Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Placental Gene Expression and Fetal Antioxidant Status, DNA-Methylation and Phenotype

 , , , , , , and
, , , , , , and
Abstract
:
1. Introduction
2. Results
2.1. Maternal and Fetal Plasma Hydroxytyrosol Concentrations
2.2. Placental Candidate Gene Expression
2.3. Fetal Antioxidant Status
2.4. Fetal Development
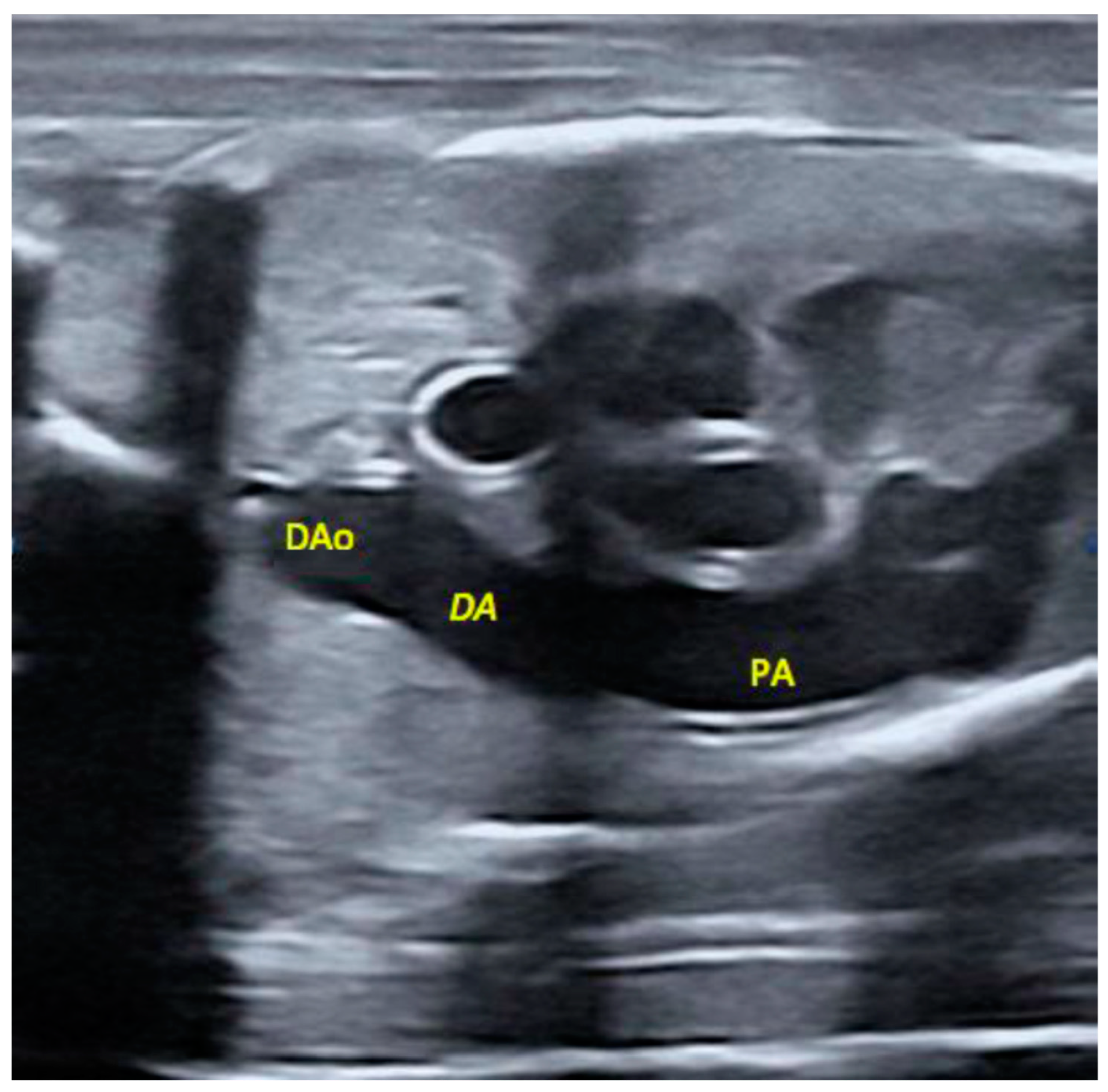
2.5. Morphological Appearance of Fetal Descending Aorta and Ductus Arteriosus
2.6. Fetal Muscle Fibers
2.7. Fetal Metabolic Status (Glycemic and Lipid Profiles)
2.8. Fetal DNA Methylation
3. Discussion
4. Methods
4.1. Ethics Statement
4.2. Animals and Experimental Procedures
4.3. Measuring, Weighting and Sampling of Fetuses
4.4. Evaluation of Maternal and Fetal Hydroxytyrosol Concentrations
4.5. Evaluation of Maternal and Fetal Antioxidant Status
4.6. Evaluation of Maternal and Fetal Metabolic Status
4.7. Evaluation of Placental Candidate Gene Expression
4.8. Evaluation of Fetal DNA Methylation
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Statti, G.A.; Menichini, F. Biological and pharmacological activities of iridoids: Recent developments. Mini Rev. Med. Chem. 2008, 8, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Rigacci, S.; Stefani, M. Nutraceutical Properties of Olive Oil Polyphenols. An Itinerary from Cultured Cells through Animal Models to Humans. Int. J. Mol. Sci. 2016, 17, 843. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Gomez, M.; Garcia-Contreras, C.; Torres-Rovira, L.; Pesantez, J.L.; Gonzalez-Añover, P.; Gomez-Fidalgo, E.; Sanchez-Sanchez, R.; Ovilo, C.; Isabel, B.; Astiz, S.; et al. Polyphenols and IUGR pregnancies: Maternal hydroxytyrosol supplementation improves prenatal and early-postnatal growth and metabolism of the offspring. PLoS ONE 2017, 12, e0177593. [Google Scholar] [CrossRef] [PubMed]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Narang, M.; Banerjee, B.D.; Basu, S. Oxidative stress in term small for gestational age neonates born to undernourished mothers: A case control study. BMC Pediatr. 2004, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Kamath, U.; Rao, G.; Kamath, S.U.; Rai, L. Maternal and fetal indicators of oxidative stress during intrauterine growth retardation (IUGR). Indian J. Clin. Biochem. 2006, 21, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Li, H.P.; Chen, X.; Li, M.Q. Gestational diabetes induces chronic hypoxia stress and excessive inflammatory response in murine placenta. Int. J. Clin. Exp. Pathol. 2013, 6, 650–659. [Google Scholar] [PubMed]
- Prior, R.L.; Gu, L.; Wu, X.; Jacob, R.A.; Sotoudeh, G.; Kader, A.A.; Cook, R.A. Plasma antioxidant capacity changes following a meal as a measure of the ability of a food to alter in vivo antioxidant status. J. Am. Coll. Nutr. 2007, 26, 170–181. [Google Scholar] [CrossRef]
- Ly, C.; Yockell-Lelievre, J.; Ferraro, Z.M.; Arnason, J.T.; Ferrier, J.; Gruslin, A. The effects of dietary polyphenols on reproductive health and early development. Hum. Reprod. Update 2015, 21, 228–248. [Google Scholar] [CrossRef]
- Chen, B.; Tuuli, M.G.; Longtine, M.S.; Shin, J.S.; Lawrence, R.; Inder, T.; Michael Nelson, D. Pomegranate juice and punicalagin attenuate oxidative stress and apoptosis in human placenta and in human placental trophoblasts. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1142–E1152. [Google Scholar] [CrossRef] [Green Version]
- Burton, G.J.; Yung, H.W.; Cindrova-Davies, T.; Charnock-Jones, D.S. Placental endoplasmic reticulum stress and oxidative stress in the pathophysiology of unexplained intrauterine growth restriction and early onset preeclampsia. Placenta 2009, 30 (Suppl. A), S43–S48. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, E.; Burton, G.J. The role of oxidative stress in placental-related diseases of pregnancy. J. Gynecol. Obstet. Biol. Reprod. (Paris) 2016, 45, 775–785. [Google Scholar] [CrossRef]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granados-Principal, S.; Quiles, J.L.; Ramirez-Tortosa, C.L.; Sanchez-Rovira, P.; Ramirez-Tortosa, M.C. Hydroxytyrosol: From laboratory investigations to future clinical trials. Nutr. Rev. 2010, 68, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, L.; Cicerale, S. The Health Benefiting Mechanisms of Virgin Olive Oil Phenolic Compounds. Molecules 2016, 21, 1734. [Google Scholar] [CrossRef] [PubMed]
- Zielinsky, P.; Busato, S. Prenatal effects of maternal consumption of polyphenol-rich foods in late pregnancy upon fetal ductus arteriosus. Birth Defects Res. C Embryo Today 2013, 99, 256–274. [Google Scholar] [CrossRef] [Green Version]
- Abdel Mohsen, A.H.; Amin, A.S. Risk factors and outcomes of persistent pulmonary hypertension of the newborn in neonatal intensive care unit of Al-minya university hospital in egypt. J. Clin. Neonatol. 2013, 2, 78–82. [Google Scholar] [CrossRef]
- Babaoglu, K.; Cakiroglu, Y.; Altun, G.; Doger, E.; Oguz, D. Intrauterine idiopathic severe ductal constriction diagnosed by fetal echocardiography: A cause of hydrops fetalis. Anadolu Kardiyol Derg. 2013, 13, 496–497. [Google Scholar] [CrossRef]
- Tarcan, A.; Gurakan, B.; Yildirim, S.; Ozkiraz, S.; Bilezikci, B. Persistent pulmonary hypertension in a premature newborn after 16 hours of antenatal indomethacin exposure. J. Perinat. Med. 2004, 32, 98–99. [Google Scholar] [CrossRef]
- Vanhees, K.; van Schooten, F.J.; van Waalwijk van Doorn-Khosrovani, S.B.; van Helden, S.; Munnia, A.; Peluso, M.; Briede, J.J.; Haenen, G.R.; Godschalk, R.W. Intrauterine exposure to flavonoids modifies antioxidant status at adulthood and decreases oxidative stress-induced DNA damage. Free Radic. Biol. Med. 2013, 57, 154–161. [Google Scholar] [CrossRef]
- Cao, L.; Tan, C.; Meng, F.; Liu, P.; Reece, E.A.; Zhao, Z. Amelioration of intracellular stress and reduction of neural tube defects in embryos of diabetic mice by phytochemical quercetin. Sci. Rep. 2016, 6, 21491. [Google Scholar] [CrossRef] [PubMed]
- Moritz, K.M.; Cuffe, J.S.; Wilson, L.B.; Dickinson, H.; Wlodek, M.E.; Simmons, D.G.; Denton, K.M. Review: Sex specific programming: A critical role for the renal renin-angiotensin system. Placenta 2010, 31, S40–S46. [Google Scholar] [CrossRef]
- Aiken, C.E.; Ozanne, S.E. Sex differences in developmental programming models. Reproduction 2013, 145, R1–R13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clifton, V.L.; Murphy, V.E. Maternal asthma as a model for examining fetal sex-specific effects on maternal physiology and placental mechanisms that regulate human fetal growth. Placenta 2004, 25 (Suppl. A), S45–S52. [Google Scholar] [CrossRef] [PubMed]
- Stark, R.; Wolf, O.T.; Tabbert, K.; Kagerer, S.; Zimmermann, M.; Kirsch, P.; Schienle, A.; Vaitl, D. Influence of the stress hormone cortisol on fear conditioning in humans: Evidence for sex differences in the response of the prefrontal cortex. NeuroImage 2006, 32, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Cuffe, J.S.; Dickinson, H.; Simmons, D.G.; Moritz, K.M. Sex specific changes in placental growth and MAPK following short term maternal dexamethasone exposure in the mouse. Placenta 2011, 32, 981–989. [Google Scholar] [CrossRef]
- Cuffe, J.S.; O’Sullivan, L.; Simmons, D.G.; Anderson, S.T.; Moritz, K.M. Maternal corticosterone exposure in the mouse has sex-specific effects on placental growth and mRNA expression. Endocrinology 2012, 153, 5500–5511. [Google Scholar] [CrossRef]
- Pompella, A.; Visvikis, A.; Paolicchi, A.; de Tata, V.; Casini, A.F. The changing faces of glutathione, a cellular protagonist. Biochem. Pharmacol. 2003, 66, 1499–1503. [Google Scholar] [CrossRef]
- Barth, A.; Bauer, R.; Gedrange, T.; Walter, B.; Linss, C.; Klinger, W. Influence of hypoxia and hyperthermia upon peroxidative and glutathione status in growth-restricted newborn piglets. Exp. Toxicol. Pathol. 1998, 50, 31–33. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, J.; Wu, G. Glycine metabolism in animals and humans: Implications for nutrition and health. Amino Acids 2013, 45, 463–477. [Google Scholar] [CrossRef]
- Kouka, P.; Priftis, A.; Stagos, D.; Angelis, A.; Stathopoulos, P.; Xinos, N.; Skaltsounis, A.L.; Mamoulakis, C.; Tsatsakis, A.M.; Spandidos, D.A.; et al. Assessment of the antioxidant activity of an olive oil total polyphenolic fraction and hydroxytyrosol from a Greek Olea europea variety in endothelial cells and myoblasts. Int. J. Mol. Med. 2017, 40, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Gimenez, J.L.; Pallardo, F.V. Maintenance of glutathione levels and its importance in epigenetic regulation. Front. Pharmacol. 2014, 5, 88. [Google Scholar] [PubMed]
- Zinellu, A.; Sotgiu, E.; Assaretti, S.; Sotgia, S.; Paliogiannis, P.; Pintus, G.; Mangoni, A.A.; Carru, C. Evaluation of Global Genomic DNA Methylation in Human Whole Blood by Capillary Electrophoresis UV Detection. J. Anal. Methods Chem. 2017, 2017, 4065892. [Google Scholar] [CrossRef] [PubMed]
- Chango, A.; Pogribny, I.P. Considering maternal dietary modulators for epigenetic regulation and programming of the fetal epigenome. Nutrients 2015, 7, 2748–2770. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Contreras, C.; Vazquez-Gomez, M.; Astiz, S.; Torres-Rovira, L.; Sanchez-Sanchez, R.; Gomez-Fidalgo, E.; Gonzalez, J.; Isabel, B.; Rey, A.; Ovilo, C.; et al. Ontogeny of Sex-Related Differences in Foetal Developmental Features, Lipid Availability and Fatty Acid Composition. Int. J. Mol. Sci. 2017, 18, 1171. [Google Scholar] [CrossRef] [PubMed]
- Cogollos, L.; Garcia-Contreras, C.; Vazquez-Gomez, M.; Astiz, S.; Sanchez-Sanchez, R.; Gomez-Fidalgo, E.; Ovilo, C.; Isabel, B.; Gonzalez-Bulnes, A. Effects of fetal genotype and sex on developmental response to maternal malnutrition. Reprod. Fertil. Dev. 2017, 29, 1155–1168. [Google Scholar] [CrossRef]
- Gonzalez-Bulnes, A.; Ovilo, C.; Lopez-Bote, C.J.; Astiz, S.; Ayuso, M.; Perez-Solana, M.L.; Sanchez-Sanchez, R.; Torres-Rovira, L. Gender-specific early postnatal catch-up growth after intrauterine growth retardation by food restriction in swine with obesity/leptin resistance. Reproduction 2012, 144, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Aberle, E.D. Myofiber differentiation in skeletal muscles of newborn runt and normal weight pigs. J. Anim. Sci. 1984, 59, 1651–1656. [Google Scholar] [CrossRef]
- Alvarenga, A.L.; Chiarini-Garcia, H.; Cardeal, P.C.; Moreira, L.P.; Foxcroft, G.R.; Fontes, D.O.; Almeida, F.R. Intra-uterine growth retardation affects birthweight and postnatal development in pigs, impairing muscle accretion, duodenal mucosa morphology and carcass traits. Reprod. Fertil. Dev. 2013, 25, 387–395. [Google Scholar] [CrossRef]
- Pardo, C.E.; Berard, J.; Kreuzer, M.; Bee, G. Intrauterine crowding impairs formation and growth of secondary myofibers in pigs. Animal 2013, 7, 430–438. [Google Scholar] [CrossRef]
- Zhang, X.M.; Guo, L.; Chi, M.H.; Sun, H.M.; Chen, X.W. Identification of active miRNA and transcription factor regulatory pathways in human obesity-related inflammation. BMC Bioinformatics 2015, 16, 76. [Google Scholar] [CrossRef] [PubMed]
- Mahan, D.C.; Watts, M.R.; St-Pierre, N. Macro- and micromineral composition of fetal pigs and their accretion rates during fetal development. J. Anim. Sci. 2009, 87, 2823–2832. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Bulnes, A.; Astiz, S.; Ovilo, C.; Lopez-Bote, C.J.; Torres-Rovira, L.; Barbero, A.; Ayuso, M.; Garcia-Contreras, C.; Vazquez-Gomez, M. Developmental Origins of Health and Disease in swine: Implications for animal production and biomedical research. Theriogenology 2016, 86, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Walsh Hentges, L.S.; Martin, R.J. Serum and lipoprotein lipids of fetal pigs and their dams during gestation as compared with man. Biol. Neonate 1987, 52, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Lewinska, A.; Wnuk, M.; Slota, E.; Bartosz, G. Total anti-oxidant capacity of cell culture media. Clin. Exp. Pharmacol. Physiol. 2007, 34, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Wu, Z.; Dai, Z.; Yang, Y.; Wang, W.; Liu, C.; Wang, B.; Wang, J.; Yin, Y. Dietary requirements of “nutritionally non-essential amino acids” by animals and humans. Amino Acids 2013, 44, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Zinellu, A.; Sotgia, S.; Scanu, B.; Chessa, R.; Gaspa, L.; Franconi, F.; Deiana, L.; Carru, C. Taurine determination by capillary electrophoresis with laser-induced fluorescence detection: From clinical field to quality food applications. Amino Acids 2009, 36, 35–41. [Google Scholar] [CrossRef]
- Sotgia, S.; Arru, D.; Sotgiu, E.; Mangoni, A.A.; Forteschi, M.; Pintus, G.; Carru, C.; Zinellu, A. Simultaneous determination of the main amino thiol and thione in human whole blood by CE and LC. Bioanalysis 2016, 8, 945–951. [Google Scholar] [CrossRef]
- Benitez, R.; Fernandez, A.; Isabel, B.; Nunez, Y.; de Mercado, E.; Gomez-Izquierdo, E.; Garcia-Casco, J.; Lopez-Bote, C.; Ovilo, C. Modulatory Effects of Breed, Feeding Status, and Diet on Adipogenic, Lipogenic, and Lipolytic Gene Expression in Growing Iberian and Duroc Pigs. Int. J. Mol. Sci. 2017, 19, 22. [Google Scholar] [CrossRef]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]
- Steibel, J.P.; Poletto, R.; Coussens, P.M.; Rosa, G.J. A powerful and flexible linear mixed model framework for the analysis of relative quantification RT-PCR data. Genomics 2009, 94, 146–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maternal Features | HTX | C |
|---|---|---|
| Body-Weight (Kg) | 94.7 ± 3.8 | 98.6 ± 6.9 |
| Back-fat depth (cm) | 1.9 ± 0.1 | 2.2 ± 0.3 |
| TEAC (mmol/L) | 5.6 ± 0.1 | 5.6 ± 0.1 |
| Glutathione (µmol/L) | 383.4 ± 49.8 | 388.3 ± 108.9 |
| Glycine (µmol/L) | 805.5 ± 25.5 | 757.1 ± 34.7 |
| Taurine (µmol/L) | 106.2 ± 23.1 | 83.3 ± 9.4 |
| Serine (µmol/L) | 154.2 ± 19.6 | 140.5 ± 21.6 |
| Glucose (mg/dL) | 97.5 ± 3.9 | 96.8 ± 3.7 |
| Fructosamine (mg/dL) | 321.7 ± 22.4 | 308.1 ± 15.5 |
| Triglycerides (mg/dL) | 35.3 ± 3.6 | 33.1 ± 2.9 |
| Total cholesterol (mg/dL) | 75.7 ± 7.5 | 66.6 ± 5.0 |
| HDL-cholesterol (mg/dL) | 28.4 ± 1.9 | 25.6 ± 1.1 |
| LDL-cholesterol (mg/dL) | 38.5 ± 4.6 | 33.2 ± 2.9 |
| Parameter | HTX | C | ||||
|---|---|---|---|---|---|---|
| Mean | Female | Male | Mean | Female | Male | |
| TEAC (mmol/L) | 2.2 ± 0.05 g | 2.1 ± 0.07 | 2.3 ± 0.08 | 2.1 ± 0.04 h | 2.2 ± 0.06 | 2.1 ± 0.07 |
| Glutathione (µmol/L) | 390.2 ± 15.9 a | 389.7 ± 19.1 | 395.7 ± 23.5 | 354.7 ± 17.9 a | 374.6 ± 25.6 | 338.4 ± 38.7 |
| Glycine (µmol/L) | 691.9 ± 32.1 c | 718.3 ± 14.1 | 661.3 ± 17.4 | 770.5 ± 26.6 d | 797.3 ± 50.8 | 730.4 ± 62.3 |
| Serine (µmol/L) | 262.9 ± 8.1 | 281.4 ± 10.8 | 237.9 ± 9.5 | 291.1 ± 17.7 | 306.1 ± 28.7 | 271.7 ± 16.4 |
| Taurine (µmol/L) | 304.1 ± 21.5 | 311.7 ± 19.3 | 286.9 ± 23.8 | 320.9 ± 17.8 | 314.1 ± 31.7 | 320.6 ± 38.2 |
| Parameter | HTX | C | ||||
|---|---|---|---|---|---|---|
| Mean | Female | Male | Mean | Female | Male | |
| CRL | 21.3 ± 0.2 | 21.3 ± 0.3 | 21.6 ± 0.3 | 21.4 ± 0.2 | 21.2 ± 0.4 | 21.8 ± 0.4 |
| ONL | 11.8 ± 0.1 | 11.7 ± 0.1 | 11.6 ± 0.2 | 11.4 ± 0.1 | 11.4 ± 0.1 | 11.5 ± 0.2 |
| BPD | 4.1 ± 0.1 g | 4.1 ± 0.1 | 4.0 ± 0.1 | 4.2 ± 0.1 h | 4.2 ± 0.1 | 4.3 ± 0.1 |
| TC | 17.1 ± 0.2 | 17.1 ± 0.2 | 16.9 ± 0.4 | 17.5 ± 0.2 | 17.2 ± 0.4 | 18.2 ± 0.3 |
| AC | 14.1 ± 0.3 | 14.1 ± 0.3 | 13.9 ± 0.4 | 14.3 ± 0.2 | 14.2 ± 0.4 | 14.4 ± 0.3 |
| Body-weight | 668.3 ± 18.4 e | 656.1 ± 21.0 | 667.4 ± 30.3 | 736.3 ± 15.5 f | 702.2 ± 24.3 | 765.3 ± 26.9 |
| Viscerae | 102.5 ± 2.8 c | 104.5 ± 3.7 | 99.8 ± 4.5 | 114.5 ± 3.7 d | 113.1 ± 4.5 | 116.3 ± 6.2 |
| Brain | 25.4 ± 0.6 | 25.8 ± 0.3 | 24.8 ± 1.5 | 25.4 ± 0.3 | 25.4 ± 0.4 | 25.4 ± 0.4 |
| Heart | 6.4 ± 0.2 | 6.3 ± 0.3 | 6.5 ± 0.3 | 6.7 ± 0.2 | 6.5 ± 0.3 | 7.0 ± 0.3 |
| Lungs | 22.6 ± 0.8 c | 22.3 ± 0.8 | 22.9 ± 1.5 | 24.9 ± 0.8 d | 23.7 ± 1.0 | 26.4 ± 1.0 |
| Liver | 19.2 ± 0.6 c | 19.3 ± 0.7 | 18.9 ± 1.1 | 20.9 ± 0.6 d | 20.2 ± 0.8 | 21.8 ± 0.8 |
| Kidneys | 6.2 ± 0.2 | 6.2 ± 0.2 | 6.1 ± 0.3 | 6.5 ± 0.2 | 6.5 ± 0.3 | 6.7 ± 0.3 |
| Intestine | 27.7 ± 0.9 g | 27.9 ± 1.2 | 27.3 ± 1.6 | 32.0 ± 1.1 h | 31.3 ± 1.7 | 32.9 ± 1.3 |
| Pancreas | 0.8 ± 0.01 c | 0.8 ± 0.01 | 0.7 ± 0.01 | 0.9 ± 0.01 d | 0.9 ± 0.1 | 0.9 ± 0.1 |
| Spleen | 1.3 ± 0.1 | 1.2 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.07 | 1.3 ± 0.1 | 1.4 ± 0.1 |
| Adrenal | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 | 0.1 ± 0.01 |
| Placenta | 249.3 ± 10.3 | 237.6 ± 10.7 | 265.4 ± 19.1 | 271.8 ± 8.9 | 265.3 ± 11.5 | 280.2 ± 14.0 |
| Parameter | HTX | C | ||||
|---|---|---|---|---|---|---|
| Mean | Female | Male | Mean | Female | Male | |
| F2 | 30.1 ± 0.7 | 29.9 ± 1.0 | 31.8 ± 0.8 | 29.7 ± 0.8 | 32.0 ± 0.9 | 25.9 ± 1.4 |
| Area | 9.8 ± 0.3 | 9.6 ± 0.5 | 10.0 ± 0.4 | 10.4 ± 0.5 | 10.9 ± 0.6 | 7.9 ± 0.4 |
| Parameter | HTX | C | ||||
|---|---|---|---|---|---|---|
| Mean | Female | Male | Mean | Female | Male | |
| Glucose (mg/dL) | 123.6 ± 14.0 c | 133.2 ± 20.2 | 110.5 ± 18.4 | 177.6 ± 19.8 d | 200.2 ± 27.8 | 148.2 ± 27.0 |
| Fructosamine (mg/dL) | 187.2 ± 3.5 e | 183.6 ± 4.5 | 192.1 ± 5.4 | 167.5 ± 6.0 f | 174.5 ± 6.8 | 158.3 ± 10.5 |
| Triglycerides (mg/dL) | 47.6 ± 1.4 | 47.6 ± 1.8 | 47.5 ± 2.1 | 52.5 ± 2.6 | 53.6 ± 3.4 | 50.9 ± 3.9 |
| Total Cholesterol (mg/dL) | 60.3 ± 1.7 | 59.7 ± 2.2 | 61.0 ± 2.9 | 59.6 ± 2.0 | 59.6 ± 2.4 | 59.7 ± 3.4 |
| HDL-c (mg/dL) | 20.9 ± 0.5 | 21.1 ± 0.6 | 20.7 ± 0.8 | 22.6 ± 2.8 | 20.2 ± 0.6 | 25.6 ± 6.4 |
| LDL-c (mg/dL) | 28.8 ± 1.1 | 28.5 ± 1.5 | 29.2 ± 1.8 | 31.3 ± 1.1 | 30.8 ± 1.5 | 32.0 ± 1.7 |
| Gene Symbol | Gene Name | Reference Sequence ID | Primer Sequences | Amplicon Length (bp) | Efficiency (%) |
|---|---|---|---|---|---|
| CAT | Catalase | NM_214301.2 | TGGCCCCATGTGCTTTCA GGCGTTTCCTCTCCTCCTCAT | 209 | 83.75 |
| SOD1 | Superoxide dismutase 1 | NM_001190422 | GTGACTGCTGGCAAAGATGGTGTG TTTCCCGTCTTTGTACTTTCTTCA | 158 | 89.75 |
| HIF1A | Hypoxia inducible factor 1 subunit alpha | NM_001123124.1 | GGCGGCGCGAACGACAAGA CACACGCAAATAGCTGATGGTAAG | 195 | 93.8 |
| NOS2 | Nitric oxide synthase 2 | XM_013981169.2 | TGGGGCAGCGGGATGACTTTC CACCCTGGCCAGATGTTCCTC | 231 | 77.05 |
| VEFGA | Vascular endothelial growth factor A | NM_214084.1 | CATCTTCAAGCCGTCCTGTGT ATTTTCTTGCCTCGCTCTATCTTT | 211 | 92.1 |
| IGF1 | Insulin like growth factor 1 | XM_005664199.3 | TGCGGAGACAGGGGCTTTTATTTC CCTTGGGCATGTCCGTGTGG | 199 | 88.1 |
| UCP2 | Uncoupling protein 2 | ENSSSCG00000014833 | CCCTGCGGCCCGGACACATAG GACGCCTCCACTCAGCAGCAAGAC | 217 | 86.25 |
| ACTB | Beta-actin | XM_003124280.4 | TCTGGCACCACACCTTCT GATCTGGGTCATCTTCTCAC | 114 | 89.35 |
| B2M | Beta-2-microglobulin | NM_213978.1 | TTCACACCGCTCCAGTAG CCAGATACATAGCAGTTCAGG | 166 | 83 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Contreras, C.; Vazquez-Gomez, M.; Barbero, A.; Pesantez, J.L.; Zinellu, A.; Berlinguer, F.; Gonzalez-Añover, P.; Gonzalez, J.; Encinas, T.; Torres-Rovira, L.; et al. Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Placental Gene Expression and Fetal Antioxidant Status, DNA-Methylation and Phenotype. Int. J. Mol. Sci. 2019, 20, 1187. https://doi.org/10.3390/ijms20051187
Garcia-Contreras C, Vazquez-Gomez M, Barbero A, Pesantez JL, Zinellu A, Berlinguer F, Gonzalez-Añover P, Gonzalez J, Encinas T, Torres-Rovira L, et al. Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Placental Gene Expression and Fetal Antioxidant Status, DNA-Methylation and Phenotype. International Journal of Molecular Sciences. 2019; 20(5):1187. https://doi.org/10.3390/ijms20051187
Chicago/Turabian StyleGarcia-Contreras, Consolación, Marta Vazquez-Gomez, Alicia Barbero, José Luis Pesantez, Angelo Zinellu, Fiammetta Berlinguer, Pedro Gonzalez-Añover, Jorge Gonzalez, Teresa Encinas, Laura Torres-Rovira, and et al. 2019. "Polyphenols and IUGR Pregnancies: Effects of Maternal Hydroxytyrosol Supplementation on Placental Gene Expression and Fetal Antioxidant Status, DNA-Methylation and Phenotype" International Journal of Molecular Sciences 20, no. 5: 1187. https://doi.org/10.3390/ijms20051187