Global Gene Expression of Seed Coat Tissues Reveals a Potential Mechanism of Regulating Seed Size Formation in Castor Bean

Abstract
:1. Introduction
2. Results
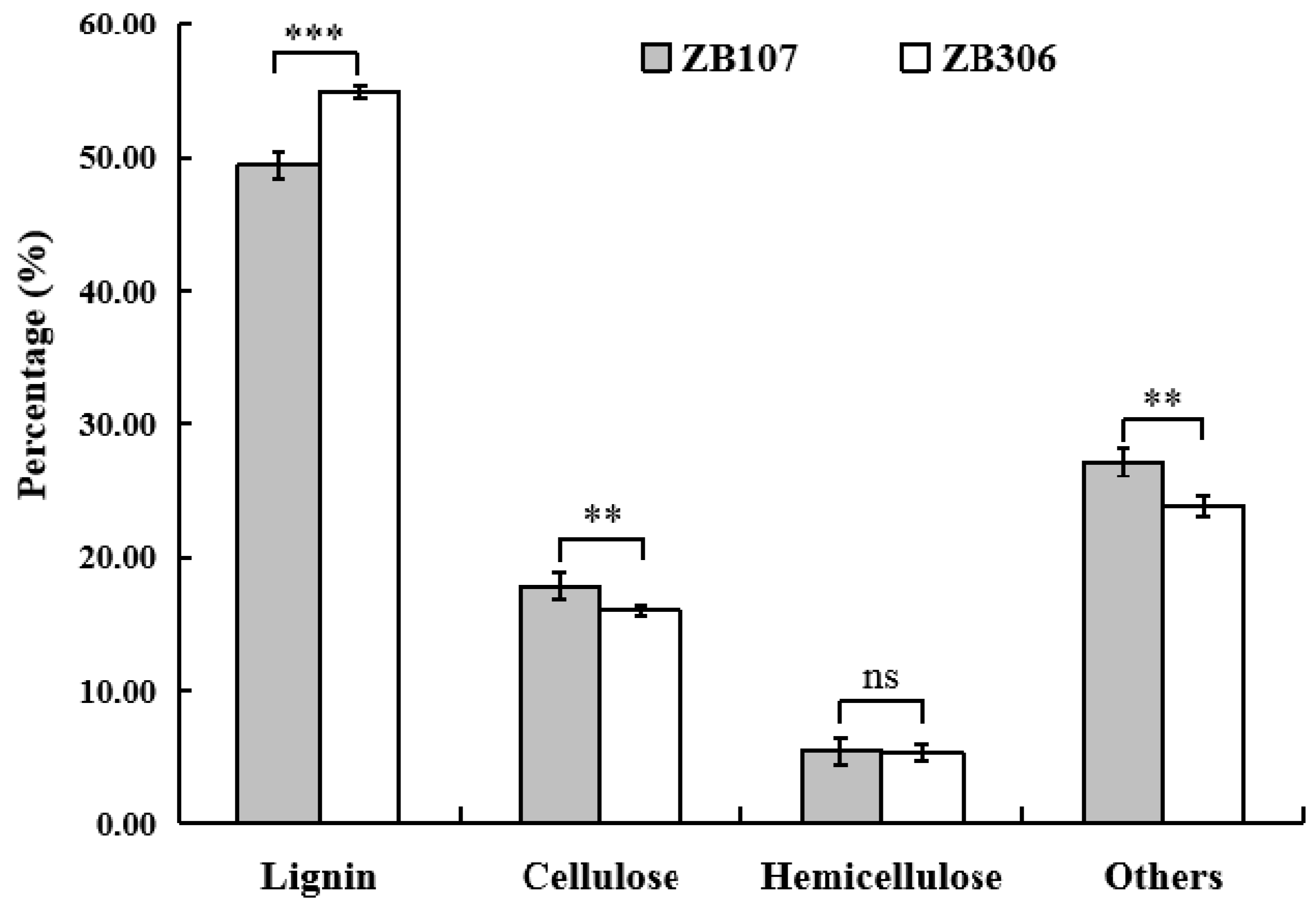
2.1. Morphological Analysis of Seed Development and Determination of Seed Coat Fiber Compositions
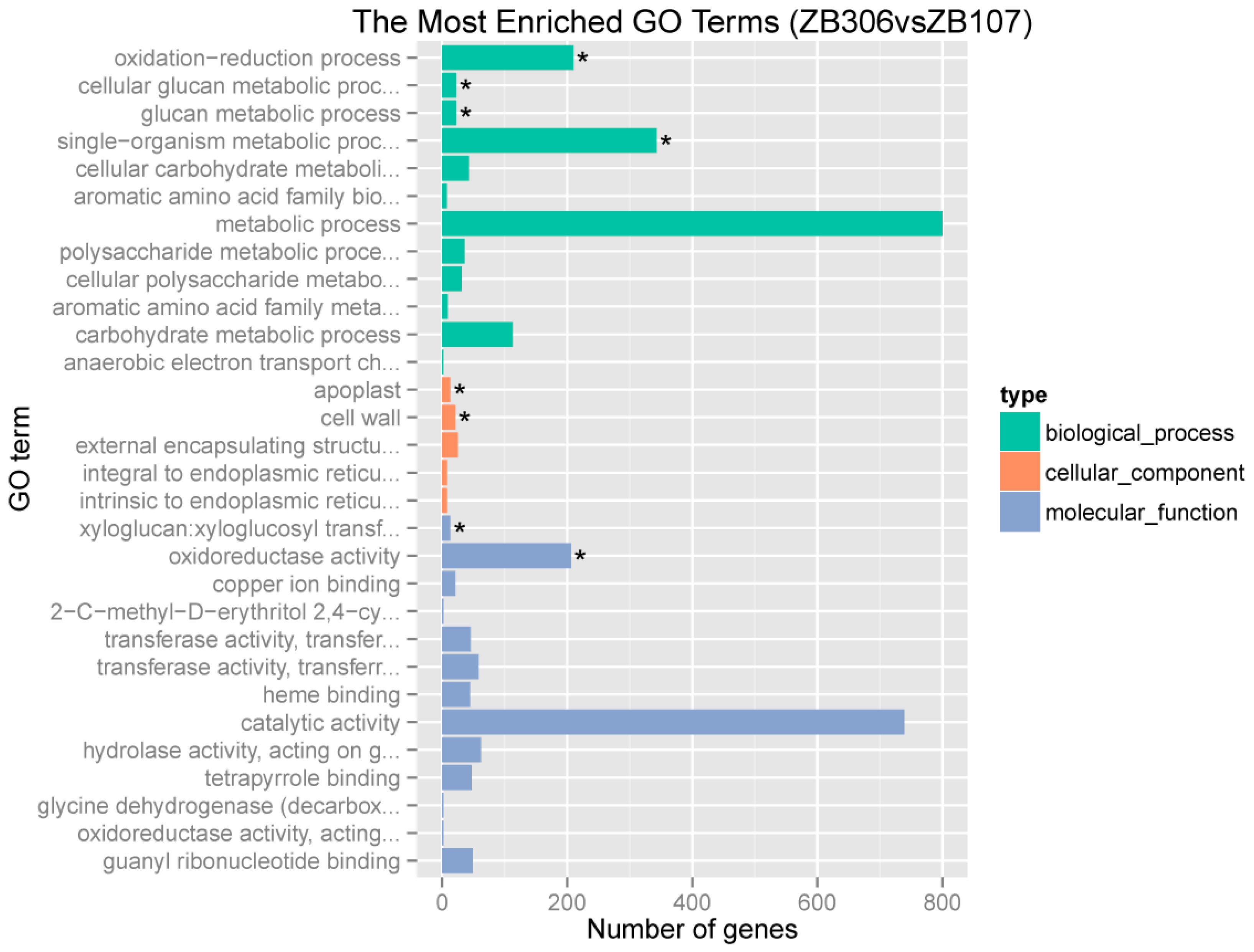
2.2. Identification of Seed Coat Specially Expressed Genes and Global Analysis of the Differentially Expressed Genes in Seed Coat between ZB107 and ZB306
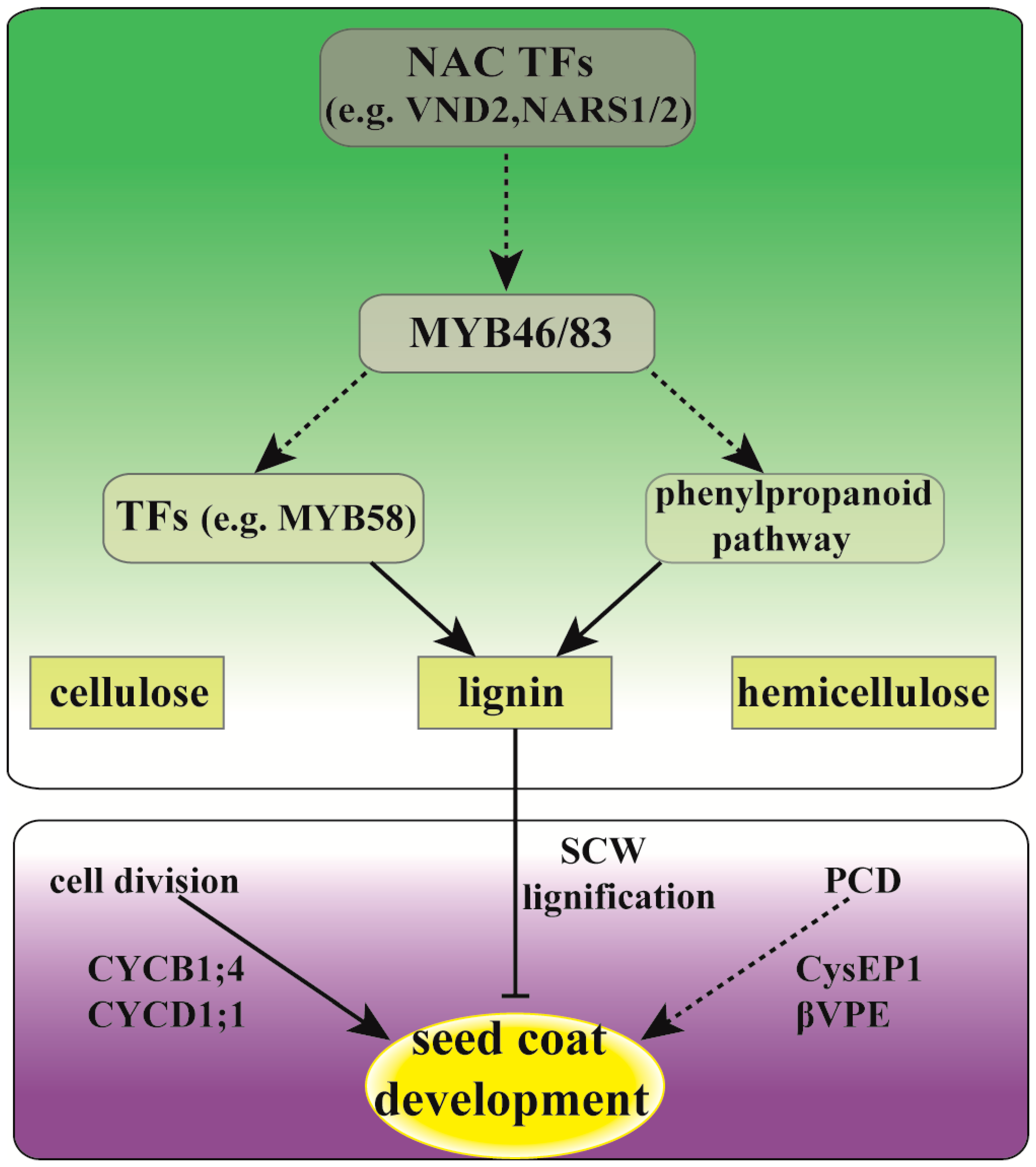
2.3. Differential Expression of Lignin Biosynthesis Related Genes in Seed Coat between ZB107 and ZB306
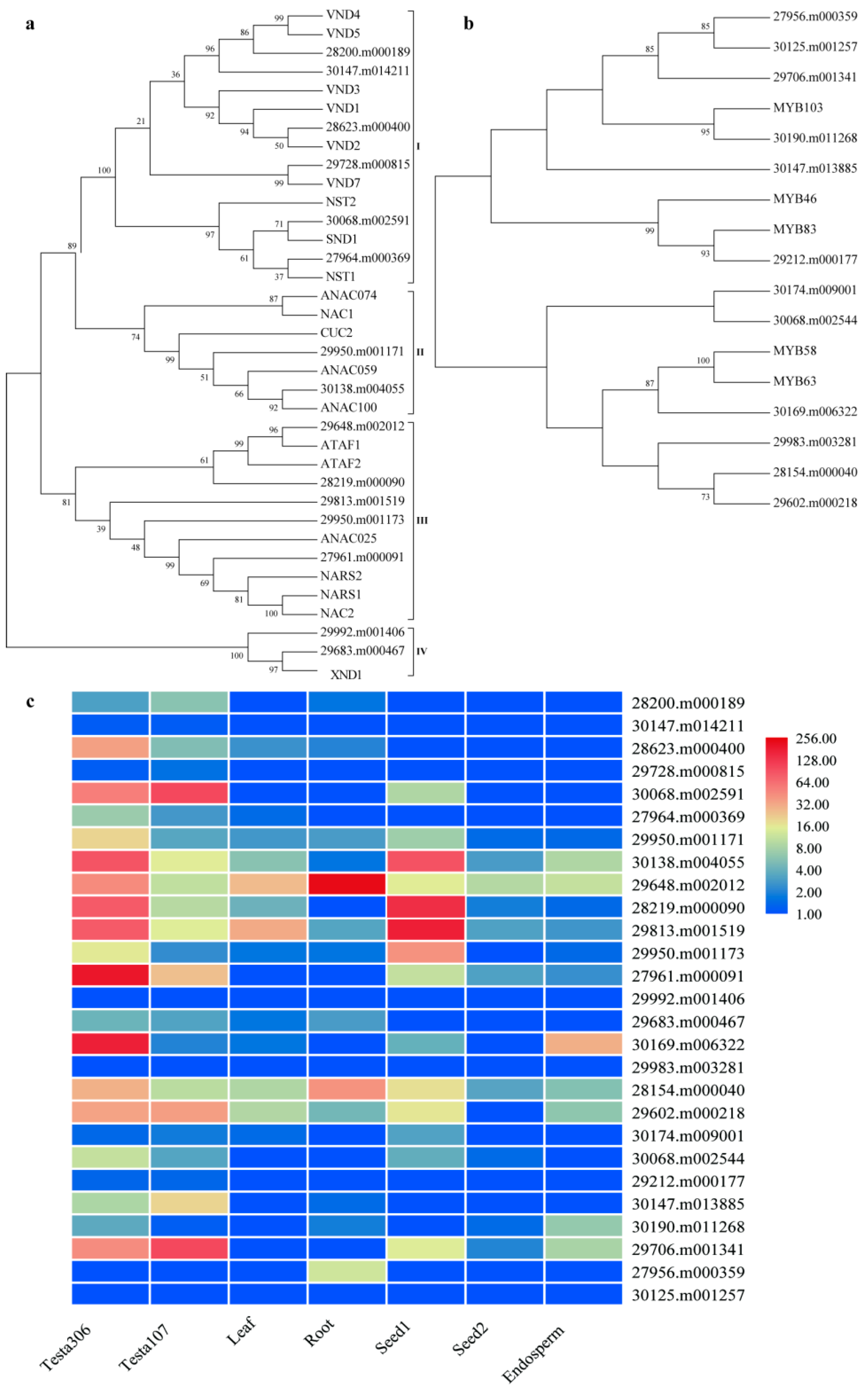
2.4. Differential Expression of Cell Development and PCD Related Genes in Seed Coats between ZB107 and ZB306
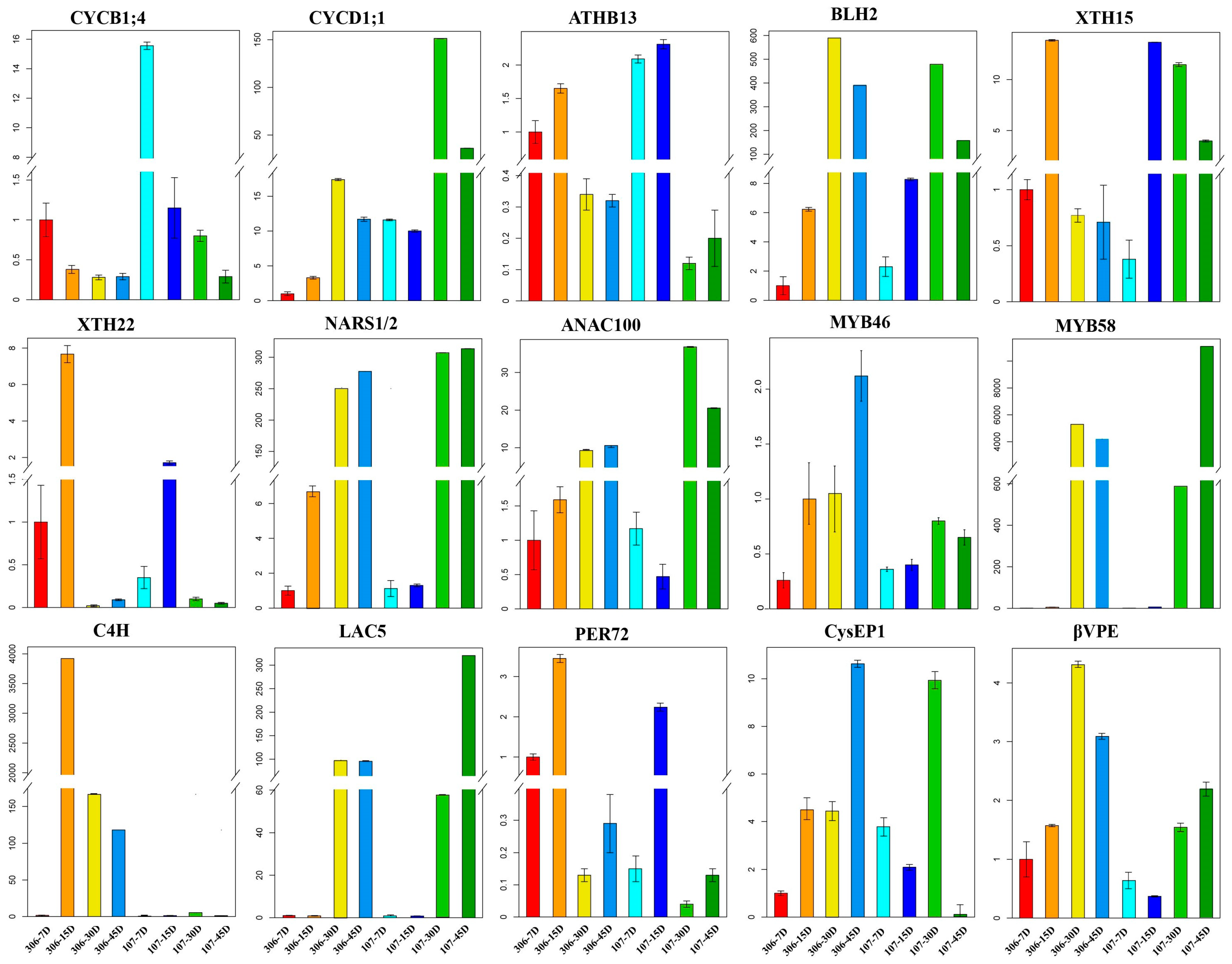
2.5. Experimental Validation of The Expression Patterns of Seed Coat Development-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Histological Analysis
4.3. Determination of Seed Coat Fiber Composition and Contents
4.4. RNA Extraction, Cdna Preparation and Illumina Sequencing
4.5. Analysis of DGE Data
4.6. Phylogenetic and Gene Expression Analysis
4.7. qRT-PCR
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DGE | digital gene expression |
| PCD | programmed cell death |
| 4CL | 4-coumarate-CoA ligase |
| C4H | cinnamate 4-hydroxylase |
| PRX | peroxidases |
| LAC | laccases |
| DAP | day after pollination |
| PAL | phenylalanine ammonia-lyase |
| CCR | cinnamoyl CoA reductase |
| CCoAOMT | caffeoyl-CoA O-methyltransferase |
| TF | transcription factor |
| SCW | secondary cell wall |
| NST | NAC Secondary wall Thickening promoting factor |
| VND | Vascular-related NAC-domain |
| XTH | xyloglucan endotransglucosylase/hydrolase protein |
| BR | brassinosteroid |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| NDF | neutral detergent fibre |
| ADF | acid detergent fiber |
References
- Makkar, H.P.S. Biofuel Co-Products as Livestock Feed: Opportunities and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; pp. 264–266. [Google Scholar]
- Chaudhury, A.M.; Koltunow, A.; Payne, T.; Luo, M.; Tucker, M.R.; Dennis, E.S.; Peacock, W.J. Control of early seed development. Annu. Rev. Cell Dev. Biol. 2001, 17, 677–699. [Google Scholar] [CrossRef] [PubMed]
- Haughn, G.; Chaudhury, A. Genetic analysis of seed coat development in Arabidopsis. Trends Plant Sci. 2005, 10, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Berger, F.; Grini, P.E.; Schnittger, A. Endosperm: an integrator of seed growth and development. Curr. Opin. Plant Biol. 2006, 9, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borisjuk, L.; Neuberger, T.; Schwender, J.; Heinzel, N.; Sunderhaus, S.; Fuchs, J.; Hay, J.O.; Tschiersch, H.; Braun, H.P.; Denolf, P.; et al. Seed architecture shapes embryo metabolism in oilseed rape. Plant Cell 2013, 25, 1625–1640. [Google Scholar] [CrossRef] [PubMed]
- Lemontey, C.; Mousset-Declas, C.; Munier-Jolain, N.; Boutin, J.P. Maternal genotype influences pea seed size by controlling both mitotic activity during early embryogenesis and final endoreduplication level/cotyledon cell size in mature seed. J. Exp. Bot. 2000, 51, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joet, T.; Laffargue, A.; Salmona, J.; Doulbeau, S.; Descroix, F.; Bertrand, B.; Lashermes, P.; Dussert, S. Regulation of galactomannan biosynthesis in coffee seeds. J. Exp. Bot. 2014, 65, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hirsch, C.N.; Sekhon, R.S.; de Leon, N.; Kaeppler, S.M. Evidence for maternal control of seed size in maize from phenotypic and transcriptional analysis. J. Exp. Bot. 2016, 67, 1907–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.K.; Wu, K.; Yuan, Q.B.; Liu, X.Y.; Liu, Z.B.; Lin, X.Y.; Zeng, R.Z.; Zhu, H.T.; Dong, G.J.; Qian, Q.; et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, L.; Corke, F.; Smith, C.; Bevan, M.W. Control of final seed and organ size by the DA1 gene family in Arabidopsis thaliana. Genes Dev. 2008, 22, 1331–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Dennis, E.S.; Berger, F.; Peacock, W.J.; Chaudhury, A. MINISEED3 (MINI3), a WRKY family gene, and HAIKU2 (IKU2), a leucine-rich repeat (LRR) KINASE gene, are regulators of seed size in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 17531–17536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimaru, K.; Hirotsu, N.; Madoka, Y.; Murakami, N.; Hara, N.; Onodera, H.; Kashiwagi, T.; Ujiie, K.; Shimizu, B.; Onishi, A.; et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat. Genet. 2013, 45, 707–711. [Google Scholar] [CrossRef]
- Figueiredo, D.D.; Kohler, C. Signalling events regulating seed coat development. Biochem. Soc. Trans. 2014, 42, 358–363. [Google Scholar] [CrossRef]
- Adamski, N.M.; Anastasiou, E.; Eriksson, S.; O’Neill, C.M.; Lenhard, M. Local maternal control of seed size by KLUH/CYP78A5-dependent growth signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 20115–20120. [Google Scholar] [CrossRef]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef]
- Greenwood, J.S.; Bewley, J.D. Seed Development in Ricinus communis (Castor Bean). 1. Descriptive Morphology. Can. J. Bot. 1982, 60, 1751–1760. [Google Scholar] [CrossRef]
- Transeau, E.N. General Botany; Discovery Publishing House: New Delhi, India, 1994; pp. 257–258. [Google Scholar]
- Xu, W.; Dai, M.Y.; Li, F.; Liu, A.Z. Genomic imprinting, methylation and parent-of-origin effects in reciprocal hybrid endosperm of castor bean. Nucleic Acids Res. 2014, 42, 6987–6998. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Yang, T.; Dong, X.; Li, D.Z.; Liu, A.Z. Genomic DNA methylation analyses reveal the distinct profiles in Castor Bean seeds with persistent endosperms. Plant Physiol. 2016, 171, 1242–1258. [Google Scholar] [CrossRef]
- Xu, W.; Li, F.; Ling, L.Z.; Liu, A.Z. Genome-wide survey and expression profiles of the AP2/ERF family in castor bean (Ricinus communis L.). BMC Genom. 2013, 14, 785. [Google Scholar] [CrossRef]
- Weber, H.; Borisjuk, L.; Wobus, U. Controlling seed development and seed size in Vicia faba: A role for seed coat-associated invertases and carbohydrate state. Plant J. 1996, 10, 823–834. [Google Scholar] [CrossRef]
- Smykal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 351. [Google Scholar]
- Tobimatsu, Y.; Chen, F.; Nakashima, J.; Escamilla-Trevino, L.L.; Jackson, L.; Dixon, R.A.; Ralph, J. Coexistence but independent biosynthesis of catechyl and guaiacyl/syringyl lignin polymers in seed coats. Plant Cell 2013, 25, 2587–2600. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, E.; Matus, J.T.; Finezzo, L.; Zenoni, S.; Loyola, R.; Guzzo, F.; Schlechter, R.; Ageorges, A.; Arce-Johnson, P.; Tornielli, G.B. The phenylpropanoid pathway is controlled at different branches by a set of R2R3-MYB C2 repressors in grapevine. Plant Physiol. 2015, 167, 1448–1470. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Ralph, J.; Akiyama, T.; Lu, F.C.; Pazo, J.R.; Kim, H.; Christensen, J.H.; Van Reusel, B.; Storme, V.; De Rycke, R.; et al. Engineering traditional monolignols out of lignin by concomitant up-regulation of F5H1 and down-regulation of COMT in Arabidopsis. Plant J. 2010, 64, 885–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, J.; Fernandez-Perez, F.; Yebra, T.; Novo-Uzal, E.; Pomar, F.; Pedreno, M.A.; Cuello, J.; Guera, A.; Esteban-Carrasco, A.; Zapata, J.M. Bioinformatic and functional characterization of the basic peroxidase 72 from Arabidopsis thaliana involved in lignin biosynthesis. Planta 2013, 237, 1599–1612. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Zhang, S.C.; Yu, Y.; Luo, Y.C.; Liu, Q.; Ju, C.L.; Zhang, Y.C.; Qu, L.H.; Lucas, W.J.; Wang, X.J.; et al. MiR397b regulates both lignin content and seed number in Arabidopsis via modulating a laccase involved in lignin biosynthesis. Plant Biotechnol. J. 2014, 12, 1132–1142. [Google Scholar] [CrossRef] [Green Version]
- Zhong, R.Q.; Demura, T.; Ye, Z.H. SND1, a NAC domain transcription factor, is a key regulator of secondary wall synthesis in fibers of Arabidopsis. Plant Cell 2006, 18, 3158–3170. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Zhou, J.; McCarthy, R.L.; Ye, Z.H. A battery of transcription factors involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell 2008, 20, 2763–2782. [Google Scholar] [CrossRef] [PubMed]
- Barros, J.; Serk, H.; Granlund, I.; Pesquet, E. The cell biology of lignification in higher plants. Ann. Bot. 2015, 115, 1053–1074. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.G. Cellulose biosynthesis and deposition in higher plants. New Phytol. 2008, 178, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef]
- Xu, W.; Yang, T.Q.; Wang, B.; Han, B.; Zhou, H.K.; Wang, Y.; Li, D.Z.; Liu, A.Z. Differential expression networks and inheritance patterns of long non-coding RNAs in castor bean seeds. Plant J. 2018, 95, 324–340. [Google Scholar] [CrossRef]
- Zhao, Q.; Nakashima, J.; Chen, F.; Yin, Y.B.; Fu, C.X.; Yun, J.F.; Shao, H.; Wang, X.Q.; Wang, Z.Y.; Dixon, R.A. LACCASE is necessary and nonredundant with peroxidase for lignin polymerization during vascular development in Arabidopsis. Plant Cell 2013, 25, 3976–3987. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.L.; Lee, C.H.; Zhong, R.Q.; Ye, Z.H. MYB58 and MYB63 are transcriptional activators of the lignin biosynthetic pathway during secondary cell wall formation in Arabidopsis. Plant Cell 2009, 21, 248–266. [Google Scholar] [CrossRef]
- Zhong, R.; Richardson, E.A.; Ye, Z.H. The MYB46 transcription factor is a direct target of SND1 and regulates secondary wall biosynthesis in Arabidopsis. Plant Cell 2007, 19, 2776–2792. [Google Scholar] [CrossRef]
- McCarthy, R.L.; Zhong, R.Q.; Ye, Z.H. MYB83 is a direct target of SND1 and acts redundantly with MYB46 in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell Physiol. 2009, 50, 1950–1964. [Google Scholar] [CrossRef] [PubMed]
- Groover, A.; Jones, A.M. Tracheary element differentiation uses a novel mechanism coordinating programmed cell death and secondary cell wall synthesis. Plant Physiol. 1999, 119, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Kushalappa, K.; Godt, D.; Pidkowich, M.S.; Pastorelli, S.; Hepworth, S.R.; Haughn, G.W. The Arabidopsis BEL1-LIKE HOMEODOMAIN proteins SAW1 and SAW2 act redundantly to regulate KNOX expression spatially in leaf margins. Plant Cell 2007, 19, 2719–2735. [Google Scholar] [CrossRef] [PubMed]
- Ribone, P.A.; Capella, M.; Chan, R.L. Functional characterization of the homeodomain leucine zipper I transcription factor AtHB13 reveals a crucial role in Arabidopsis development. J. Exp. Bot. 2015, 66, 5929–5943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.; Wang, X.; Yang, M.; Yang, L.; He, G.; Deng, X.W. A new regulator of seed size control in Arabidopsis identified by a genome-wide association study. New Phytol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rojas, M.; Meneses, M.; Oviedo, K.; Carrasco, C.; Defilippi, B.; Gonzalez-Aguero, M.; Leon, G.; Hinrichsen, P. Exogenous gibberellic acid application induces the overexpression of key genes for pedicel lignification and an increase in berry drop in table grape. Plant Physiol. Biochem. 2018, 126, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Li, J.; Ban, L.P.; Wu, Y.D.; Wu, X.M.; Wang, Y.Q.; Wen, H.Y.; Chapurin, V.; Dzyubenko, N.; Li, Z.Y.; et al. Functional characterization of a gibberellin receptor and its application in alfalfa biomass improvement. Sci. Rep. 2017, 7, 41296. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.; Simpson, D.; Gietl, C. Programmed cell death in castor bean endosperm is associated with the accumulation and release of a cysteine endopeptidase from ricinosomes. Proc. Natl. Acad. Sci. USA 1999, 96, 14159–14164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, A.J.; Soares, E.L.; Costa, J.H.; Costa, W.L.; Soares, A.A.; Nogueira, F.C.; Domont, G.B.; Campos, F.A. Differential expression of cysteine peptidase genes in the inner integument and endosperm of developing seeds of Jatropha curcas L. (Euphorbiaceae). Plant Sci. 2013, 213, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Y. Signaling pathways of seed size control in plants. Curr. Opin. Plant Biol. 2016, 33, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.B.; Fan, C.C.; Xing, Y.Z.; Jiang, Y.H.; Luo, L.J.; Sun, L.; Shao, D.; Xu, C.J.; Li, X.H.; Xiao, J.H.; et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 2011, 43, 1266–1269. [Google Scholar] [CrossRef]
- Kang, Y.J.; Shim, K.C.; Lee, H.S.; Jeon, Y.A.; Kim, S.H.; Kang, J.W.; Yun, Y.T.; Park, I.K.; Ahn, S.N. Fine mapping and candidate gene analysis of the quantitative trait locus gw8.1 associated with grain length in rice. Genes Genom. 2018, 40, 389–397. [Google Scholar] [CrossRef]
- Wang, Y.X.; Xiong, G.S.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.X.; Zeng, L.J.; Xu, E.B.; Xu, J.; et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef]
- Si, L.Z.; Chen, J.Y.; Huang, X.H.; Gong, H.; Luo, J.H.; Hou, Q.Q.; Zhou, T.Y.; Lu, T.T.; Zhu, J.J.; Shangguan, Y.Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef]
- Dewitte, W.; Riou-Khamlichi, C.; Scofield, S.; Healy, J.M.S.; Jacqmard, A.; Kilby, N.J.; Murray, J.A.H. Altered cell cycle distribution, hyperplasia, and inhibited differentiation in Arabidopsis caused by the D-type cyclin CYCD3. Plant Cell 2003, 15, 79–92. [Google Scholar] [CrossRef]
- Jofuku, K.D.; Omidyar, P.K.; Gee, Z.; Okamuro, J.K. Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc. Natl. Acad. Sci. USA 2005, 102, 3117–3122. [Google Scholar] [CrossRef] [Green Version]
- Garcia, D.; Fitz Gerald, J.N.; Berger, F. Maternal control of integument cell elongation and zygotic control of endosperm growth are coordinated to determine seed size in Arabidopsis. Plant Cell 2005, 17, 52–60. [Google Scholar] [CrossRef]
- Xu, W.; Purugganan, M.M.; Polisensky, D.H.; Antosiewicz, D.M.; Fry, S.C.; Braam, J. Arabidopsis TCH4, regulated by hormones and the rnvironment, encodes a xyloglucan endotransglycosylase. Plant Cell 1995, 7, 1555–1567. [Google Scholar] [CrossRef]
- Huang, H.Y.; Jiang, W.B.; Hu, Y.W.; Wu, P.; Zhu, J.Y.; Liang, W.Q.; Wang, Z.Y.; Lin, W.H. BR signal influences arabidopsis ovule and seed number through regulating related genes expression by BZR1. Mol. Plant 2013, 6, 456–469. [Google Scholar] [CrossRef]
- Chen, F.; Tobimatsu, Y.; Jackson, L.; Nakashima, J.; Ralph, J.; Dixon, R.A. Novel seed coat lignins in the Cactaceae: Structure, distribution and implications for the evolution of lignin diversity. Plant J. 2013, 73, 201–211. [Google Scholar] [CrossRef]
- Wang, J.P.; Matthews, M.L.; Williams, C.M.; Shi, R.; Yang, C.M.; Tunlaya-Anukit, S.; Chen, H.C.; Li, Q.Z.; Liu, J.; Lin, C.Y.; et al. Improving wood properties for wood utilization through multi-omics integration in lignin biosynthesis. Nat. Commun. 2018, 9, 1579. [Google Scholar] [CrossRef] [Green Version]
- Rohde, A.; Morreel, K.; Ralph, J.; Goeminne, G.; Hostyn, V.; De Rycke, R.; Kushnir, S.; Van Doorsselaere, J.; Joseleau, J.P.; Vuylsteke, M.; et al. Molecular phenotyping of the pal1 and pal2 mutants of Arabidopsis thaliana reveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism. Plant Cell 2004, 16, 2749–2771. [Google Scholar] [CrossRef]
- Lee, Y.; Rubio, M.C.; Alassimone, J.; Geldner, N. A mechanism for localized lignin deposition in the endodermis. Cell 2013, 153, 402–412. [Google Scholar] [CrossRef]
- Liang, M.; Davis, E.; Gardner, D.; Cai, X.; Wu, Y. Involvement of AtLAC15 in lignin synthesis in seeds and in root elongation of Arabidopsis. Planta 2006, 224, 1185–1196. [Google Scholar] [CrossRef]
- Rao, X.L.; Dixon, R.A. Current models for transcriptional regulation of secondary cell wall biosynthesis in grasses. Front. Plant Sci. 2018, 9, 399. [Google Scholar] [CrossRef]
- Dominguez, F.; Cejudo, F.J. Programmed cell death (PCD): An essential process of cereal seed development and germination. Front. Plant Sci. 2014, 5, 366. [Google Scholar] [CrossRef]
- Lopez-Fernandez, M.P.; Maldonado, S. Programmed cell death in seeds of angiosperms. J. Integr. Plant Biol. 2015, 57, 996–1002. [Google Scholar] [CrossRef]
- Kunieda, T.; Mitsuda, N.; Ohme-Takagi, M.; Takeda, S.; Aida, M.; Tasaka, M.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. NAC family proteins NARS1/NAC2 and NARS2/NAM in the outer integument regulate embryogenesis in Arabidopsis. Plant Cell 2008, 20, 2631–2642. [Google Scholar] [CrossRef]
- Ohashi-Ito, K.; Oda, Y.; Fukuda, H. Arabidopsis VASCULAR-RELATED NAC-DOMAIN6 directly regulates the genes that govern programmed cell death and secondary wall formation during xylem differentiation. Plant Cell 2010, 22, 3461–3473. [Google Scholar] [CrossRef]
- Courtois-Moreau, C.L.; Pesquet, E.; Sjodin, A.; Muniz, L.; Bollhoner, B.; Kaneda, M.; Samuels, L.; Jansson, S.; Tuominen, H. A unique program for cell death in xylem fibers of Populus stem. Plant J. 2009, 58, 260–274. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Wang, L.K.; Feng, Z.X.; Wang, X.; Wang, X.W.; Zhang, X.G. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔTC method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Gene ID | ZB306 2 | ZB107 3 | Log2FC | p-Value | Gene Annotation |
|---|---|---|---|---|---|---|
| PAL | 30078.m002319 | 596.57 | 44.30 | 3.75 | 1.1 × 10−119 | Phenylalanine ammonia-lyase |
| PAL | 28507.m000156 | 1307.44 | 73.76 | 4.15 | 3.5 × 10−277 | Phenylalanine ammonia-lyase |
| C4H | 43540.m000048 | 84.22 | 14.64 | 2.52 | 4.32 × 10−13 | Trans-cinnamate 4-monooxygenase |
| 4CL | 28429.m000109 | 190.24 | 26.69 | 2.83 | 4.07 × 10−31 | 4-coumarate:CoA ligase 1 |
| 4CL | 30131.m006921 | 2568.80 | 84.70 | 4.92 | 0 | 4-coumarate:CoA ligase 1 |
| 4CL | 30073.m002251 | 122.55 | 20.20 | 2.60 | 6.14 × 10−19 | 4-coumarate:CoA ligase 2 |
| CCR | 29588.m000854 | 410.02 | 145.68 | 1.49 | 3.51 × 10−29 | Cinnamoyl-CoA reductase 1 |
| CCoAOMT | 29968.m000634 | 202.55 | 71.36 | 1.51 | 2.19 × 10−15 | caffeoyl-CoA O-methyltransferase |
| PRX10 | 29676.m001629 | 154.37 | 42.63 | 1.86 | 7.32 × 10−16 | Peroxidase 10 |
| PRX19 | 29900.m001566 | 0.00 | 27.62 | −4.97 | 5.23 × 10−08 | Peroxidase 19 |
| PRX47 | 29983.m003295 | 3.87 | 27.99 | −2.86 | 5.1 × 10−06 | Peroxidase 47 |
| PRX64 | 30170.m014275 | 0.00 | 52.27 | −8.22 | 1.97 × 10−12 | Peroxidase 64 |
| PRX64 | 28962.m000432 | 7.21 | 34.10 | −2.24 | 1.04 × 10−05 | Peroxidase 64 |
| PRX72 | 29634.m002067 | 154.73 | 2519.30 | −4.03 | 0 | Peroxidase 72 |
| LAC4 | 29610.m000409 | 113.41 | 0.00 | 9.26 | 3.53 × 10−22 | Laccase 4 |
| LAC5 | 29751.m001786 | 37.27 | 131.03 | −1.81 | 4.23 × 10−14 | Laccase 5 |
| LAC17 | 30004.m000428 | 80.18 | 1604.66 | −4.32 | 0 | Laccase 17 |
| Pathway | Gene 1 | Gene ID | ZB306 2 | ZB107 3 | Log2FC | Gene Annotation |
|---|---|---|---|---|---|---|
| ABA | PP2C | 29739.m003582 | 38.33 | 78.40 | −1.03 | protein phosphatase 2c |
| ABA | ABF | 29801.m003176 | 21.63 | 51.15 | −1.24 | DNA binding protein |
| Auxin | AUX|IAA | 29598.m000460 | 48.70 | 114.35 | −1.23 | Auxin-responsive protein IAA16 |
| Auxin | AUX|IAA | 29841.m002748 | 3.87 | 59.12 | −3.93 | Auxin-induced protein AUX22 |
| Auxin | AUX|IAA | 29844.m003174 | 8.44 | 52.27 | −2.63 | Auxin-responsive protein IAA7 |
| Auxin | AUX1 | 29908.m006146 | 5.45 | 26.13 | −2.26 | amino acid transporter |
| Auxin | AUX1 | 29969.m000264 | 3.52 | 25.21 | −2.84 | amino acid transporter |
| Auxin | GH3 | 30129.m000366 | 149.98 | 32.99 | 2.18 | Indole-3-acetic acid-amido synthetase GH3.17 |
| Auxin | SAUR | 30131.m007151 | 6.51 | 26.50 | −2.03 | Indole-3-acetic acid-induced protein ARG7 |
| Auxin | PIN3 | 29816.m000677 | 7.91 | 77.47 | −3.29 | Auxin efflux carrier component 3 |
| BR | CYCD3 | 29801.m003121 | 4.04 | 48.37 | −3.58 | cyclin d |
| BR | TCH4 | 30179.m000569 | 0.00 | 241.12 | −8.10 | Xyloglucan endotransglucosylase/hydrolase protein 22 precursor |
| Cytokinine | A-ARR | 28094.m000169 | 4.40 | 22.98 | −2.39 | two-component response regulator ARR-A |
| Cytokinine | A-ARR | 29908.m006123 | 122.73 | 355.48 | −1.53 | Two-component response regulator ARR9 |
| Ethylene | CTR1 | 29428.m000323 | 93.54 | 25.21 | 1.89 | serine/threonine-protein kinase CTR1 |
| Ethylene | ETR | 29603.m000534 | 632.27 | 52.82 | 3.58 | ethylene receptor |
| Ethylene | EBF1/2 | 28320.m001145 | 325.80 | 39.11 | 3.06 | EIN3-binding F-box protein 1 |
| Ethylene | EBF1/2 | 29848.m004629 | 258.29 | 64.31 | 2.01 | EIN3-binding F-box protein 1 |
| Ethylene | ERF1/2 | 29895.m000321 | 13.71 | 0.00 | 6.21 | Ethylene-responsive transcription factor 1B |
| Ethylene | ETR | 29986.m001660 | 111.30 | 45.04 | 1.31 | ethylene receptor |
| GA | DELLA | 28677.m000055 | 38.33 | 79.51 | −1.05 | DELLA protein GAI |
| GA | DELLA | 30131.m007029 | 16.00 | 0.00 | 6.43 | DELLA protein GAI |
| GA | GID1 | 29703.m001506 | 54.68 | 18.16 | 1.59 | Gibberellin receptor GID1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, A.; Wang, Z.; Zhang, Y.; Li, F.; Liu, A. Global Gene Expression of Seed Coat Tissues Reveals a Potential Mechanism of Regulating Seed Size Formation in Castor Bean. Int. J. Mol. Sci. 2019, 20, 1282. https://doi.org/10.3390/ijms20061282
Yu A, Wang Z, Zhang Y, Li F, Liu A. Global Gene Expression of Seed Coat Tissues Reveals a Potential Mechanism of Regulating Seed Size Formation in Castor Bean. International Journal of Molecular Sciences. 2019; 20(6):1282. https://doi.org/10.3390/ijms20061282
Chicago/Turabian StyleYu, Anmin, Zaiqing Wang, Yang Zhang, Fei Li, and Aizhong Liu. 2019. "Global Gene Expression of Seed Coat Tissues Reveals a Potential Mechanism of Regulating Seed Size Formation in Castor Bean" International Journal of Molecular Sciences 20, no. 6: 1282. https://doi.org/10.3390/ijms20061282