Evolution and Functional Analysis of orf1 Within nif Gene Cluster from Paenibacillus graminis RSA19

State Key Laboratory for Agrobiotechnology, Key Laboratory of Soil Microbiology of Agriculture Ministry and College of Biological Sciences, China Agricultural University, Beijing 100193, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(5), 1145; https://doi.org/10.3390/ijms20051145
Submission received: 6 January 2019
/
Revised: 27 February 2019
/
Accepted: 1 March 2019
/
Published: 6 March 2019
(This article belongs to the Section Molecular Microbiology)
Abstract
:Paenibacillus is a genus of Gram-positive, facultative anaerobic and endospore-forming bacteria. Genomic sequence analysis has revealed that a compact nif (nitrogen fixation) gene cluster comprising 9–10 genes nifBHDKENX(orf1)hesAnifV is conserved in diazotrophic Paenibacillus species. The evolution and function of the orf1 gene within the nif gene cluster of Paenibacillus species is unknown. In this study, a careful comparison analysis of the compositions of the nif gene clusters from various diazotrophs revealed that orf1 located downstream of nifENX was identified in anaerobic Clostridium ultunense, the facultative anaerobic Paenibacillus species and aerobic diazotrophs (e.g., Azotobacter vinelandii and Azospirillum brasilense). The predicted amino acid sequences encoded by the orf1 gene, part of the nif gene cluster nifBHDKENXorf1hesAnifV in Paenibacillus graminis RSA19, showed 60–90% identity with those of the orf1 genes located downstream of nifENX from different diazotrophic Paenibacillus species, but shared no significant identity with those of the orf1 genes from different taxa of diazotrophic organisms. Transcriptional analysis showed that the orf1 gene was expressed under nitrogen fixation conditions from the promoter located upstream from nifB. Mutational analysis suggested that the orf1 gene functions in nitrogen fixation in the presence of a high concentration of O2.
1. Introduction
Biological nitrogen fixation, the conversion of atmospheric N2 to NH3, plays an important role in the global nitrogen cycle and in world agriculture. The ability to fix nitrogen is widely, but sporadically distributed among Archaea and Bacteria which includes these families: Proteobacteria, Firmicutes, Cyanobacteria, Actinobacteria, Chloroflexi and Chlorobi [1]. Most biological nitrogen fixation is catalyzed by a molybdenum-dependent nitrogenase, which consists of two component proteins: Fe protein and MoFe protein. The MoFe protein component is an α2β2 heterotetramer (encoded by nifD and nifK) that contains two metalloclusters: FeMo-co, a [Mo-7Fe-9S-C-homocitrate] cluster, which serves as the active site of substrate binding and reduction and the P-cluster, a [8Fe-7S] cluster which shuttles electrons to FeMo-co. The Fe protein (encoded by nifH) is a γ2 homodimer bridged by an intersubunit (4Fe-4S) cluster that serves as the obligate electron donor to the MoFe protein [2,3].
Nitrogenase emerged in anaerobes and later diversified into facultative anaerobes and aerobes. The transition of nitrogenase from anaerobic to facultative anaerobic and aerobic organisms was accompanied by a substantial increase in the number of nif genes from a minimum of 7 to a maximum of 20 genes [4]. For example, a minimum of 7 nif genes (nifHDKEBI1I2) were identified in the mesophilic archaeon Methanocaldococcus sp. strain FS406-22, a nif gene cluster composed of 9–10 genes (nifBHDKENX (orf1) hesAnifV) was conserved in facultative Paenibacillus species, and 20 nif genes (nifJHDKTYENXUSVWZMFLABQ) were found in Klebsiella oxytoca [5]. The obligate aerobes Azotobacter vinelandii contains the most complex nif gene cluster composed of nifHDKTYENXUSVZMFLABQ as well as nafY, iscA and other function-unknown genes, which colocalize with the nif genes [4]. A careful comparison analysis of the repertoire of nif genes in known diazotrophic species demonstrates that a minimum set of six genes nifHDKENB coding for structural and biosynthetic components were present [1]. Genetic and biochemical studies on the two model diazotrophs K. oxytoca and A. vinelandii revealed that nifH, nifD and nifK genes encode the structural subunits, the nifE, nifN, nifX, nifB, nifQ, nifV, nifY, nifS, nifU and nifH contribute to the synthesis and insertion of FeMo-co into nitrogenase, and nifL and nifA are involved in the regulation of other nif gene transcription [3]. In archaea and some Gram-positive anaerobes, NifI1 and NifI2, the GlnB homologues, regulate nitrogenase activities at a post-transcriptional level by interacting with nifHDK in response to N availability [6].
Paenibacillus is a large genus of Gram-positive, facultative anaerobic, endospore-forming bacteria. The facultative anaerobic Paenibacillus is in the middle stage of evolutionary history from anaerobes to aerobes. The genus Paenibacillus currently comprises more than 150 named species, more than 20 of which have a nitrogen fixation ability [7,8]. Our comparative genomic analysis showed that diazotrophic Paenibacillus species fell into two distinct sub-groups (Sub-group I and Sub-group II). The N2-fixing strains (e.g., Paenibacillus polymyxa WLY78) within Sub-group I had a nif gene cluster consisting of nine genes (nifB, nifH, nifD, nifK, nifE, nifN, nifX, hesA and nifV) within a 10 kb region, while the N2-fixing strains (e.g., Paenibacillus graminis RSA19) within Sub-group II have a nif gene cluster composed of 10 genes (nifB, nifH, nifD, nifK, nifE, nifN, nifX, orf1, hesA and nifV) within a 11 kb region. The orf1 located between nifX and hesA is the major pronounced difference in the nif cluster between Sub-group I and Sub-group II [8]. Each of the two nif gene clusters is organized as a single operon and transcribed under the σ70 promoter located in front of nifB [9,10]. Recently, our studies have revealed that GlnR simultaneously acts as an activator and a repressor for nif gene transcription by binding to two different loci of the single nif promoter region according to nitrogen availability [11]. The orf1 located downstream of nifENX was also found in some other diazotrophic bacteria, such as Azospirillum brasilense Sp7 and Rhodobacter capsulatus [12,13]. Although orf1 gene behind nifENX was often conserved in a lot of nif gene clusters of diazotrophs, its evolution and function were not fully clarified.
In this study, we aimed to investigate the evolution of orf1 located downstream of nifENX during the evolutionary history of nitrogenase from anaerobes to aerobes, and determine the function of orf1 in nitrogen fixation. We selected P. graminis RSA19, a gram-positive, facultative anaerobic and endospore-forming bacterium isolated from the rhizosphere of maize, for this study. Our results revealed that orf1 located downstream of nifENX was originally found in the nif gene cluster of Clostridium ultunense and some Paenibacillus species/strains of Firmicutes and then perhaps it was transferred to other aerobes. The expression and transcription of the orf1 gene were determined by qRT-PCR and RT-PCR. Mutation analysis showed a much lower nitrogenase activity in the Δorf1 mutant than in the wild-type strain, which was observed under high oxygen. These results indicated that orf1 plays a role in the protection of the nitrogenase against inactivation by O2.
2. Results and Discussion
2.1. Acquisition and Inheritance of Paenibacillus orf1 Gene
The orf1 gene is a part of the nif gene cluster (nifB, nifH, nifD, nifK, nifE, nifN, nifX, orf1, hesA and nifV) in P. graminis RSA19 and some other Paenibacillus species or strains. To investigate the evolution of orf1, we here performed a comparison of the nif gene clusters from the representatives of diazotrophic methanogenic archae and bacteria in six taxonomic phyla: Actinobacteria, Chlorobi, Chloroflexi, Cyanobacteria, Firmicutes and Proteobacteria.
As shown in Figure 1, the simplest nif gene organization is found in methanogenic archae. For examples, Methanocaldococcus sp. strain FS406-22, Methanobacterium thermoauttatrophicum, Methanococcus maripaludis and Methanosarcina mazei Gö1, carry a single nif gene cluster composed of six to eight nif genes (nifH, nifI1, nifI2, nifD, nifK, nifE, nifN and nifX) in a single operon [4,14,15,16]. The nifB gene, which is essentially required for nitrogenase, is located elsewhere on genomes outside of the nif gene cluster in these archae. The nifB nifH nifD nifK nifE nifN are responsible for the synthesis of nitrogenase, and the nifI1 nifI2 are involved in the regulation of nitrogenase activity in these diazotrophic archae [17,18].
The regulatory genes nifI1 and nifI2, which were common in methanogenic archae were also found in most of the anaerobic, Gram-positive bacteria. For example, in the strictly anaerobic, Gram-positive phototrophic bacterium Heliobacterium chlorum, the positions of nifI1 and nifI2 were in the front of nifH in the nif gene cluster composed of 11 genes in the order nifI1, nifI2, nifH, nifD, nifK, nifE, nifN, nifX, fdx, nifB and nifV [19,20]. Notably, nifB emerged as a part of the nif gene cluster in these anaerobic bacteria. A nif gene cluster composed of nifV nifA (orf1 orf2 orf3) nifH nifI1 nifI2 nifD nifK nifE nifN nifB was found in Chlorobium tepidum, which is a green sulphur bacterium [21].
Both Paenibacillus and Clostridium belong to Firmicutes. Clostridium is a genus of obligate anaerobic bacteria, while Paenibacillus is a genus of facultative anaerobic bacteria. The gene organization nifH-nifI1-nifI2-nifD-nifK-nifE-nifN, which was common in methanogenic archae was found in anaerobic Clostridium acetobutylicum and Clostridium beijerinckii [22]. For example, a nif gene cluster composed of nine genes (nifH nifI1 nifI2 nifD nifK nifE nifN-B nifVw nifVα) was found in C. acetobutylicum ATCC 824 [22].
A nif gene cluster consisting of nine genes within a 10 kb region in the order nifB, nifH, nifD, nifK, nifE, nifN, nifX, hesA and nifV was identified in diazotrophic Paenibacillus species. Importantly, an orf1 emerged downstream of nifENX within the nif gene cluster in some Paenibacillus species [8]. The regulation genes nifI1 and nifI2, which were conserved in anaerobic archae and bacteria were lost in the nif gene clusters of the facultative anaerobic Paenibacillus. In place of nifI1 and nifI2, a glnR which is not associated with the nif gene cluster regulated the transcription of the nif gene cluster in Paenibacillus [11]. Interestingly, from the genome sequence data of Clostridium ultunense strain Esp [23], we found that a nif gene cluster (nifB nifV orf1 nifH nifD nifK nifE nifN nifX orf1 hesA nifW nifZ), which was different from those of C. acetobutylicum and C. beijerinckii, emerged in this bacterium. The organization of nifH-nifD-nifK-nifE-nifN-nifX-orf1-hesA, which is conserved in diazotrophic Paenibacillus, was for the first time found in Clostridium. From the hypothesis that the transition of Nif from anoxic to oxic environments, the organization of nifH-nifD-nifK-nifE-nifN-nifX-orf1-hesA including orf1 in Paenibacillus should be evolved via horizontal gene transfer from C. ultunense strain Esp. However, compared to the 9 to 10 genes within the nif genes cluster of Paenibacillus, the numbers of the 13 genes within the nif genes cluster of C. ultunense were larger, and the 2 genes nifW and nifZ, which were found in aerobic diazotrophic bacteria, emerged in this bacterium. From the hypothesis that the evolution of Nif during the transition from anaerobic to aerobic metabolism was accompanied by a substantial increase in the number of nif genes, the organization of nifH-nifD-nifK-nifE-nifN-nifX-orf1-hesA including orf1 in C. ultunense strain Esp should be evolved via horizontal gene transfer from Paenibacillus.
As shown in Figure 1, the organization nifH-nifD-nifK-nifE-nifN-nifX-orf1, which is conserved in facultative anaerobic Paenibacillus, was also identified in Frankia (e.g., Frankia sp. EulK1 [24]). The nifHDKENXorf1 cluster was also identified, but was separated into two clusters: nifHDK and ENXorf1 in Cyanobacteria (e.g., Cyanothece sp. 51,142 [25]) and Proteobacteria: Rhodobacter capsulatus, Herbaspirillum seropedicae, Rhodopseudomonas palustris, Pseudomonas stutzeri and Azotobacter vinelandii, with NifA as a positive transcription regulator in these diazotrophs [26,27,28,29].
2.2. Identity and Phylogenetic Analysis of the Orf1
The orf1 gene of P. graminis RSA19 is 567 bp in length. The predicted protein of Orf1 has a molecular weight of 20.5 kDa and pI of 8.43. The deduced-amino acid sequence of the orf1 gene product in P. graminis RSA19 was aligned with the corresponding sequences from various known diazotrophs extracted from GenBank databases (Figure 2). The identities of Orf1 of P. graminis RSA19 with those from other Paenibacillus species at amino acid levels ranged between 60% and 90%. Except that P. graminis Orf1 showed the highest identities with those of other diazotrophic Paenibacillus spp., which showed the highest identity (32.89%) with that of C. ultunense. The P. graminis Orf1 showed 18–32% identities with those of Frankia sp. EuIK1, Herbaspirillum seropedicae, Cyanothece sp. ATCC 51142, Rhodopseudomonas palustris, Rhodobacter capsulatus and Azotobacter vinelandii (Table S1).
Structure analysis revealed that the predicated protein encoded by the orf1 located downstream of nifENX in cyanobacterium Cyanothece 51,142 had a conserved domain (DUF269) [30]. We found that the Orf1 of P. graminis RSA19 and those from other diazotrophs also had the conserved domain (DUF269). Amino acid residues within this domain were conserved, such as G124, G126, G131, F146, and F148 (Figure 2). The predicted 3-dimensional structure of Orf1 protein using SWISS-MODEL online software (https://swissmodel.expasy.org.) was identical to that of Cce_0566 from Cyanothece (Figure S1). The DUF269 domain, which is only found in the nitrogen-fixing species may play some role in nitrogen fixation.
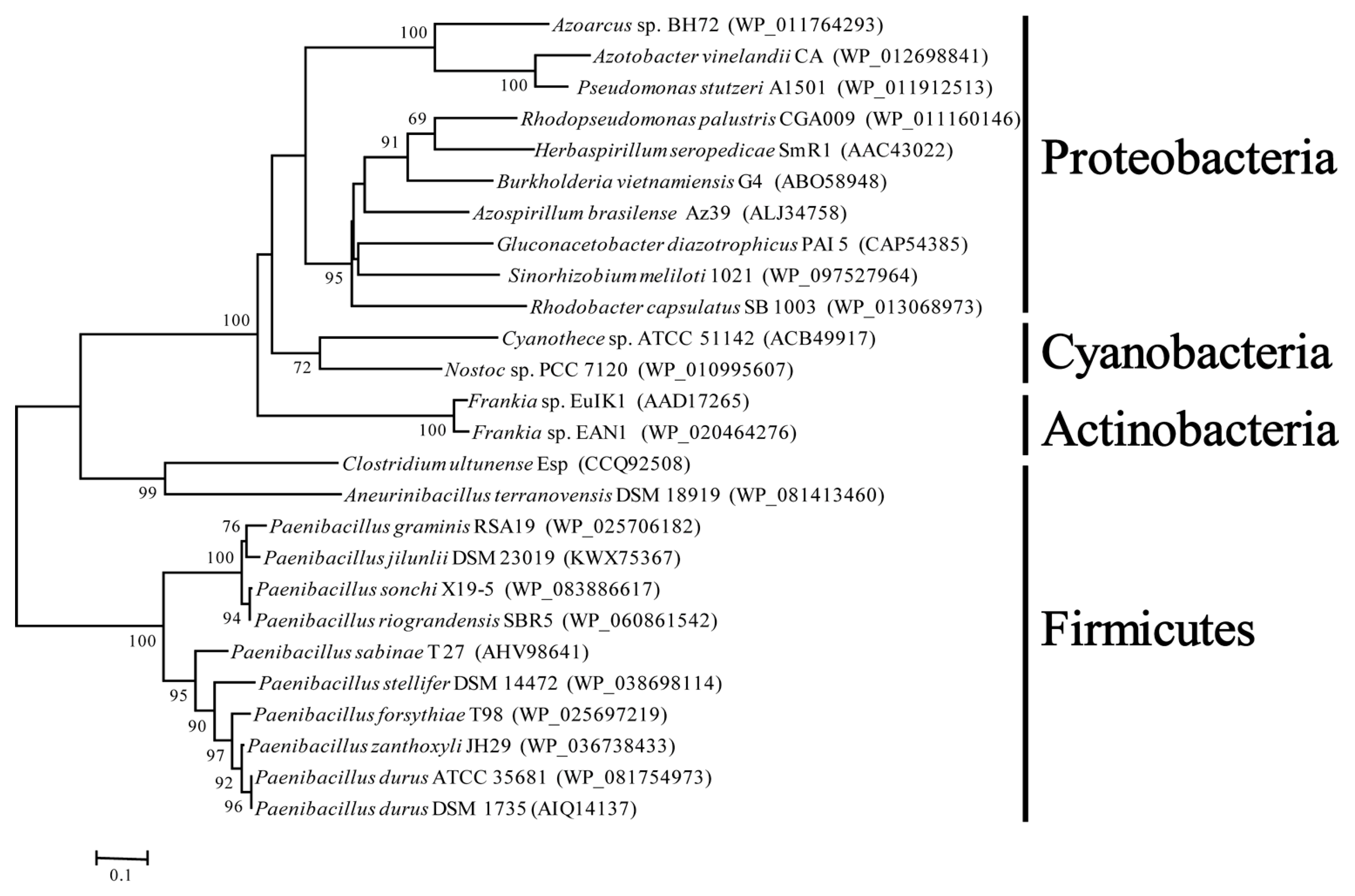
The phylogenetic analysis showed that orf1 genes downstream of nifENX from facultative anaerobic P. graminis RSA19 and other Paenibacillus species (P. sonchi, P. riograndensis, P. sabine and P. durus) formed a monophyletic group, and orf1 genes downstream of nifENX from aerobic bacteria, such as Frankia, Cyanobacteria and Proteobacteria, formed another group (Figure 3). The data were consistent with our previous results that Paenibacillus and Frankia were sister groups by the NifBHDKENX phylogenetic analysis [8]. The results suggested that the orf1 located downstream of nifENX was originally found in Paenibacillus species and C. ultunense, and then perhaps was transferred to anaerobic and aerobic bacteria, such as Cyanobacteria and Proteobacteria.
2.3. Transcriptional Analysis of the P. graminis orf1
In this study, sequence analysis and RT-PCR demonstrated that the 10 genes nifB nifH nifD nifK nifE nifN nifX orf1 hesA nifV in P. graminis RSA19 were organized as an operon under the control of a -35/-10 type σ70-dependent promoter located upstream of nifB (Figure 4). Similar results that the 10 genes nifB nifH nifD nifK nifE nifN nifX orf1 hesA nifV in P. sabinae T27 are organized as an operon were reported in [10]. In this study, qRT-PCR results showed that these 10 genes within the nif gene operon under N2-fixing condition (without NH4+ and O2) exhibited a significant transcript ranging from 60- to 1300-fold compared to non-N2-fixing condition (100 mM NH4+ and 21% O2) (Figure 5), suggesting that nif gene expression in P. graminis RSA19 was strongly regulated by ammonium and oxygen. We found that the transcripts of nifBHDK gene were much higher than those of nifENXorf1hesAnifV, although these genes were co-transcribed from a common promoter in front of nifB. The different abundances of these co-transcribed genes suggested that these transcripts have different processing and stabilities, similar results were also found in P. polymyxa WLY78 [31] and A. vinelandii [32]. Taken together, the results suggest that the orf1 gene is expressed under nitrogen fixation conditions from the promoter located upstream from nifB in P. graminis RSA19. Likewise, the orf1 expression occurred only under nitrogen-fixing conditions in Frankia and Cyanothece 51,142 [33,34,35].
2.4. Mutation and Functional Analysis of the orf1 Gene
To elucidate the function of orf1 in nitrogen fixation of P. graminis RSA19, we constructed an in-frame deletion mutant Δorf1, a complementary strain (Δorf1/orf1) and an overexpression strain (WT/orf1) as described in the materials and methods section. In comparison with wild-type P. graminis RSA19, which exhibits the highest nitrogenase activity under anaerobic and microaerobic conditions and no activity in the presence of more than 12% O2, activities in Δorf1 mutant were a little lower compared to the wild-type strain under low (0–1%) oxygen conditions (Figure 6). Especially at high concentrations of oxygen, a much lower nitrogenase activity in the Δorf1 mutant than in the wild-type strain was observed. Complementation and overexpression of orf1 by introduction of orf1 carried on multicopy vector pHY300PLK into Δorf1 mutant and wild-type strain led to the enhancement of activity in the presence of high oxygen. These results suggest that orf1 is beneficial for nitrogen fixation at high oxygen concentrations. Furthermore, we found that Δorf1 mutant grew equally well both in the absence and presence of high oxygen concentrations. These results for the first time indicate physiological involvement of orf1 gene products in the protection of the biosynthetic pathway or protection of the nitrogenase under high oxygen conditions.
It was previously reported that insertional mutation of the orf1 gene of H. seropedicae had no effect on nitrogenase activity under a normal nitrogen-limited condition [36], but resulted in a significant decrease in nitrogenase activity under low iron levels [37]. In this study, we found that both the Δorf1 mutant and wild-type strain exhibited a significant reduction in nitrogenase activity under iron limitation compared to an iron sufficiency condition (Figure 7). Our results showed that orf1 gene was not closely related to nitrogenase activity mediated by iron.
A recurrent characteristic of the location of orf1 in many diazotrophs is the presence in their neighborhood, usually in the same operon, of nifENX genes involved in the iron-molybdenum cofactor biosynthesis, suggesting its possible participation in FeMoco biosynthesis. Functional analysis of orf1 gene located downstream of nifENX was also performed in several diazotrophs, but obvious functions of orf1 gene in nitrogen fixation are still lacking. For example, mutation of a ferredoxin-like gene of R. capsulatus, which is co-transcribed with the nifENX genes did not abolish its nitrogenase activity [12], but this ferredoxin-like protein may be used as an alternative electron donor or source of iron-sulfur centers. The fact that deletion of orf1 resulted in no loss of activity under normal nitrogen fixation conditions (anaerobic or low oxygen and without ammonium) was also observed in A. vinelandii, H. seropedicae and R. capsulatus [12,36,38], which was in agreement with our results. Our results suggested that orf1 gene in P. graminis RSA19 was required for nitrogen fixation under high oxygen conditions. How the orf1 affects nitrogenase activity in the presence of oxygen needs to be determined in the future. Since nitrogenase is sensitive to oxygen, a variety of strategies are used to provide protection from oxygen, including consumption of excess oxygen by respiration and regulation of nif gene expression [39].
3. Materials and Methods
3.1. Strains, Plasmids and Growth Conditions
Bacterial strains and plasmids used in this study are summarized in Table 1. P. graminis [40] and derivatives were routinely grown in an LD medium (per liter contained: 2.5 g NaCl, 5 g yeast and 10 g tryptone) at 30 °C with shaking. For assays of nitrogenase activity and nif expression, P. graminis RSA19 strains were grown in nitrogen-limited media under anaerobic conditions. For measuring the effect of oxygen on nitrogenase activity, P. graminis and derative strains were cultivated at different concentrations of O2 in nitrogen-limited media. Nitrogen-limited medium contained (per liter): 10.4 g Na2HPO4, 3.4 g KH2PO4, 26 mg CaCl2•2H2O, 30 mg MgSO4, 0.3 mg MnSO4, 36 mg Ferric citrate, 7.6 mg Na2MoO4•2H2O, 10 mg p-aminobenzoic acid, 5 µg biotin, 2 mM glutamate and 4 g glucose as a carbon source. Iron-free medium was prepared by omitting ferric citrate from the nitrogen-limited medium.
Escherichia coli strains JM109 was used for regular plasmid extraction, and the Dam− Dcm−strain Trans110 was used to produce unmethylated plasmid DNA. Thermo-sensitive vector pRN5101 [41] was used for gene disruption in P. graminis RSA19. Shuttle vector pHY300PLK was used for complementation and overexpression experiment. When appropriate, antibiotics were added in the following concentrations: 100 μg/mL ampicillin, 12.5 μg/mL tetracycline and 5 μg/mL erythromycin for maintenance of plasmids.
3.2. Molecular Techniques
Plasmid and genomic DNA extraction, gel electrophoresis, restriction mapping, transformation and molecular cloning, amplification by PCR were performed by standard methods [42] or as recommended by the instructions on the products.
3.3. Construction of orf1 Deletion Mutant
Deletion of orf1 was performed by a homologous recombination method. Amplification of a 931-bp DNA fragment located upstream of orf1 using the primer set orf1UF and orf1UR and a 912-bp DNA fragment located downstream of orf1 using the set orf1DF and orf1DR was performed (Table 2). The two fragments flanking coding region of Δorf1 were then fused with Hind III/BamH Ι digested pRN5101 vector using Gibson assembly master mix (New England Biolabs, Ipswich, USA), generating a recombinant plasmid pRDorf1. The recombinant plasmid pRDorf1 was electroporated into P. graminis RSA19. Then, the single-crossover transformants were selected for erythromycin resistance (Emr). Subsequently, the double-crossover transformants were selected from the initial erythromycin transformants after several rounds of nonselective growth at 39 °C and confirmed by PCR amplification. Correct recombination was checked using primers orf1UF and orf1DR (Table 2), followed by nucleotide sequencing of the amplicon obtained.
Complementation and overexpression of orf1 by the introduction of orf1 under nifB promoter of nif cluster operon carried on multicopy vector pHY300PLK into orf1 deletion mutant and wild-type strain. A 1147 bp DNA fragment containing the coding region of orf1 and a 576 bp promoter region of nif operon were PCR amplified, and then the two fragments were fused together using Gibson assembly master mix. These fragments were digested with BamH I/Hind III, and ligated into vector pHY300PLK, generating vector pHYorf1. The recombinant plasmid was transformed into Δorf1 mutant and P. graminis RSA19, and tetracycline-resistant (Tcr) transformants were selected and confirmed by PCR and sequencing. The primers used here are listed in Table 2. Plasmids were transformed to P. graminis RSA19 by electroporation with the methods described by Zhang et al. in [43] and Bach et al. in [44] with modifications.
3.4. RT-PCR and qRT-PCR Analysis
For RT-PCR, P. graminis RSA19 was grown in N2-fixing conditions (without NH4Cl and O2). For qRT-PCR, P. graminis RSA19 and Δorf1 mutant strain were grown in N2-fixing conditions (without NH4Cl and O2) and non-N2-fixing conditions (100 mM ammonium and 21% O2). The integrity and size distribution of the RNA was verified by agarose gel electrophoresis, and the concentration was determined spectrophotometrically. Synthesis of cDNA was carried out using RT Prime Mix according to the manufacturer’s specifications (Takara Bio, Tokyo, Japan). The nif gene transcripts were detected by using an RT-PCR Kit with 16S rDNA as a control. Primers for nif genes and 16S rDNA used for PCR are listed in Table 2. qRT-PCR was performed on Applied Biosystems 7500 Real-Time System and detected by the SYBR Green detection system with the following program: 95 °C for 15 min, 1 cycle; 95 °C for 10 s and 65 °C for 30 s, 40 cycles. The relative expression level was calculated using the 2−ΔΔCT method [45], where ΔΔCT = (CT gene of nif − CT gene of 16S rRNA) N2-fixing condition − (CT gene of nif − CT gene of 16S rRNA) non-N2-fixing condition The CT value is the cycle threshold at which the detected fluorescence crosses an arbitrarily placed threshold. Each experiment was performed in triplicate.
3.5. Acetylene Reduction Assays of Nitrogenase Activity
Acetylene reduction assays were performed as described previously to measure nitrogenase activity [9]. P. graminis RSA19 and its mutant strains were grown overnight in LD medium. The cultures were collected by centrifugation, washed three times with sterilized water and then resuspended in nitrogen-limited medium containing 2 mM glutamate as a nitrogen source to a final OD600 of 0.2–0.4. Then, 4 ml of the culture was transferred to a 25-mL test tube and the test tube was sealed with a rubber stopper. The headspace (21 mL) in the tube was then evacuated and replaced with argon gas. Then, C2H2 (10% of the headspace volume) was injected into the test tubes. For measuring the effect of oxygen on nitrogenase activity, O2 (0–12% of the headspace volume) was added at the same time. Cultures were incubated at 30 °C. C2H4 production was analyzed by gas chromatography. The nitrogenase activity was expressed as nmol C2H4/mg protein/hr. All treatments were in three replicates and all the experiments were repeated three or more times. The results of the nitrogenase activity were analyzed by analysis of the one-way analysis of variance (ANOVA) using SPSS software version 20 (SPSS Inc., Chicago, IL, USA). Means of different strains were compared using the least significant difference (LSD) at a 0.05 or 0.01 level of probability.
4. Conclusions
P. graminis RSA19 has an orf1 gene within the nif gene cluster (nifB nifH nifD nifK nifE nifN nifX orf1 hesA nifV). The orf1 located downstream of nifENX was also identified in some other facultative anaerobic Paenibacillus species, anaerobic Clostridium ultunense and aerobic diazotrophs (e.g., Azotobacter vinelandii and Azospirillum brasilense). The orf1 emerged in anaerobes and facultative anaerobes and later recruited into aerobes during the evolution process. The orf1 gene may play a role in protection of the nitrogenase against oxygen.
Supplementary Materials
Supplementary Materials can be found at https://www.mdpi.com/1422-0067/20/5/1145/s1.
Author Contributions
S.C. conceived and designed the experiments; Q.L. performed the experiments; Q.L., H.Z. and X.L. analyzed the data; Q.L. and S.C. wrote the paper.
Founding
This work was supported by the National Nature Science Foundation of China (Grant No. 31770083) and the Innovative Project of SKLAB (Grant No. 2017SKLAB1-1).
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Dos Santos, P.C.; Fang, Z.; Mason, S.W.; Setubal, J.C.; Dixon, R. Distribution of nitrogen fixation and nitrogenase-like sequences amongst microbial genomes. BMC Genom. 2012, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Ribbe, M.W. Biosynthesis of the metalloclusters of molybdenum nitrogenase. Microbiol. Mol. Biol. Rev. 2011, 75, 664–677. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.M.; Ludden, P.W. Biosynthesis of the iron-molybdenum cofactor of nitrogenase. Annu. Rev. Microbiol. 2008, 62, 93–111. [Google Scholar] [CrossRef] [PubMed]
- Boyd, E.S.; Costas, A.M.G.; Hamilton, T.L.; Mus, F.; Peters, J.W. Evolution of molybdenum nitrogenase during the transition from anaerobic to aerobic metabolism. J. Bacteriol. 2015, 197, 1690–1699. [Google Scholar] [CrossRef] [PubMed]
- Arnold, W.; Rump, A.; Klipp, W.; Priefer, U.B.; Puhler, A. Nucleotide sequence of a 24,206-base-pair DNA fragment carrying the entire nitrogen fixation gene cluster of Klebsiella pneumoniae. J. Mol. Biol. 1988, 203, 715–738. [Google Scholar] [CrossRef]
- Leigh, J.A. Nitrogen fixation in methanogens: The archaeal perspective. Curr. Issues Mol. Biol. 2000, 2, 125–131. [Google Scholar] [PubMed]
- Jin, H.J.; Lv, J.; Chen, S.F. Paenibacillus sophorae sp. nov., a nitrogen-fixing species isolated from the rhizosphere of Sophora japonica. Int. J. Syst. Evol. Microbiol. 2011, 61, 767–771. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.B.; Du, Z.L.; Bai, L.Q.; Tian, C.F.; Zhang, Y.Z.; Xie, J.Y.; Wang, T.S.; Liu, X.M.; Chen, X.; Cheng, Q.; et al. Comparative genomic analysis of N2-fixing and non-N2-fixing Paenibacillus spp.: Organization, evolution and expression of the nitrogen fixation genes. PLoS Genet. 2014, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Zhang, L.H.; Liu, Z.Z.; Zhao, D.H.; Liu, X.M.; Zhang, B.; Xie, J.B.; Hong, Y.Y.; Li, P.F.; Chen, S.F.; et al. A minimal nitrogen fixation gene cluster from Paenibacillus sp. WLY78 enables expression of active nitrogenase in Escherichia coli. PLoS Genet. 2013, 9, 11. [Google Scholar] [CrossRef]
- Li, X.X.; Deng, Z.P.; Liu, Z.Z.; Yan, Y.L.; Wang, T.S.; Xie, J.B.; Lin, M.; Cheng, Q.; Chen, S.F. The genome of Paenibacillus sabinae T27 provides insight into evolution, organization and functional elucidation of nif and nif-like genes. BMC Genom. 2014, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.S.; Zhao, X.Y.; Shi, H.W.; Sun, L.; Li, Y.B.; Li, Q.; Zhang, H.W.; Chen, S.F.; Li, J.L. Positive and negative regulation of transferred nif genes mediated by indigenous GlnR in Gram-positive Paenibacillus polymyxa. PLoS Genet. 2018, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Morenovivian, C.; Hennecke, S.; Puhler, A.; Klipp, W. Open reading frame-5 (ORF5), encoding a ferredoxin-like protein, and nifQ are cotranscribed with nifE,nifN,nifX,and ORF4 in Rhodobacter capsulatus. J. Bacteriol. 1989, 171, 2591–2598. [Google Scholar] [CrossRef]
- Potrich, D.P.; Bressel, T.A.; Schrank, I.S.; Passaglia, L.M.P. Sequencing and promoter analysis of the nifENXorf3orf5fdxAnifQ operon from Azospirillum brasilense Sp7. Braz. J. Med Biol. Res. 2001, 34, 1379–1395. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, C.; Veit, K.; Gottschalk, G.; Schmitz, R.A. Functional organization of a single nif cluster in the mesophilic archaeon Methanosarcina mazei strain Gö1. Archaea 2002, 1, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kessler, P.S.; Blank, C.; Leigh, J.A. The nif gene operon of the methanogenic archaeon Methanococcus maripaludis. J. Bacteriol. 1998, 180, 1504–1511. [Google Scholar] [PubMed]
- Smith, D.R.; DoucetteStamm, L.A.; Deloughery, C.; Lee, H.M.; Dubois, J.; Aldredge, T.; Bashirzadeh, R.; Blakely, D.; Cook, R.; Gilbert, K.; et al. Complete genome sequence of Methanobacterium thermoautotrophicum Delta H: Functional analysis and comparative genomics. J. Bacteriol. 1997, 179, 7135–7155. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.A.; Dodsworth, J.A. Nitrogen regulation in bacteria and archaea. Annu. Rev. Microbiol. 2007, 61, 349–377. [Google Scholar] [CrossRef] [PubMed]
- Lie, T.J.; Wood, G.E.; Leigh, J.A. Regulation of nif expression in Methanococcus maripaludis—Roles of the euryarchaeal repressor NrpR, 2-oxoglutarate, and two operators. J. Biol. Chem. 2005, 280, 5236–5241. [Google Scholar] [CrossRef] [PubMed]
- Enkh-Amgalan, J.; Kawasaki, H.; Oh-oka, H.; Seki, T. Cloning and characterization of a novel gene involved in nitrogen fixation in Heliobacterium chlorum: A possible regulatory gene. Arch. Microbiol. 2006, 186, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Enkh-Amgalan, J.; Kawasaki, H.; Seki, T. Molecular evolution of the nif gene cluster carrying nifl1 and nifl2 genes in the Gram-positive phototrophic bacterium Heliobacterium chlorum. Int. J. Syst. Evol. Microbiol. 2006, 56, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Eisen, J.A.; Nelson, K.E.; Paulsen, I.T.; Heidelberg, J.F.; Wu, M.; Dodson, R.J.; Deboy, R.; Gwinn, M.L.; Nelson, W.C.; Haft, D.H.; et al. The complete genome sequence of Chlorobium tepidum TLS, a photosynthetic, anaerobic, green-sulfur bacterium. Proc. Natl. Acad. Sci. USA 2002, 99, 9509–9514. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Toth, J.; Kasap, M. Nitrogen-fixation genes and nitrogenase activity in Clostridium acetobutylicum and Clostridium beijerinckii. J. Ind. Microbiol. Biotechnol. 2001, 27, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, S.; Muller, B.; Niazi, A.; Bongcam-Rudloff, E.; Schnurer, A. Draft genome sequence of Clostridium ultunense strain Esp, a syntrophic acetate-oxidizing bacterium. Genome Announc. 2013, 1, e0010713. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.J.; Kim, H.B.; Kim, J.; Kim, W.J.; Lee, H.; An, C.S. Organization of nif gene cluster in Frankia sp. EuIK1 strain, a symbiont of Elaeagnus umbellata. Arch. Microbiol. 2012, 194, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Welsh, E.A.; Liberton, M.; Stoeckel, J.; Loh, T.; Elvitigala, T.; Wang, C.; Wollam, A.; Fulton, R.S.; Clifton, S.W.; Jacobs, J.M.; et al. The genome of Cyanothece 51142, a unicellular diazotrophic cyanobacterium important in the marine nitrogen cycle. Proc. Natl. Acad. Sci. USA 2008, 105, 15094–15099. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Samanta, S.K.; Rey, F.E.; Wu, L.Y.; Liu, X.D.; Yan, T.F.; Zhou, J.Z.; Harwood, C.S. Functional genomic analysis of three nitrogenase isozymes in the photosynthetic bacterium Rhodopseudomonas palustris. J. Bacteriol. 2005, 187, 7784–7794. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, F.O.; Monteiro, R.A.; Wassem, R.; Cruz, L.M.; Ayub, R.A.; Colauto, N.B.; Fernandez, M.A.; Fungaro, M.H.P.; Grisard, E.C.; Hungria, M.; et al. Genome of Herbaspirillum seropedicae strain SmR1, a specialized diazotrophic endophyte of tropical grasses. PLoS Genet. 2011, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Setubal, J.C.; dos Santos, P.; Goldman, B.S.; Ertesvag, H.; Espin, G.; Rubio, L.M.; Valla, S.; Almeida, N.F.; Balasubramanian, D.; Cromes, L.; et al. Genome sequence of Azotobacter vinelandii, an obligate aerobe specialized to support diverse anaerobic metabolic processes. J. Bacteriol. 2009, 191, 4534–4545. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.L.; Yang, J.; Dou, Y.T.; Chen, M.; Ping, S.Z.; Peng, J.P.; Lu, W.; Zhang, W.; Yao, Z.Y.; Li, H.Q.; et al. Nitrogen fixation island and rhizosphere competence traits in the genome of root-associated Pseudomonas stutzeri A1501. Proc. Natl. Acad. Sci. USA 2008, 105, 7564–7569. [Google Scholar] [CrossRef] [PubMed]
- Buchko, G.W.; Robinson, H. Crystal structure of cce_0566 from Cyanothece 51142, a protein associated with nitrogen fixation in the DUF269 family. Febs Lett. 2012, 586, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.W.; Wang, L.Y.; Li, X.X.; Liu, X.M.; Hao, T.Y.; He, X.J.; Chen, S.F. Genome-wide transcriptome profiling of nitrogen fixation in Paenibacillus sp. WLY78. BMC Microbiol. 2016, 16, 10. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, T.L.; Ludwig, M.; Dixon, R.; Boyd, E.S.; Dos Santos, P.C.; Setubal, J.C.; Bryant, D.A.; Dean, D.R.; Peters, J.W. Transcriptional profiling of nitrogen fixation in Azotobacter vinelandii. J. Bacteriol. 2011, 193, 4477–4486. [Google Scholar] [CrossRef] [PubMed]
- Bickhart, D.M.; Benson, D.R. Transcriptomes of Frankia sp strain CcI3 in growth transitions. BMC Microbiol. 2011, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Lurthy, T.; Alloisio, N.; Fournier, P.; Anchisi, S.; Ponsero, A.; Normand, P.; Pujic, P.; Boubakri, H. Molecular response to nitrogen starvation by Frankia alni ACN14a revealed by transcriptomics and functional analysis with a fosmid library in Escherichia coli. Res. Microbiol. 2018, 169, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Stoeckel, J.; Welsh, E.A.; Liberton, M.; Kunnvakkam, R.; Aurora, R.; Pakrasi, H.B. Global transcriptomic analysis of Cyanothece 51142 reveals robust diurnal oscillation of central metabolic processes. Proc. Natl. Acad. Sci. USA 2008, 105, 6156–6161. [Google Scholar] [CrossRef] [PubMed]
- Klassen, G.; Pedrosa, F.O.; Souza, E.M.; Yates, M.G.; Rigo, L.U. Sequencing and functional analysis of the nifENXorf1orf2 gene cluster of Herbaspirillum seropedicae. Fems Microbiol. Lett. 1999, 181, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Klassen, G.; Pedrosa, F.D.; de Souza, E.M.; Yates, M.G.; Rigo, L.U. Nitrogenase activity of Herbaspirillum seropedicae grown under low iron levels requires the products of nifXorf1 genes. Fems Microbiol. Lett. 2003, 224, 255–259. [Google Scholar] [CrossRef]
- Jacobson, M.R.; Brigle, K.E.; Bennett, L.T.; Setterquist, R.A.; Wilson, M.S.; Cash, V.L.; Beynon, J.; Newton, W.E.; Dean, D.R. Physical and genetic map of the major nif gene-cluster from Azotobacter vinelandii. J. Bacteriol. 1989, 171, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Berge, O.; Guinebretiere, M.H.; Achouak, W.; Normand, P.; Heulin, T. Paenibacillus graminis sp nov and Paenibacillus odorifer sp nov., isolated from plant roots, soil and food. Int. J. Syst. Evol. Microbiol. 2002, 52, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Villafane, R.; Bechhofer, D.H.; Narayanan, C.S.; Dubnau, D. Replication control genes of plasmid PE194. J. Bacteriol. 1987, 169, 4822–4829. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W.; Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zhang, W.; Ding, Y.; Yao, L.; Liu, K.; Du, B. Construction of gene knock-out system for Paenibacillus polymyxa SC2. Acta Microbiol. Sin. 2013, 53, 1258–1266. [Google Scholar]
- Bach, E.; Fernandes, G.D.; Passaglia, L.M.P. How to transform a recalcitrant Paenibacillus strain: From culture medium to restriction barrier. J. Microbiol. Methods 2016, 131, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of the nif gene clusters from the representatives of anaerobic, facultative anaerobic, and aerobic N2-fixing microorganisms. Sequence of the identified regions was retrieved from GenBank and used to draw the diagram. Genes of the same color in different organisms are homologous. The orf1 and its homologous genes are colored in red.
Figure 1.
Comparison of the nif gene clusters from the representatives of anaerobic, facultative anaerobic, and aerobic N2-fixing microorganisms. Sequence of the identified regions was retrieved from GenBank and used to draw the diagram. Genes of the same color in different organisms are homologous. The orf1 and its homologous genes are colored in red.

Figure 2.
Comparison of the predicted amino acid sequences of the Paenibacillus graminis (Pg) Orf1 proteins with analogous gene products from Paenibacillus sabinae (Ps), Paenibacillus riograndensis (Pr), Clostridium ultunense (Cu), Frankia sp. EuIK1(Fsp), Herbaspirillum seropedicae (Hs), Cyanothece sp. ATCC 51,142 (Csp), Rhodopseudomonas palustris (Rp), Rhodobacter capsulatus (Rc), Azotobacter vinelandii (Av) and Azospirillum brasilense (Ab). A black background indicates conserved residues in all aligned sequences, dark grey indicates conserved residues in at least 75% of the aligned sequences, and light grey indicates conserved residues in at least 50% of the aligned sequences.
Figure 2.
Comparison of the predicted amino acid sequences of the Paenibacillus graminis (Pg) Orf1 proteins with analogous gene products from Paenibacillus sabinae (Ps), Paenibacillus riograndensis (Pr), Clostridium ultunense (Cu), Frankia sp. EuIK1(Fsp), Herbaspirillum seropedicae (Hs), Cyanothece sp. ATCC 51,142 (Csp), Rhodopseudomonas palustris (Rp), Rhodobacter capsulatus (Rc), Azotobacter vinelandii (Av) and Azospirillum brasilense (Ab). A black background indicates conserved residues in all aligned sequences, dark grey indicates conserved residues in at least 75% of the aligned sequences, and light grey indicates conserved residues in at least 50% of the aligned sequences.

Figure 3.
Neighbor-joining phylogenetic tree based on amino acid sequences of Orf1. Bootstrap analysis was performed with 1000 cycles. Only bootstrap values greater than 50% are shown at the branch points. Bar 0.1 substitutions per amino acid positions.
Figure 3.
Neighbor-joining phylogenetic tree based on amino acid sequences of Orf1. Bootstrap analysis was performed with 1000 cycles. Only bootstrap values greater than 50% are shown at the branch points. Bar 0.1 substitutions per amino acid positions.

Figure 4.
The nif genes of P. graminis RSA19 are organized in an operon as determined by RT-PCR. (A) Outline of the strategy. Primers used and amplified products (numbered) are given below the schematic representation of the genes. (B) Result of RT-PCR reactions with RNA from P. graminis RSA19 grown under N2-fixing conditions. The numbering on the top of the gels corresponds to the product numbers drawn schematically in the outline given above. RT, standard RT-PCR reaction; (−), negative control in which no reverse transcriptase was added to the RT reaction; (+), positive control in which genomic DNA was used as template in the RT-PCR.
Figure 4.
The nif genes of P. graminis RSA19 are organized in an operon as determined by RT-PCR. (A) Outline of the strategy. Primers used and amplified products (numbered) are given below the schematic representation of the genes. (B) Result of RT-PCR reactions with RNA from P. graminis RSA19 grown under N2-fixing conditions. The numbering on the top of the gels corresponds to the product numbers drawn schematically in the outline given above. RT, standard RT-PCR reaction; (−), negative control in which no reverse transcriptase was added to the RT reaction; (+), positive control in which genomic DNA was used as template in the RT-PCR.

Figure 5.
qRT-PCR analysis of transcripts of nifBHDKENXorf1hesAnifV genes of P. graminis RSA19 under N2-fixing conditions. All values indicate relative fold increase of transcription standardized against 16S rDNA transcript levels.
Figure 5.
qRT-PCR analysis of transcripts of nifBHDKENXorf1hesAnifV genes of P. graminis RSA19 under N2-fixing conditions. All values indicate relative fold increase of transcription standardized against 16S rDNA transcript levels.

Figure 6.
Effect of the different concentrations of O2 on Nitrogenase activity of P. graminis RSA19 (WT), Δorf1 (deletion mutant), Δorf1/orf1 (complementation strain) and WT/orf1 (overexpression strain). The nitrogenase activities of these strains were assayed by the C2H4 reduction method and expressed as nmol C2H4/mg protein/hr. The nitrogenase activity of the WT was used as a control. ** indicates significant differences between the control and other strains determined by LSD at P < 0.01.
Figure 6.
Effect of the different concentrations of O2 on Nitrogenase activity of P. graminis RSA19 (WT), Δorf1 (deletion mutant), Δorf1/orf1 (complementation strain) and WT/orf1 (overexpression strain). The nitrogenase activities of these strains were assayed by the C2H4 reduction method and expressed as nmol C2H4/mg protein/hr. The nitrogenase activity of the WT was used as a control. ** indicates significant differences between the control and other strains determined by LSD at P < 0.01.

Figure 7.
Nitrogenase activity of P. graminis RSA19 (WT), Δorf1 (deletion mutant), Δorf1/orf1 (complementation strain) and WT/orf1 (overexpression strain) under iron limitation. These strains were grown anaerobically in a normal nitrogen-limited medium (Control) and nitrogen-limited medium without ferric citrate (-Fe). The nitrogenase activities of these strains were assayed by C2H4 reduction method and expressed as nmol C2H4/mg protein/hr.
Figure 7.
Nitrogenase activity of P. graminis RSA19 (WT), Δorf1 (deletion mutant), Δorf1/orf1 (complementation strain) and WT/orf1 (overexpression strain) under iron limitation. These strains were grown anaerobically in a normal nitrogen-limited medium (Control) and nitrogen-limited medium without ferric citrate (-Fe). The nitrogenase activities of these strains were assayed by C2H4 reduction method and expressed as nmol C2H4/mg protein/hr.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial strains and plasmids used in this study.
| Strain or Plasmid | Genotype and/or Relevant Characteristics | Source or Reference |
|---|---|---|
| Strains | ||
| Paenibacillus graminis | ||
| RSA19 | Wild-type strain | [40] |
| Δorf1 | orf1 in-frame deletion mutant | This study |
| Δorf1/orf1 | Complementation strain of Δorf1 with orf1 carried in plasmid pHYorf1 | This study |
| WT/orf1 | orf1 overexpression strain which is a derivative of P. graminis RSA19 carrying an orf1 gene in plasmid pHYorf1 | This study |
| E. coli | ||
| JM109 | General cloning host; recA1, endA1, gyrA96, thi-1, hsdR17, supE44, relA1, Δ(lac-proAB)/F’[traD36, proAB+, lacIq, lacZΔM15] | Sangon Biotech Co. |
| Trans110 | rpsL (Str R) thr leu thi-1 lacY galK galT ara tonA tsx dam dcm supE44 Δ(lac-proAB)/F′[traD36 proAB lacIq lacZΔM15] | Transgen Biotech Co. |
| Plasmids | ||
| pHY300PLK | Multiple-copy E. coli-Bacillus shuttle vector, Tcr | TaKaRa |
| pRN5101 | Temperature-sensitive E. coli-Bacillus shuttle vector, Emr | [41] |
| pRDorf1 | orf1 deletion vector based on pRN5101 | This study |
| pHYorf1 | orf1 overexpression vector with orf1 in pHY300PLK | This study |
Table 2.
Primers used in this study.
| Primer Name | Sequence | Location/Target |
|---|---|---|
| For qRT-PCR | ||
| 16SF | TTTGTCGTCAGCCTCGTGTTCGTG | qRT-PCR for control (16S rDNA) |
| 16SR | ATCCCCACCTTCCTCCGGTTTG | |
| RBF | GCATCCGCCATGTCACTATCACC * | qRT-PCR for nifB |
| RBR | CGCACACCTTCGTCGAACACC | |
| RHF | GACTCCACACGCCTGATTCTGAAC * | qRT-PCR for nifH |
| RHR | CCGCCGCACTCTACGTTGATG * | |
| RDF | CACTGCCACCGCTCCATGAAC * | qRT-PCR for nifD |
| RDR | CACGCAGGCTCTCATAAGTCTTGG * | |
| RKF | CAGTCATCTCAGCCGCCACTTC * | qRT-PCR for nifK |
| RKR | TCCAAGCCGTCGATCAGATTGTTC * | |
| REF | CGGTCATCCCAGTGAACAGC * | qRT-PCR for nifE |
| RER | CCCGCACTGTTCATCAGCTT * | |
| RNF | ACACCGCTGATTGCAGGAATGG * | qRT-PCR for nifN |
| RNR | TGCCGTGCGAATTGCTGATCC * | |
| RXF | GCGGTGAGCTGCTAGAACTGC * | qRT-PCR for nifX |
| RXR | GGCTGCCGAACGGAACCTTAAC * | |
| ROF | GACTTCAACTGCGCCGTATCTCC * | qRT-PCR for orf1 |
| ROR | TCCGCTCTTCTCTTCCATGACTCC * | |
| RAF | GTTGAAGGAAGCGACGGTGATGG * | qRT-PCR for hesA |
| RAR | GACCAGAATCAGCTTGCCGACAC | |
| RVF | AGGAGGATATTGCGGCGATTGC | qRT-PCR for nifV |
| RVR | CCGAGACAGGAATAGACACATGGC * | |
| For construction of orf1 deletion and overexpression strains | ||
| orf1UF | CGGCCACGATGCGTCCGGCGTAGAGGATCCGCTGGGGACCCCCTATAAG | Upstream of orf1 |
| orf1UR | TAAATTTGCAGCTTGGCTTCCTCCCCTTCCC | |
| orf1DF | GGGGAGGAAGCCAAGCTGCAAATTTAGGATCGG | Downstream of orf1 |
| orf1DR | TCATGGCGACCACACCCGTCCTGTGGATCCGCTTGCTCAACTCCGCATTC | |
| nifPF | TTATAACAGGAATTCCCGGGGATCCTGCTGCTTCCTCCTCATTTG | Promoter region of nif operon |
| nifPR | ACGATTCCATTTCCCACCTCCTAAAAGTAAC | |
| orf1F | GAGGTGGGAAATGGAATCGTCGGCTAAC | orf1 ORF |
| orf1R | ATGGAAAAACGCTTTGCCCAAGCTTATCCTTCAAGGCTGAAC | |
* Primers also used in the RT-PCR.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, Q.; Liu, X.; Zhang, H.; Chen, S. Evolution and Functional Analysis of orf1 Within nif Gene Cluster from Paenibacillus graminis RSA19. Int. J. Mol. Sci. 2019, 20, 1145. https://doi.org/10.3390/ijms20051145
AMA Style
Li Q, Liu X, Zhang H, Chen S. Evolution and Functional Analysis of orf1 Within nif Gene Cluster from Paenibacillus graminis RSA19. International Journal of Molecular Sciences. 2019; 20(5):1145. https://doi.org/10.3390/ijms20051145
Chicago/Turabian StyleLi, Qin, Xiaomeng Liu, Haowei Zhang, and Sanfeng Chen. 2019. "Evolution and Functional Analysis of orf1 Within nif Gene Cluster from Paenibacillus graminis RSA19" International Journal of Molecular Sciences 20, no. 5: 1145. https://doi.org/10.3390/ijms20051145
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.