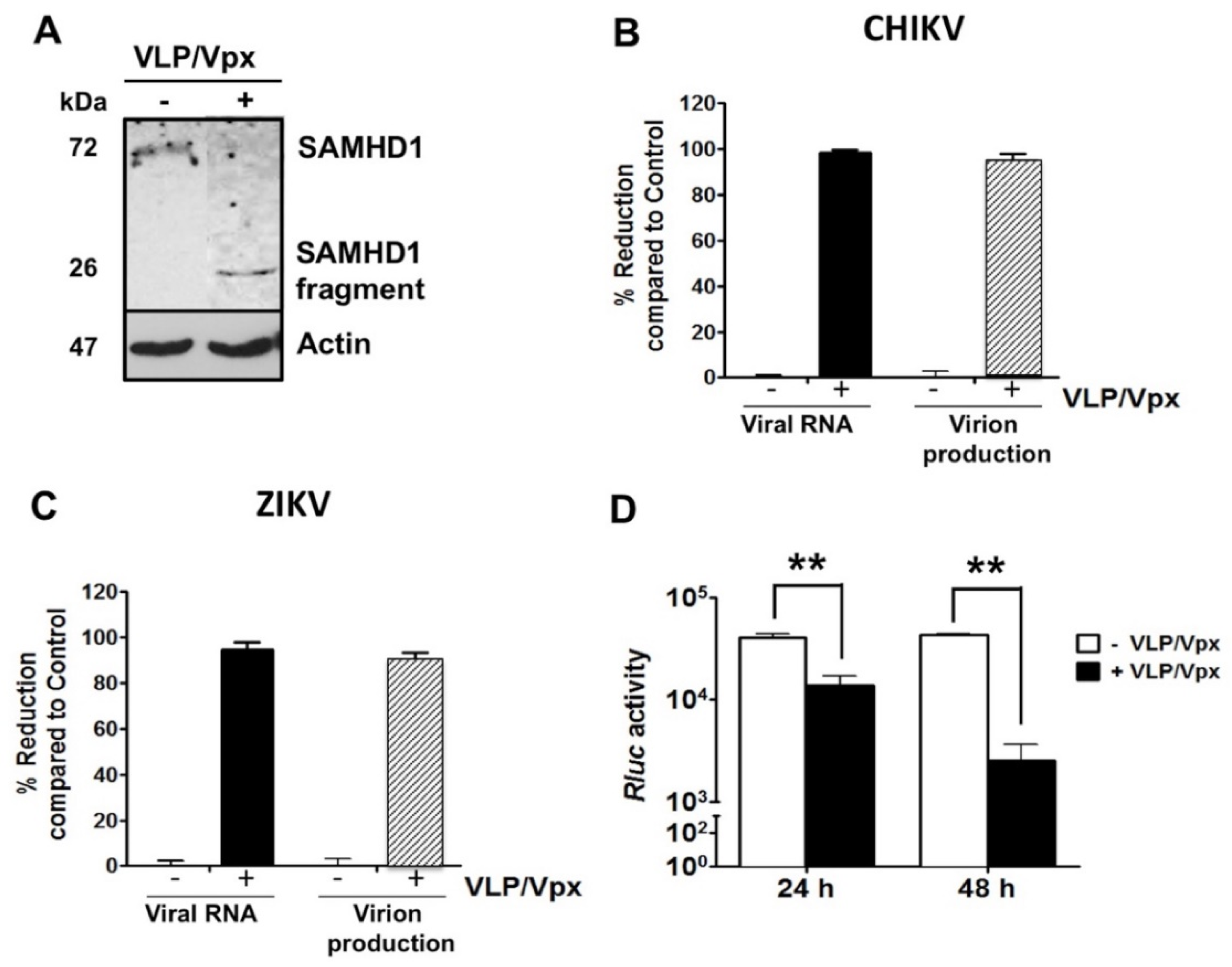
2.4. HIV-2 Vpx Protein-Induced SAMHD1 Degradation Inhibits CHIKV and ZIKV Replication
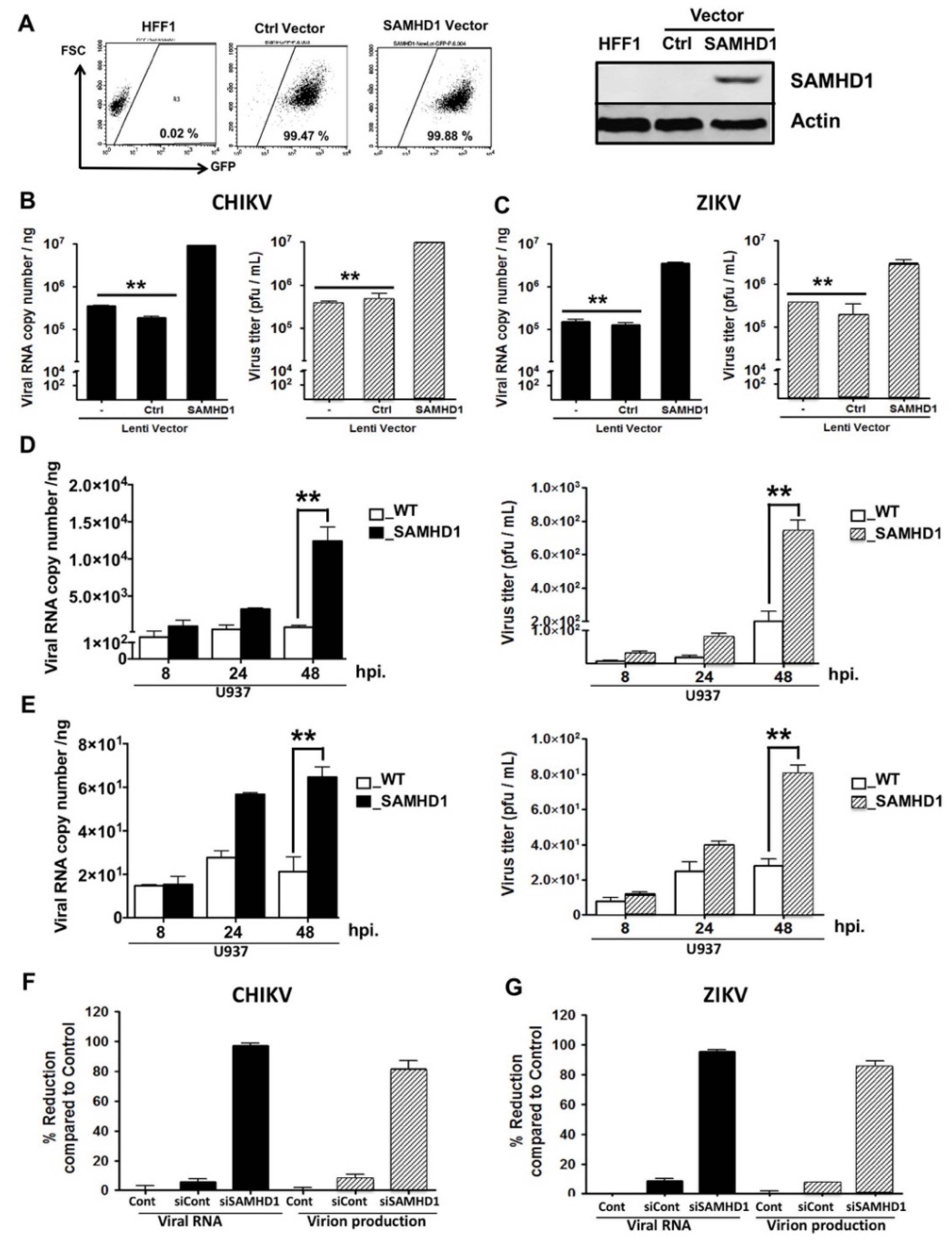
2.5. SAMHD1 Enhances Both CHIKV and ZIKV Replication in HFF1 Cells
3. Discussion
4. Materials and Methods
4.1. Cells and Viruses
4.2. Antibodies, Plasmids and Reagents
4.3. SILAC Labeling and Virus Infection
4.4. Mass Spectrometry Analysis
4.5. Bioinformatics Analysis
4.6. VLPs/Vpx Treatment
4.7. Infection
4.8. Small Interfering RNA (siRNA) Transfection
4.9. Generation of SAMHD1-Expressing Cell Line
4.10. Western Blotting Analysis
4.11. Viral RNA Quantification and Gene Expression by Real Time RT-PCR
4.12. CHIKV Replicon Cell Line Based Assay
4.13. Plaque Assay
4.14. Data Analysis and Statistical Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khan, K.; Bogoch, I.; Brownstein, J.S.; Miniota, J.; Nicolucci, A.; Hu, W.; Nsoesie, E.O.; Cetron, M.; Creatore, M.I.; German, M.; et al. Assessing the origin of and potential for international spread of chikungunya virus from the Caribbean. PLoS Curr. Influenza 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 2015, 372, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.E. Reemergence of chikungunya virus. J. Virol. 2014, 88, 11644–11647. [Google Scholar] [CrossRef]
- Pellot, A.S.; Alessandri, J.L.; Robin, S.; Sampériz, S.; Attali, T.; Brayer, C.; Pasquet, M.; Jaffar-Bandjee, M.C.; Benhamou, L.S.; Tiran-Rajaofera, I.; et al. Severe forms of chikungunya virus infection in a pediatric intensive care unit on Reunion Island. Med. Trop. 2012, 72 Spec No, 88–93. [Google Scholar]
- Das, T.; Jaffar-Bandjee, M.C.; Hoarau, J.J.; Krejbich Trotot, P.; Denizot, M.; Lee-Pat-Yuen, G.; Sahoo, R.; Guiraud, P.; Ramful, D.; Robin, S.; et al. Chikungunya fever: CNS infection and pathologies of a re-emerging arbovirus. Prog. Neurobiol. 2010, 91, 121–129. [Google Scholar] [CrossRef]
- Zanluca, C.; Dos Santos, C.N.D. Zika virus—An overview. Microbes Infect. 2016, 18, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Zanluca, C.; Melo, V.C.A.; de Mosimann, A.L.P.; Santos, G.I.V.D.; Santos, C.N.D.D.; Luz, K. First report of autochthonous transmission of Zika virus in Brazil. Mem. Inst. Oswaldo Cruz 2015, 110, 569–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamel, R.; Liégeois, F.; Wichit, S.; Pompon, J.; Diop, F.; Talignani, L.; Thomas, F.; Desprès, P.; Yssel, H.; Missé, D. Zika virus: Epidemiology, clinical features and host-virus interactions. Microbes Infect. 2016, 18, 441–449. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [PubMed]
- Oliveira, D.B.L.; Almeida, F.J.; Durigon, E.L.; Mendes, É.A.; Braconi, C.T.; Marchetti, I.; Andreata-Santos, R.; Cunha, M.P.; Alves, R.P.S.; Pereira, L.R.; et al. Prolonged Shedding of Zika Virus Associated with Congenital Infection. N. Engl. J. Med. 2016, 375, 1202–1204. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, N.D.; Kilbourn, A.M.; Karesh, W.B.; Rahman, H.A.; Bosi, E.J.; Cropp, B.C.; Andau, M.; Spielman, A.; Gubler, D.J. Sylvatic transmission of arboviruses among Bornean orangutans. Am. J. Trop. Med. Hyg. 2001, 64, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Oster, A.M.; Russell, K.; Stryker, J.E.; Friedman, A.; Kachur, R.E.; Petersen, E.E.; Jamieson, D.J.; Cohn, A.C.; Brooks, J.T. Update: Interim Guidance for Prevention of Sexual Transmission of Zika Virus--United States, 2016. MMWR Morb. Mortal Wkly. Rep. 2016, 65, 323–325. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, A.M.; Sapiano, M.R.P.; Basavaraju, S.V.; Kuehnert, M.J.; Rivera-Garcia, B. Survey of Blood Collection Centers and Implementation of Guidance for Prevention of Transfusion-Transmitted Zika Virus Infection--Puerto Rico, 2016. MMWR Morb. Mortal Wkly. Rep. 2016, 65, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Briant, L.; Desprès, P.; Choumet, V.; Missé, D. Role of skin immune cells on the host susceptibility to mosquito-borne viruses. Virology 2014, 464–465, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host inflammatory response to mosquito bites enhances the severity of arbovirus infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef] [PubMed]
- Abere, B.; Wikan, N.; Ubol, S.; Auewarakul, P.; Paemanee, A.; Kittisenachai, S.; Roytrakul, S.; Smith, D.R. Proteomic analysis of chikungunya virus infected microgial cells. PLoS ONE 2012, 7, e34800. [Google Scholar] [CrossRef] [PubMed]
- Thio, C.L.-P.; Yusof, R.; Abdul-Rahman, P.S.A.; Karsani, S.A. Differential proteome analysis of chikungunya virus infection on host cells. PLoS ONE 2013, 8, e61444. [Google Scholar] [CrossRef] [PubMed]
- Wintachai, P.; Wikan, N.; Kuadkitkan, A.; Jaimipuk, T.; Ubol, S.; Pulmanausahakul, R.; Auewarakul, P.; Kasinrerk, W.; Weng, W.-Y.; Panyasrivanit, M.; et al. Identification of prohibitin as a Chikungunya virus receptor protein. J. Med. Virol. 2012, 84, 1757–1770. [Google Scholar] [CrossRef]
- Issac, T.H.K.; Tan, E.L.; Chu, J.J.H. Proteomic profiling of chikungunya virus-infected human muscle cells: Reveal the role of cytoskeleton network in CHIKV replication. J. Proteomics 2014, 108, 445–464. [Google Scholar] [CrossRef]
- Ekchariyawat, P.; Hamel, R.; Bernard, E.; Wichit, S.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; Choumet, V.; Yssel, H.; Desprès, P.; et al. Inflammasome signaling pathways exert antiviral effect against Chikungunya virus in human dermal fibroblasts. Infect. Genet. Evol. 2015, 32, 401–408. [Google Scholar] [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of zika virus infection in human skin cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [PubMed]
- Goujon, C.; Rivière, L.; Jarrosson-Wuilleme, L.; Bernaud, J.; Rigal, D.; Darlix, J.-L.; Cimarelli, A. SIVSM/HIV-2 Vpx proteins promote retroviral escape from a proteasome-dependent restriction pathway present in human dendritic cells. Retrovirology 2007, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Goujon, C.; Arfi, V.; Pertel, T.; Luban, J.; Lienard, J.; Rigal, D.; Darlix, J.-L.; Cimarelli, A. Characterization of simian immunodeficiency virus SIVSM/human immunodeficiency virus type 2 Vpx function in human myeloid cells. J. Virol. 2008, 82, 12335–12345. [Google Scholar] [CrossRef] [PubMed]
- Laguette, N.; Sobhian, B.; Casartelli, N.; Ringeard, M.; Chable-Bessia, C.; Ségéral, E.; Yatim, A.; Emiliani, S.; Schwartz, O.; Benkirane, M. SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 2011, 474, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, D.C.; Ennis-Adeniran, V.; Hedden, J.J.; Groom, H.C.T.; Rice, G.I.; Christodoulou, E.; Walker, P.A.; Kelly, G.; Haire, L.F.; Yap, M.W.; et al. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature 2011, 480, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Powell, R.D.; Holland, P.J.; Hollis, T.; Perrino, F.W. Aicardi-Goutieres syndrome gene and HIV-1 restriction factor SAMHD1 is a dGTP-regulated deoxynucleotide triphosphohydrolase. J. Biol. Chem. 2011, 286, 43596–43600. [Google Scholar] [CrossRef]
- Sommer, A.F.R.; Rivière, L.; Qu, B.; Schott, K.; Riess, M.; Ni, Y.; Shepard, C.; Schnellbächer, E.; Finkernagel, M.; Himmelsbach, K.; et al. Restrictive influence of SAMHD1 on Hepatitis B Virus life cycle. Sci. Rep. 2016, 6, 26616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhu, M.; Pan, X.; Zhu, Y.; Yan, H.; Jiang, T.; Shen, Y.; Dong, X.; Zheng, N.; Lu, J.; et al. Inhibition of Hepatitis B virus replication by SAMHD1. Biochem. Biophys. Res. Commun. 2014, 450, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Sourisseau, M.; Schilte, C.; Casartelli, N.; Trouillet, C.; Guivel-Benhassine, F.; Rudnicka, D.; Sol-Foulon, N.; Le Roux, K.; Prevost, M.-C.; Fsihi, H.; et al. Characterization of reemerging chikungunya virus. PLoS Pathog. 2007, 3, e89. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.J.; Bhasker, C.R.; Merrick, W.C.; Sen, G.C. Viral stress-inducible protein p56 inhibits translation by blocking the interaction of eIF3 with the ternary complex eIF2.GTP.Met-tRNAi. J. Biol. Chem. 2003, 278, 39477–39482. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Lassnig, C.; Eberle, C.-A.; Górna, M.W.; Baumann, C.L.; Burkard, T.R.; Bürckstümmer, T.; Stefanovic, A.; Krieger, S.; Bennett, K.L.; et al. IFIT1 is an antiviral protein that recognizes 5′-triphosphate RNA. Nat. Immunol. 2011, 12, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Schmeisser, H.; Mejido, J.; Balinsky, C.A.; Morrow, A.N.; Clark, C.R.; Zhao, T.; Zoon, K.C. Identification of alpha interferon-induced genes associated with antiviral activity in Daudi cells and characterization of IFIT3 as a novel antiviral gene. J. Virol. 2010, 84, 10671–10680. [Google Scholar] [CrossRef] [PubMed]
- Terenzi, F.; Saikia, P.; Sen, G.C. Interferon-inducible protein, P56, inhibits HPV DNA replication by binding to the viral protein E1. EMBO J. 2008, 27, 3311–3321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikia, P.; Fensterl, V.; Sen, G.C. The inhibitory action of P56 on select functions of E1 mediates interferon’s effect on human papillomavirus DNA replication. J. Virol. 2010, 84, 13036–13039. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, M.A.G.; Ribaudo, M.; Guo, J.-T.; Barik, S. Identification of Interferon-Stimulated Gene Proteins That Inhibit Human Parainfluenza Virus Type 3. J. Virol. 2016, 90, 11145–11156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynaud, J.M.; Kim, D.Y.; Atasheva, S.; Rasalouskaya, A.; White, J.P.; Diamond, M.S.; Weaver, S.C.; Frolova, E.I.; Frolov, I. IFIT1 Differentially Interferes with Translation and Replication of Alphavirus Genomes and Promotes Induction of Type I Interferon. PLoS Pathog. 2015, 11, e1004863. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Koonin, E.V. The HD domain defines a new superfamily of metal-dependent phosphohydrolases. Trends Biochem. Sci. 1998, 23, 469–472. [Google Scholar] [CrossRef]
- Hrecka, K.; Hao, C.; Gierszewska, M.; Swanson, S.K.; Kesik-Brodacka, M.; Srivastava, S.; Florens, L.; Washburn, M.P.; Skowronski, J. Vpx relieves inhibition of HIV-1 infection of macrophages mediated by the SAMHD1 protein. Nature 2011, 474, 658–661. [Google Scholar] [CrossRef] [Green Version]
- Berger, A.; Sommer, A.F.R.; Zwarg, J.; Hamdorf, M.; Welzel, K.; Esly, N.; Panitz, S.; Reuter, A.; Ramos, I.; Jatiani, A.; et al. SAMHD1-deficient CD14+ cells from individuals with Aicardi-Goutières syndrome are highly susceptible to HIV-1 infection. PLoS Pathog. 2011, 7, e1002425. [Google Scholar] [CrossRef]
- Lahouassa, H.; Daddacha, W.; Hofmann, H.; Ayinde, D.; Logue, E.C.; Dragin, L.; Bloch, N.; Maudet, C.; Bertrand, M.; Gramberg, T.; et al. SAMHD1 restricts the replication of human immunodeficiency virus type 1 by depleting the intracellular pool of deoxynucleoside triphosphates. Nat. Immunol. 2012, 13, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, J.; Choi, J.; Oh, C.; Kim, S.; Seo, M.; Kim, S.-Y.; Seo, D.; Kim, J.; White, T.E.; Brandariz-Nuñez, A.; et al. The ribonuclease activity of SAMHD1 is required for HIV-1 restriction. Nat. Med. 2014, 20, 936–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Ryoo, J.; Oh, C.; Hwang, S.; Ahn, K. SAMHD1 specifically restricts retroviruses through its RNase activity. Retrovirology 2015, 12, 46. [Google Scholar] [CrossRef] [PubMed]
- White, T.E.; Brandariz-Nuñez, A.; Valle-Casuso, J.C.; Amie, S.; Nguyen, L.; Kim, B.; Brojatsch, J.; Diaz-Griffero, F. Contribution of SAM and HD domains to retroviral restriction mediated by human SAMHD1. Virology 2013, 436, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Hollenbaugh, J.A.; Gee, P.; Baker, J.; Daly, M.B.; Amie, S.M.; Tate, J.; Kasai, N.; Kanemura, Y.; Kim, D.-H.; Ward, B.M.; et al. Host factor SAMHD1 restricts DNA viruses in non-dividing myeloid cells. PLoS Pathog. 2013, 9, e1003481. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.T.; White, T.E.; Brandariz-Núñez, A.; Diaz-Griffero, F.; Weitzman, M.D. SAMHD1 restricts herpes simplex virus 1 in macrophages by limiting DNA replication. J. Virol. 2013, 87, 12949–12956. [Google Scholar] [CrossRef]
- Zhao, K.; Du, J.; Han, X.; Goodier, J.L.; Li, P.; Zhou, X.; Wei, W.; Evans, S.L.; Li, L.; Zhang, W.; et al. Modulation of LINE-1 and Alu/SVA retrotransposition by Aicardi-Goutières syndrome-related SAMHD1. Cell Rep. 2013, 4, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Bonifati, S.; Qin, Z.; St Gelais, C.; Kodigepalli, K.M.; Barrett, B.S.; Kim, S.H.; Antonucci, J.M.; Ladner, K.J.; Buzovetsky, O.; et al. SAMHD1 suppresses innate immune responses to viral infections and inflammatory stimuli by inhibiting the NF-κB and interferon pathways. Proc Natl Acad Sci USA 2018, 115, E3798–E3807. [Google Scholar] [CrossRef] [PubMed]
- Descours, B.; Cribier, A.; Chable-Bessia, C.; Ayinde, D.; Rice, G.; Crow, Y.; Yatim, A.; Schwartz, O.; Laguette, N.; Benkirane, M. SAMHD1 restricts HIV-1 reverse transcription in quiescent CD4+ T-cells. Retrovirology 2012, 9, 87. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized P.P.B.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Reimand, J.; Arak, T.; Vilo, J. Profiler—A web server for functional interpretation of gene lists (2011 update). Nucleic Acids Res. 2011, 39, W307–W315. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Cribier, A.; Descours, B.; Valadão, A.L.C.; Laguette, N.; Benkirane, M. Phosphorylation of SAMHD1 by cyclin A2/CDK1 regulates its restriction activity toward HIV-1. Cell Rep. 2013, 3, 1036–1043. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteoform Name | Gene Name | Uniprot ID | Average log2 Ratio |
|---|---|---|---|
| Chikungunya virus infected cells proteins | |||
| Interferon-induced GTP-binding protein Mx1 | MX1 * | P20591 | 5.043 |
| Interferon-induced protein with tetratricopeptide repeats 3 | IFIT3 * | O14879 | 3.591 |
| Interferon-induced protein with tetratricopeptide repeats 1 | IFIT1 * | P09914 | 3.449 |
| Interferon-induced protein with tetratricopeptide repeats 2 | IFIT2 | P09913 | 2.954 |
| Ubiquitin-like protein ISG15 | ISG15 * | P05161 | 2.580 |
| Probable ATP-dependent RNA helicase DDX58 | DDX58 * | O95786 | 2.444 |
| Signal transducer and activator of transcription 1-alpha/beta | STAT1 * | P42224 | 1.659 |
| 2′-5′-oligoadenylate synthase 3 | OAS3 * | Q9Y6K5 | 1.626 |
| Deoxynucleoside triphosphate triphosphohydrolase SAMHD1 | SAMHD1 * | Q9Y3Z3 | 1.208 |
| HLA class I histocompatibility antigen, B-45 alpha chain | HLA-B | P30483 | 1.119 |
| Cytosol aminopeptidase | LAP3 | P28838 | 1.049 |
| Interferon-induced 35 kDa protein | IFI35 | P80217 | 1.015 |
| E3 ubiquitin-protein ligase RNF213 | RNF213 | Q63HN8 | 0.895 |
| Gamma-interferon-inducible protein 16 | IFI16 | Q16666 | 0.878 |
| Double-stranded RNA-activated protein kinase | EIF2AK2 * | P19525 | 0.838 |
| HLA class I histocompatibility antigen, A-69 alpha chain | HLA-A | P10316 | 0.752 |
| Zika virus infected cells proteins | |||
| Interferon-induced GTP-binding protein Mx1 | MX1 * | P20591 | 5.622 |
| Interferon-induced protein with tetratricopeptide repeats 1 | IFIT1 * | P09914 | 3.362 |
| Interferon-induced protein with tetratricopeptide repeats 3 | IFIT3 * | O14879 | 3.076 |
| Ubiquitin-like protein ISG15 | ISG15 * | P05161 | 2.430 |
| 2′-5′-oligoadenylate synthase 3 | OAS3 * | Q9Y6K5 | 2.190 |
| Poly [ADP-ribose] polymerase 9 | PARP9 | Q8IXQ6-2 | 2.118 |
| Phospholipid scramblase 1 | PLSCR1 | O15162 | 2.085 |
| Probable ATP-dependent RNA helicase DDX58 | DDX58 * | O95786 | 1.974 |
| Deoxynucleoside triphosphate triphosphohydrolase SAMHD1 | SAMHD1 * | Q9Y3Z3 | 1.739 |
| Signal transducer and activator of transcription 1-alpha/beta | STAT1 * | P42224 | 1.440 |
| Synaptopodin-2 | SYNPO2 | Q9UMS6 | 1.415 |
| E3 ubiquitin-protein ligase DTX3L | DTX3L | Q8TDB6 | 1.397 |
| 2′-5′-oligoadenylate synthase 1 | OAS1 | P00973 | 1.135 |
| E3 ubiquitin-protein ligase TRIM21 | TRIM21 | P19474 | 1.085 |
| Double-stranded RNA-activated protein kinase | EIF2AK2 * | P19525 | 1.071 |
| Interferon-induced GTP-binding protein Mx2 | MX2 | P20592 | 0.993 |
| Chikungunya Virus | ||
| Biological Processes | GO Term | Corrected p-Value |
| GO ID | ||
| GO:0060337 | Type I interferon signaling pathway | 9.94 × 10−21 |
| GO:0051607 | Defense response to virus * | 3.53 × 10−16 |
| GO:0048525 | Negative regulation of virus infection process * | 1.37 × 10−10 |
| GO:0045071 | Negative regulation of viral genome replication | 4.25 × 10−10 |
| GO:0031348 | Negative regulation of defense response | 0.0000326 |
| GO:0035455 | Response to interferon-alpha * | 0.000153 |
| GO:0045088 | Regulation of innate immune response * | 0.000212 |
| GO:0060333 | Interferon-gamma-mediated signaling pathway | 0.000249 |
| GO:0002698 | Negative regulation of immune effector process | 0.000407 |
| GO:0043330 | Response to exogenous dsRNA * | 0.00281 |
| GO:0050688 | Regulation of defense response to virus | 0.00602 |
| KEGG | Pathway Name | Corrected p-Value |
| Pathway ID | ||
| KEGG:05168 | Herpes simplex infection | 4.12 × 10−8 |
| KEGG:05160 | Hepatitis C * | 0.0000275 |
| KEGG:05162 | Measles * | 0.0000286 |
| KEGG:05164 | Influenza A | 0.000101 |
| KEGG:05169 | Epstein-Barr virus infection | 0.00553 |
| Reactome | Pathway Name | Corrected p-Value |
| Pathway ID | ||
| REAC:913531 | Interferon Signaling * | 5.41 × 10−19 |
| Zika Virus | ||
| Biological Processes | GO Term | Corrected p-Value |
| GO ID | ||
| GO:0051607 | Defense response to virus * | 5.74 × 10−16 |
| GO:0071357 | Cellular response to type I interferon | 2.77 × 10−15 |
| GO:0048525 | Negative regulation of viral process * | 3.85 × 10−15 |
| GO:1903901 | Negative regulation of viral life cycle | 7.82 × 10−13 |
| GO:0035455 | Response to interferon-alpha * | 0.000000633 |
| GO:0034341 | Response to interferon-gamma | 0.0000822 |
| GO:0043330 | Response to exogenous dsRNA * | 0.00456 |
| GO:0045088 | Regulation of innate immune response* | 0.00899 |
| KEGG | Pathway Name | Corrected p-Value |
| Pathway ID | ||
| KEGG:05160 | Hepatitis C * | 0.0104 |
| KEGG:05162 | Measles * | 0.0108 |
| Reactome | Pathway Name | Corrected p-Value |
| Pathway ID | ||
| REAC:913531 | Interferon Signaling * | 1.22 × 10−15 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wichit, S.; Hamel, R.; Zanzoni, A.; Diop, F.; Cribier, A.; Talignani, L.; Diack, A.; Ferraris, P.; Liegeois, F.; Urbach, S.; et al. SAMHD1 Enhances Chikungunya and Zika Virus Replication in Human Skin Fibroblasts. Int. J. Mol. Sci. 2019, 20, 1695. https://doi.org/10.3390/ijms20071695
Wichit S, Hamel R, Zanzoni A, Diop F, Cribier A, Talignani L, Diack A, Ferraris P, Liegeois F, Urbach S, et al. SAMHD1 Enhances Chikungunya and Zika Virus Replication in Human Skin Fibroblasts. International Journal of Molecular Sciences. 2019; 20(7):1695. https://doi.org/10.3390/ijms20071695
Chicago/Turabian StyleWichit, Sineewanlaya, Rodolphe Hamel, Andreas Zanzoni, Fodé Diop, Alexandra Cribier, Loïc Talignani, Abibatou Diack, Pauline Ferraris, Florian Liegeois, Serge Urbach, and et al. 2019. "SAMHD1 Enhances Chikungunya and Zika Virus Replication in Human Skin Fibroblasts" International Journal of Molecular Sciences 20, no. 7: 1695. https://doi.org/10.3390/ijms20071695