Telomeres and Telomerase in Cutaneous Squamous Cell Carcinoma

 , ,
, ,
Abstract
:1. Introduction
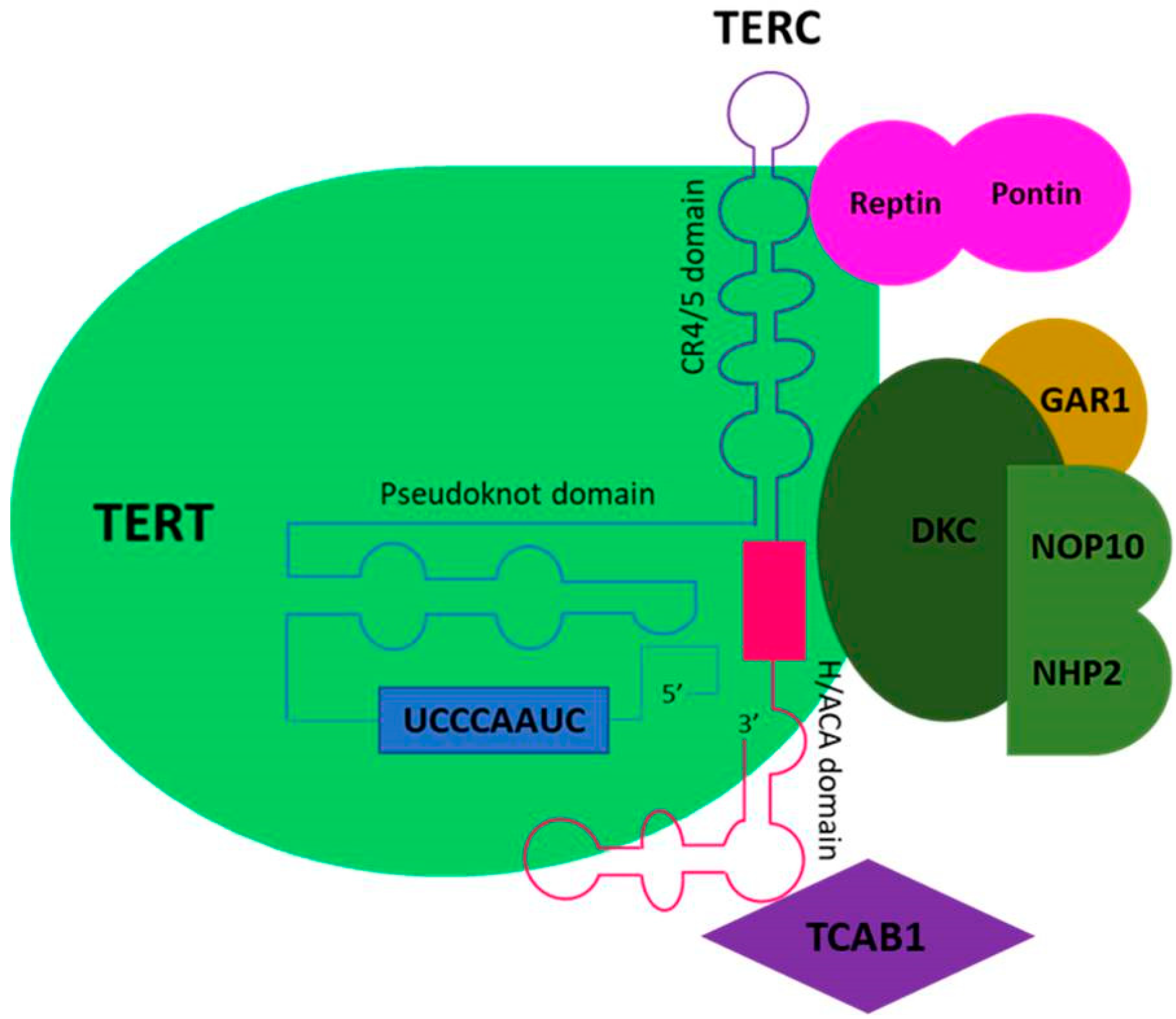
2. Physiologic Role of Telomeres
3. Telomere Biology in Cancerogenesis
4. Telomere and Telomerase Activity in Cutaneous Squamous Cell Carcinoma
4.1. Telomere Length
4.2. Telomerase Activity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Motley, R.; Kersey, P.; Lawrence, C.; British Association of Dermatologists; British Association of Plastic Surgeons; Royal College of Radiologists, Faculty of Clinical Oncology. Multiprofessional guidelines for the management of the patient with primary cutaneous squamous cell carcinoma. Br. J. Dermatol. 2002, 146, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, D.S.; Stern, R.S. Nonmelanoma cancers of the skin. N. Engl. J. Med. 1992, 327, 1649–1662. [Google Scholar] [CrossRef]
- Miller, D.L.; Weinstock, M.A. Nonmelanoma skin cancer in the United States: Incidence. J. Am. Acad. Dermatol. 1994, 30 Pt 1, 774–778. [Google Scholar] [CrossRef]
- Gloster, H.M., Jr.; Brodland, D.G. The epidemiology of skin cancer. Dermatol. Surg. 1996, 22, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Holme, S.A.; Malinovszky, K.; Roberts, D.L. Changing trends in non-melanoma skin cancer in South Wales, 1988–1998. Br. J. Dermatol. 2000, 143, 1224–1229. [Google Scholar] [CrossRef] [PubMed]
- Revenga Arranz, F.; Paricio Rubio, J.F.; Mar Vazquez Salvado, M.; del Villar Sordo, V. Descriptive epidemiology of basal cell carcinoma and cutaneous squamous cell carcinoma in Soria (north-eastern Spain) 1998–2000: A hospital-based survey. J. Eur. Acad. Dermatol. Venereol. 2004, 18, 137–141. [Google Scholar] [CrossRef] [PubMed]
- De Vries, E.; Trakatelli, M.; Kalabalikis, D.; Ferrandiz, L.; Ruiz-de-Casas, A.; Moreno-Ramirez, D.; Sotiriadis, D.; Ioannides, D.; Aquilina, S.; Apap, C.; et al. Known and potential new risk factors for skin cancer in European populations: A multicentre case-control study. Br. J. Dermatol. 2012, 167 (Suppl. 2), 1–13. [Google Scholar] [CrossRef]
- Peris, K.; Alaibac, M.; Argenziano, G.; Di Stefani, A.; Fargnoli, M.C.; Frascione, P.; Gualdi, G.; Longo, C.; Moscarella, E.; Naldi, L.; et al. Cutaneous squamous cell carcinoma. Italian Guidelines by SIDeMaST adapted to and updating EADO/EDF/EORTC guidelines. G. Ital. Dermatol. Venereol. 2018, 153, 747–762. [Google Scholar] [CrossRef]
- Fargnoli, M.C.; Piccioni, A.; Neri, L.; Tambone, S.; Pellegrini, C.; Peris, K. Long-term efficacy and safety of daylight photodynamic therapy with methyl amninolevulinate for actinic keratosis of the face and scalp. Eur. J. Dermatol. 2017, 27, 89–91. [Google Scholar] [PubMed]
- Werner, R.N.; Sammain, A.; Erdmann, R.; Hartmann, V.; Stockfleth, E.; Nast, A. The natural history of actinic keratosis: A systematic review. Br. J. Dermatol. 2013, 169, 502–518. [Google Scholar] [CrossRef]
- Alam, M.; Ratner, D. Cutaneous squamous-cell carcinoma. N. Engl. J. Med. 2001, 344, 975–983. [Google Scholar] [CrossRef]
- Warner, C.L.; Cockerell, C.J. The new seventh edition American Joint Committee on Cancer staging of cutaneous non-melanoma skin cancer: A critical review. Am. J. Clin. Dermatol. 2011, 12, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Orlandi, A.; Costanza, G.; Di Stefani, A.; Piccioni, A.; Di Cesare, A.; Chiricozzi, A.; Ferlosio, A.; Peris, K.; Fargnoli, M.C. Expression of IL-23/Th17-related cytokines in basal cell carcinoma and in the response to medical treatments. PLoS ONE 2017, 12, e0183415. [Google Scholar] [CrossRef] [PubMed]
- Boukamp, P. Non-melanoma skin cancer: What drives tumor development and progression? Carcinogenesis 2005, 26, 1657–1667. [Google Scholar] [CrossRef]
- Forbes, S.A.; Beare, D.; Boutselakis, H.; Bamford, S.; Bindal, N.; Tate, J.; Cole, C.G.; Ward, S.; Dawson, E.; Ponting, L.; et al. COSMIC: Somatic cancer genetics at high-resolution. Nucleic Acids Res. 2017, 45, D777–D783. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Maturo, M.G.; Martorelli, C.; Suppa, M.; Antonini, A.; Kostaki, D.; Verna, L.; Landi, M.T.; Peris, K.; Fargnoli, M.C. Characterization of melanoma susceptibility genes in high-risk patients from Central Italy. Melanoma Res. 2017, 27, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Lencina, J.J.; Rachakonda, S.; García-Casado, Z.; Srinivas, N.; Skorokhod, A.; Requena, C.; Soriano, V.; Kumar, R.; Nagore, E. TERT promoter mutation subtypes and survival in stage I and II melanoma patients. Int. J. Cancer 2019, 144, 1027–1036. [Google Scholar] [CrossRef]
- Taylor, R.S.; Ramirez, R.D.; Ogoshi, M.; Chaffins, M.; Piatyszek, M.A.; Shay, J.W. Detection of telomerase activity in malignant and nonmalignant skin conditions. J. Investig. Dermatol. 1996, 106, 759–765. [Google Scholar] [CrossRef]
- Parris, C.N.; Jezzard, S.; Silver, A.; MacKie, R.; McGregor, J.M.; Newbold, R.F. Telomerase activity in melanoma and non-melanoma skin cancer. Br. J. Cancer 1999, 79, 47–53. [Google Scholar] [CrossRef]
- Pellegrini, C.; Maturo, M.G.; Di Nardo, L.; Ciciarelli, V.; Gutierrez Garcia-Rodrigo, C.; Fargnoli, M.C. Understanding the Molecular Genetics of Basal Cell Carcinoma. Int. J. Mol. Sci. 2017, 18, 2485. [Google Scholar] [CrossRef]
- Gunes, C.; Rudolph, K.L. The role of telomeres in stem cells and cancer. Cell 2013, 152, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandakumar, J.; Cech, T.R. Finding the end: Recruitment of telomerase to telomeres. Nat. Rev. Mol. Cell Biol. 2013, 14, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Rachakonda, S.; Kong, H.; Srinivas, N.; Garcia-Casado, Z.; Requena, C.; Fallah, M.; Heidenreich, B.; Planelles, D.; Traves, V.; Schadendorf, D.; et al. Telomere length, telomerase reverse transcriptase promoter mutations, and melanoma risk. Genes Chromosomes Cancer 2018, 57, 564–572. [Google Scholar] [CrossRef]
- Szostak, J.W.; Blackburn, E.H. Cloning yeast telomeres on linear plasmid vectors. Cell 1982, 29, 245–255. [Google Scholar] [CrossRef]
- Blackburn, E.H. Structure and function of telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Levy, M.Z.; Allsopp, R.C.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere end-replication problem and cell aging. J. Mol. Biol. 1992, 225, 951–960. [Google Scholar] [CrossRef]
- Harley, C.B. Telomere loss: Mitotic clock or genetic time bomb? Mutat. Res. 1991, 256, 271–282. [Google Scholar] [CrossRef]
- Blackburn, E.H. Switching and signaling at the telomere. Cell 2001, 106, 661–673. [Google Scholar] [CrossRef]
- Sharpless, N.E.; DePinho, R.A. Telomeres, stem cells, senescence, and cancer. J. Clin. Investig. 2004, 113, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Liu, D.; Songyang, Z. The telosome/shelterin complex and its functions. Genome Biol. 2008, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- De Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Martinez, P.; Blasco, M.A. Telomeric and extra-telomeric roles for telomerase and the telomere-binding proteins. Nat. Rev. Cancer 2011, 11, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Cesare, A.J.; Reddel, R.R. Alternative lengthening of telomeres: Models, mechanisms and implications. Nat. Rev. Genet. 2010, 11, 319–330. [Google Scholar] [CrossRef]
- Morin, G.B. The human telomere terminal transferase enzyme is a ribonucleoprotein that synthesizes TTAGGG repeats. Cell 1989, 59, 521–529. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 1985, 43 Pt 1, 405–413. [Google Scholar] [CrossRef]
- Chen, Z.; Smith, K.J.; Skelton, H.G., 3rd; Barrett, T.L.; Greenway, H.T., Jr.; Lo, S.C. Telomerase activity in Kaposi’s sarcoma, squamous cell carcinoma, and basal cell carcinoma. Exp. Biol. Med. 2001, 226, 753–757. [Google Scholar] [CrossRef]
- Pogacic, V.; Dragon, F.; Filipowicz, W. Human H/ACA small nucleolar RNPs and telomerase share evolutionarily conserved proteins NHP2 and NOP10. Mol. Cell. Biol. 2000, 20, 9028–9040. [Google Scholar] [CrossRef]
- Daniel, M.; Peek, G.W.; Tollefsbol, T.O. Regulation of the human catalytic subunit of telomerase (hTERT). Gene 2012, 498, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Akincilar, S.C.; Unal, B.; Tergaonkar, V. Reactivation of telomerase in cancer. Cell. Mol. Life Sci. 2016, 73, 1659–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmeyer, K.; Raggioli, A.; Rudloff, S.; Anton, R.; Hierholzer, A.; Del Valle, I.; Hein, K.; Vogt, R.; Kemler, R. Wnt/beta-catenin signaling regulates telomerase in stem cells and cancer cells. Science 2012, 336, 1549–1554. [Google Scholar] [CrossRef]
- Wong, C.W.; Hou, P.S.; Tseng, S.F.; Chien, C.L.; Wu, K.J.; Chen, H.F.; Ho, H.N.; Kyo, S.; Teng, S.C. Kruppel-like transcription factor 4 contributes to maintenance of telomerase activity in stem cells. Stem Cells 2010, 28, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Aviv, A.; Anderson, J.J.; Shay, J.W. Mutations, Cancer and the Telomere Length Paradox. Trends Cancer 2017, 3, 253–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Zhou, Z.; Wei, S.; Liu, Z.; Pooley, K.A.; Dunning, A.M.; Svenson, U.; Roos, G.; Hosgood, H.D., 3rd; Shen, M.; et al. Shortened telomere length is associated with increased risk of cancer: A meta-analysis. PLoS ONE 2011, 6, e20466. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Wright, W.E. Role of telomeres and telomerase in cancer. Semin. Cancer Biol. 2011, 21, 349–353. [Google Scholar] [CrossRef] [Green Version]
- Wentzensen, I.M.; Mirabello, L.; Pfeiffer, R.M.; Savage, S.A. The association of telomere length and cancer: A meta-analysis. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Gramatges, M.M.; Telli, M.L.; Balise, R.; Ford, J.M. Longer relative telomere length in blood from women with sporadic and familial breast cancer compared with healthy controls. Cancer Epidemiol. Biomark. Prev. 2010, 19, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Svenson, U.; Nordfjall, K.; Stegmayr, B.; Manjer, J.; Nilsson, P.; Tavelin, B.; Henriksson, R.; Lenner, P.; Roos, G. Breast cancer survival is associated with telomere length in peripheral blood cells. Cancer Res. 2008, 68, 3618–3623. [Google Scholar] [CrossRef]
- Zhang, C.; Doherty, J.A.; Burgess, S.; Hung, R.J.; Lindstrom, S.; Kraft, P.; Gong, J.; Amos, C.I.; Sellers, T.A.; Monteiro, A.N.; et al. Genetic determinants of telomere length and risk of common cancers: A Mendelian randomization study. Hum. Mol. Genet. 2015, 24, 5356–5366. [Google Scholar] [CrossRef]
- Wright, W.E.; Piatyszek, M.A.; Rainey, W.E.; Byrd, W.; Shay, J.W. Telomerase activity in human germline and embryonic tissues and cells. Dev. Genet. 1996, 18, 173–179. [Google Scholar] [CrossRef]
- Collins, K.; Mitchell, J.R. Telomerase in the human organism. Oncogene 2002, 21, 564–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, N.D.; Baird, D.M. Telomere length maintenance in stem cell populations. Biochim. Biophys. Acta 2009, 1792, 324–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weise, J.M.; Gunes, C. Differential regulation of human and mouse telomerase reverse transcriptase (TERT) promoter activity during testis development. Mol. Reprod. Dev. 2009, 76, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Kyo, S.; Takakura, M.; Fujiwara, T.; Inoue, M. Understanding and exploiting hTERT promoter regulation for diagnosis and treatment of human cancers. Cancer Sci. 2008, 99, 1528–1538. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Q.; Li, K.; Chen, L.; Li, W.; Hou, M.; Liu, T.; Yang, J.; Lindvall, C.; Björkholm, M.; et al. Telomerase reverse transcriptase promotes epithelial-mesenchymal transition and stem cell-like traits in cancer cells. Oncogene 2013, 32, 4203–4213. [Google Scholar] [CrossRef] [PubMed]
- Lagunas, A.M.; Wu, J.; Crowe, D.L. Telomere DNA damage signaling regulates cancer stem cell evolution, epithelial mesenchymal transition, and metastasis. Oncotarget 2017, 8, 80139–80155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Badawy, A.; Ghoneim, N.I.; Nasr, M.A.; Elkhenany, H.; Ahmed, T.A.; Ahmed, S.M.; El-Badri, N. Telomerase reverse transcriptase coordinates with the epithelial-to-mesenchymal transition through a feedback loop to define properties of breast cancer stem cells. Biol. Open 2018, 7, bio034181. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Di Nardo, L.; Cipolloni, G.; Martorelli, C.; De Padova, M.; Antonini, A.; Maturo, M.G.; Del Regno, L.; Strafella, S.; Micantonio, T.; et al. Heterogeneity of BRAF, NRAS, and TERT Promoter Mutational Status in Multiple Melanomas and Association with MC1R Genotype: Findings from Molecular and Immunohistochemical Analysis. J. Mol. Diagn. 2018, 20, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Nagore, E.; Heidenreich, B.; Requena, C.; García-Casado, Z.; Martorell-Calatayud, A.; Pont-Sanjuan, V.; Jimenez-Sanchez, A.I.; Kumar, R. TERT promoter mutations associate with fast-growing melanoma. Pigment Cell Melanoma Res. 2016, 29, 236–238. [Google Scholar] [CrossRef]
- Pópulo, H.; Boaventura, P.; Vinagre, J.; Batista, R.; Mendes, A.; Caldas, R.; Pardal, J.; Azevedo, F.; Honavar, M.; Guimarães, I.; et al. TERT promoter mutations in skin cancer: The effects of sun exposure and X-irradiation. J. Investig. Dermatol. 2014, 134, 2251–2257. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, J.; Pinto, V.; Celestino, R.; Reis, M.; Populo, H.; Boaventura, P.; Melo, M.; Catarino, T.; Lima, J.; Lopes, J.M.; et al. Telomerase promoter mutations in cancer: An emerging molecular biomarker? Virchows Arch. 2014, 465, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Khan, R.; Gupta, N.; Seth, T.; Sharma, A.; Kalaivani, M.; Sharma, A. Identifying the biomarker potential of telomerase activity and shelterin complex molecule, telomeric repeat binding factor 2 (TERF2), in multiple myeloma. Leuk. Lymphoma 2018, 59, 1677–1689. [Google Scholar] [CrossRef] [PubMed]
- The Telomeres Mendelian Randomization Collaboration; Haycock, P.C.; Burgess, S.; Nounu, A.; Zheng, J.; Okoli, G.N.; Bowden, J.; Wade, K.H.; Timpson, N.J.; Evans, D.M.; et al. Association Between Telomere Length and Risk of Cancer and Non-Neoplastic Diseases: A Mendelian Randomization Study. JAMA Oncol. 2017, 3, 636–651. [Google Scholar] [CrossRef] [PubMed]
- Ribero, S.; Mangino, M.; Bataille, V. Skin phenotypes can offer some insight about the association between telomere length and cancer susceptibility. Med. Hypotheses 2016, 97, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Horn, T. Comments on quantitative real-time PCR for measurement of telomere length. Cancer Investig. 2008, 26, 867. [Google Scholar]
- Bataille, V.; Kato, B.S.; Falchi, M.; Gardner, J.; Kimura, M.; Lens, M.; Perks, U.; Valdes, A.M.; Bennett, D.C.; Aviv, A.; et al. Nevus size and number are associated with telomere length and represent potential markers of a decreased senescence in vivo. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1499–1502. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.J. Telomeres and cancer: From crisis to stability to crisis to stability. Cell 2012, 148, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Qureshi, A.A.; Prescott, J.; Guo, Q.; Ye, L.; Hunter, D.J.; De Vivo, I. A prospective study of telomere length and the risk of skin cancer. J. Investig. Dermatol. 2009, 129, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, L.J.; Middleton, P.G.; Rees, J.L. Changes in mean telomere length in basal cell carcinomas of the skin. Genes Chromosomes Cancer 1995, 12, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Griewank, K.G.; Murali, R.; Schilling, B.; Schimming, T.; Moller, I.; Moll, I.; Schwamborn, M.; Sucker, A.; Zimmer, L.; Schadendorf, D.; et al. TERT promoter mutations are frequent in cutaneous basal cell carcinoma and squamous cell carcinoma. PLoS ONE 2013, 8, e80354. [Google Scholar] [CrossRef]
- Toll, A.; Salgado, R.; Yebenes, M.; Martin-Ezquerra, G.; Gilaberte, M.; Baro, T.; Sole, F.; Alameda, F.; Espinet, B.; Pujol, R.M. MYC gene numerical aberrations in actinic keratosis and cutaneous squamous cell carcinoma. Br. J. Dermatol. 2009, 161, 1112–1118. [Google Scholar] [CrossRef]
- Desmaze, C.; Soria, J.C.; Freulet-Marriere, M.A.; Mathieu, N.; Sabatier, L. Telomere-driven genomic instability in cancer cells. Cancer Lett. 2003, 194, 173–182. [Google Scholar] [CrossRef]
- Perrem, K.; Lynch, A.; Conneely, M.; Wahlberg, H.; Murphy, G.; Leader, M.; Kay, E. The higher incidence of squamous cell carcinoma in renal transplant recipients is associated with increased telomere lengths. Hum. Pathol. 2007, 38, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Leufke, C.; Leykauf, J.; Krunic, D.; Jauch, A.; Holtgreve-Grez, H.; Bohm-Steuer, B.; Brocker, E.B.; Mauch, C.; Utikal, J.; Hartschuh, W.; et al. The telomere profile distinguishes two classes of genetically distinct cutaneous squamous cell carcinomas. Oncogene 2014, 33, 3506–3518. [Google Scholar] [CrossRef] [PubMed]
- Yamada-Hishida, H.; Nobeyama, Y.; Nakagawa, H. Correlation of telomere length to malignancy potential in non-melanoma skin cancers. Oncol. Lett. 2018, 15, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Caini, S.; Raimondi, S.; Johansson, H.; De Giorgi, V.; Zanna, I.; Palli, D.; Gandini, S. Telomere length and the risk of cutaneous melanoma and non-melanoma skin cancer: A review of the literature and meta-analysis. J. Dermatol. Sci. 2015, 80, 168–174. [Google Scholar] [CrossRef]
- Liang, G.; Qureshi, A.A.; Guo, Q.; De Vivo, I.; Han, J. No association between telomere length in peripheral blood leukocytes and the risk of nonmelanoma skin cancer. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1043–1045. [Google Scholar] [CrossRef]
- Anic, G.M.; Sondak, V.K.; Messina, J.L.; Fenske, N.A.; Zager, J.S.; Cherpelis, B.S.; Lee, J.H.; Fulp, W.J.; Epling-Burnette, P.K.; Park, J.Y.; et al. Telomere length and risk of melanoma, squamous cell carcinoma, and basal cell carcinoma. Cancer Epidemiol. 2013, 37, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Ueda, M.; Ouhtit, A.; Bito, T.; Nakazawa, K.; Lubbe, J.; Ichihashi, M.; Yamasaki, H.; Nakazawa, H. Evidence for UV-associated activation of telomerase in human skin. Cancer Res. 1997, 57, 370–374. [Google Scholar] [PubMed]
- Boldrini, L.; Loggini, B.; Gisfredi, S.; Zucconi, Y.; Di Quirico, D.; Biondi, R.; Cervadoro, G.; Barachini, P.; Basolo, F.; Pingitore, R.; et al. Evaluation of telomerase in non-melanoma skin cancer. Int. J. Mol. Med. 2003, 11, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Burnworth, B.; Arendt, S.; Muffler, S.; Steinkraus, V.; Brocker, E.B.; Birek, C.; Hartschuh, W.; Jauch, A.; Boukamp, P. The multi-step process of human skin carcinogenesis: A role for p53, cyclin D1, hTERT, p16, and TSP-1. Eur. J. Cell Biol. 2007, 86, 763–780. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.A.; Laughlin, T.S.; Rothberg, P.G. Mutations of the TERT promoter are common in basal cell carcinoma and squamous cell carcinoma. Mod. Pathol. 2014, 27, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.A.; Macedo, S.; Fernandes, M.; Pestana, A.; Pardal, J.; Batista, R.; Vinagre, J.; Sanches, A.; Baptista, A.; Lopes, J.M.; et al. TERT promoter mutations are associated with poor prognosis in cutaneous squamous cell carcinoma. J. Am. Acad. Dermatol. 2019, 80, 660–669.e6. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.A.; Kurtis, B.; Babayeva, S.; Zhuge, J.; Tantchou, I.; Cai, D.; Lafaro, R.J.; Fallon, J.T.; Zhong, M. Heterogeneity of TERT promoter mutations status in squamous cell carcinomas of different anatomical sites. Ann. Diagn. Pathol. 2015, 19, 146–148. [Google Scholar] [CrossRef]
- Nan, H.; Qureshi, A.A.; Prescott, J.; De Vivo, I.; Han, J. Genetic variants in telomere-maintaining genes and skin cancer risk. Hum. Genet. 2011, 129, 247–253. [Google Scholar] [CrossRef]



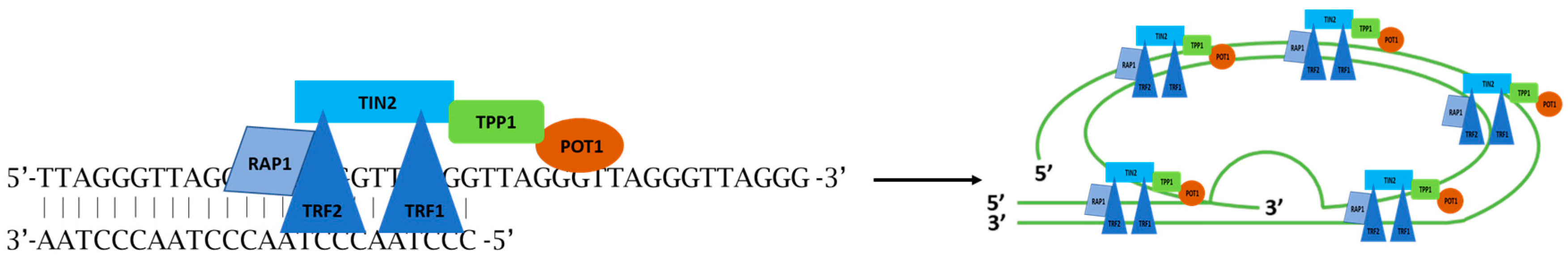{kind=link}
{kind=link}
| Study | Year | Molecular Alterations Investigated | Number of Cases | Main Findings |
|---|---|---|---|---|
| Taylor, R.S. et al. [18] | 1996 | Telomerase activity | 18 | Telomerase activity was detected in 84% of cSCC |
| Burnworth, B. et al. [83] | 2006 | TERT expression | 31 | Telomerase expression is not specifically increased during cSCC malignant progression |
| Perrem, K. et al. [75] | 2007 | Telomere length | 66 | Telomeres were longer in cSCC tumours of transplanted patients as compared to those of non-transplanted patients |
| Han, J. et al. [70] | 2009 | Telomere length | 285 | No association was observed between telomere length and cSCC risk |
| Liang, G. et al. [79] | 2011 | Telomere length | 241 | No significant association was detected between telomere length and risk of cSCC |
| Griewank, K.G. et al. [72] | 2013 | TERT promoter mutations | 34 | TERT promoter mutations were identified in 50% of cSCC cases |
| Scott, G.A. et al. [84] | 2013 | TERT promoter mutations | 26 | 50% of cSCC presented classic mutations of TERT promoter * |
| Anic, G.M. et al. [80] | 2013 | Telomere length | 136 | Long telomere length was inversely associated with cSCC risk |
| Leufke, C. et al. [76] | 2014 | Telomere profile | 32 | cSCC expressed two telomere phenotypes: (i) tissue-wide short to intermediate telomere length and (ii) tissue-wide heterogeneous longer telomere, suggesting two modes of initiation |
| Cheng, K.A. et al. [86] | 2015 | TERT promoter mutations | 84 | TERT promoter mutations were identified in 70% of cutaneous cSCC |
| Yamada-Hishida, H. et al. [77] | 2017 | Telomere length | 9 | Telomere length of cSCC was significantly shorter than those of BD and AK; telomere shortening is correlated with invasive progression |
| Campos, M.A. et al. [85] | 2019 | TERT promoter mutations | 184 | TERT promoter mutations are associated with poor prognosis in cSCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventura, A.; Pellegrini, C.; Cardelli, L.; Rocco, T.; Ciciarelli, V.; Peris, K.; Fargnoli, M.C. Telomeres and Telomerase in Cutaneous Squamous Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 1333. https://doi.org/10.3390/ijms20061333
Ventura A, Pellegrini C, Cardelli L, Rocco T, Ciciarelli V, Peris K, Fargnoli MC. Telomeres and Telomerase in Cutaneous Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2019; 20(6):1333. https://doi.org/10.3390/ijms20061333
Chicago/Turabian StyleVentura, Alessandra, Cristina Pellegrini, Ludovica Cardelli, Tea Rocco, Valeria Ciciarelli, Ketty Peris, and Maria Concetta Fargnoli. 2019. "Telomeres and Telomerase in Cutaneous Squamous Cell Carcinoma" International Journal of Molecular Sciences 20, no. 6: 1333. https://doi.org/10.3390/ijms20061333