Pulmonary Hypertension and Obesity: Focus on Adiponectin

 , ,
, , 

Abstract
1. Pulmonary Hypertension: An Overview
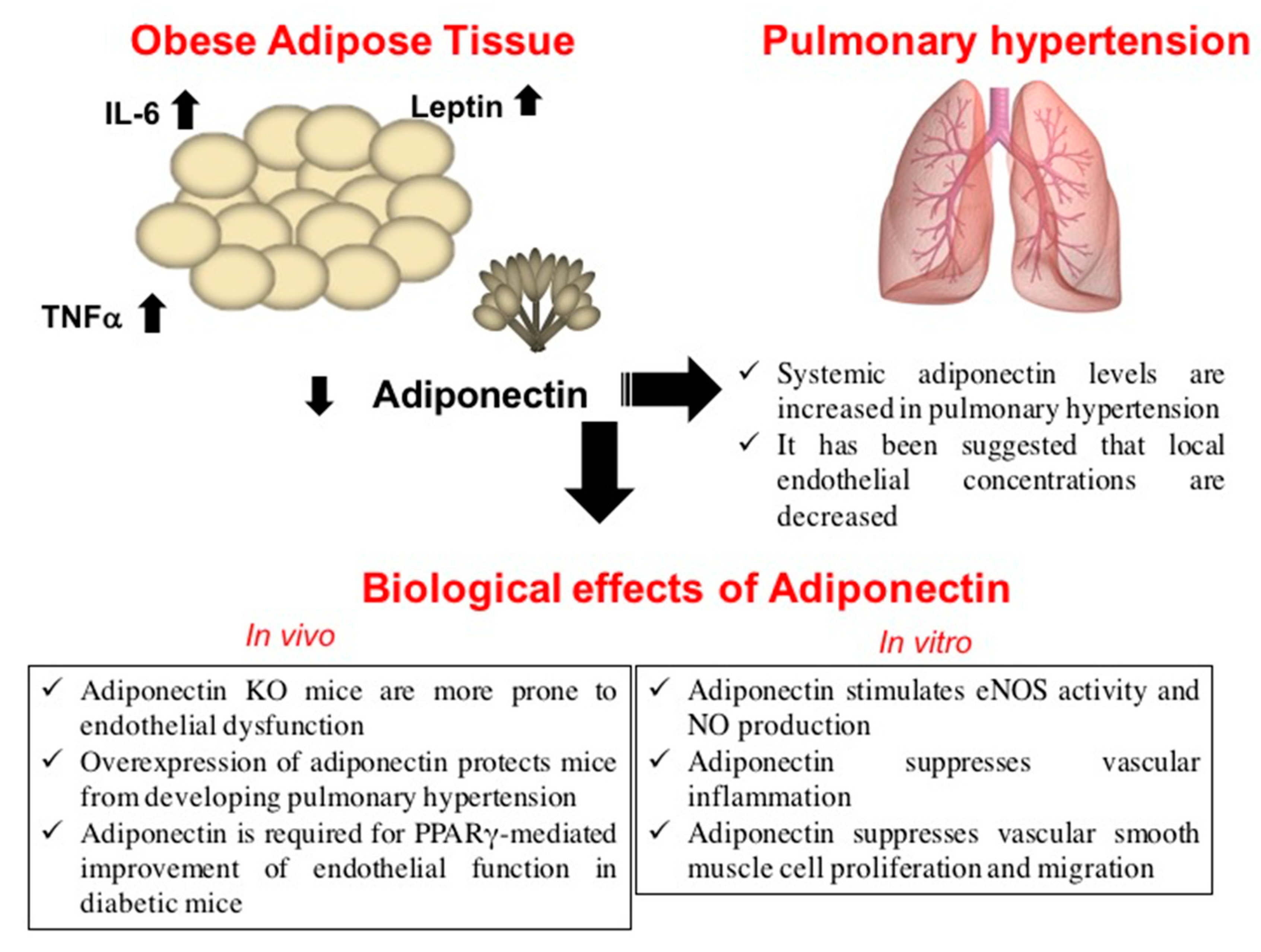
2. Obesity and Pulmonary Hypertension
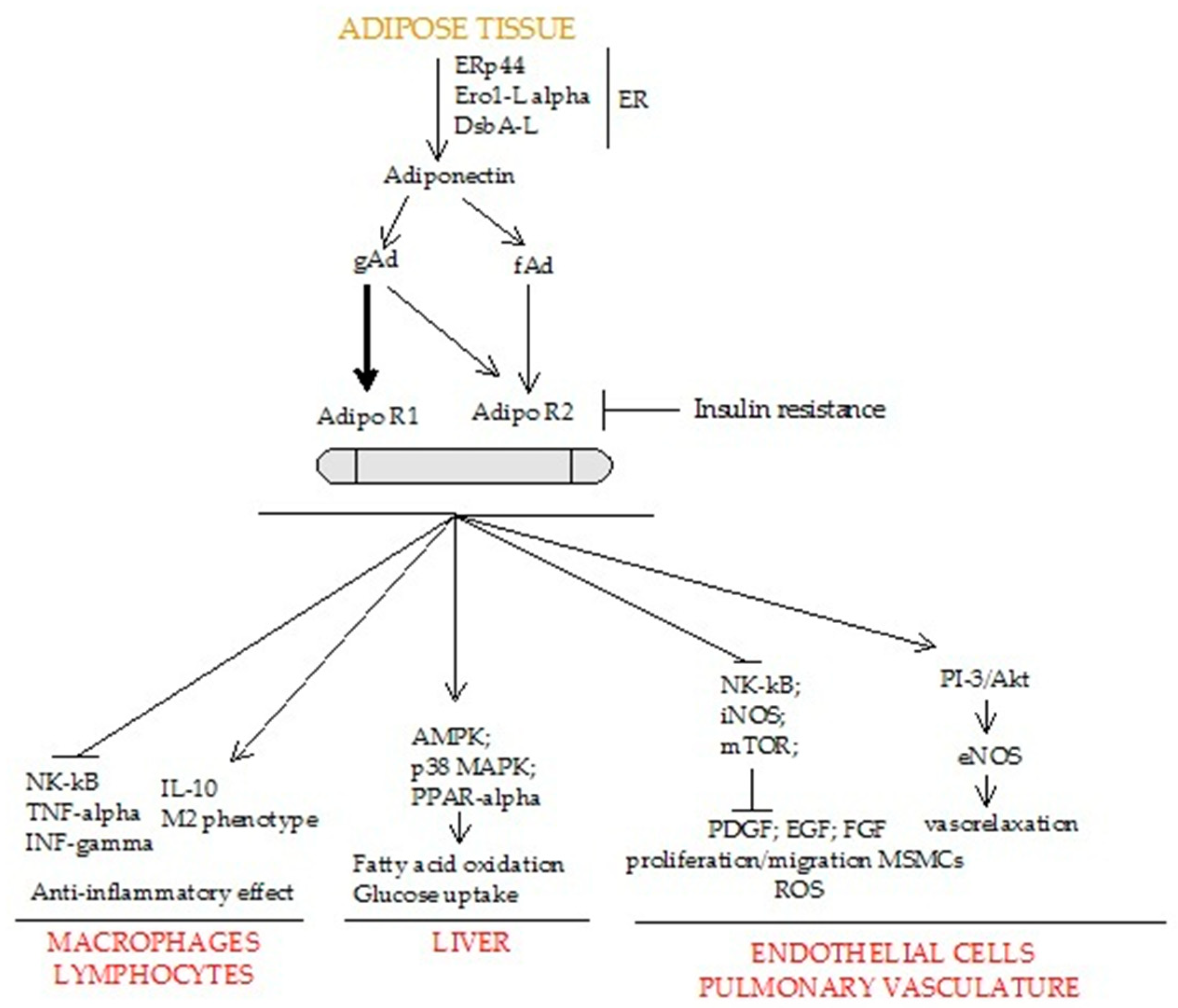
3. Biological Role of Adiponectin
4. Adiponectin Prevents Pulmonary Hypertension
5. Adiponectin Modulation in the Treatment of Pulmonary Hypertension
Funding
Conflicts of Interest
References
- Hoeper, M.M.; Bogaard, H.J.; Condliffe, R.; Frantz, R.; Khanna, D.; Kurzyna, M.; Langleben, D.; Manes, A.; Satoh, T.; Torres, F.; et al. Definitions and diagnosis of pulmonary hypertension. J. Am. Coll. Cardiol. 2013, 62, D42–D50. [Google Scholar] [CrossRef] [PubMed]
- Trisvetova, E.L.; Gubkin, S.V. Pharmacotherapy of pulmonary hypertension (according to the 2015 ESC/EAS Guidelines for the diagnosis and treatment of pulmonary hypertension). Ration. Pharmacother. Cardiol. 2016, 12, 79–86. [Google Scholar] [CrossRef]
- Guerra, G.; Perrotta, F.; Testa, G. Circulating endothelial progenitor cells biology and regenerative medicine in pulmonary vascular diseases. Curr. Pharm. Biotechnol. 2018, 19, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Johnson, C.; Mincey, K.D. Obesity Epidemiology Worldwide. Gastroenterol. Clin. N. Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed]
- McQuillan, B.M.; Picard, M.H.; Leavitt, M.; Weyman, A.E. Clinical correlates and reference intervals for pulmonary artery systolic pressure among echocardiographically normal subjects. Circulation 2001, 104, 2797–2802. [Google Scholar] [CrossRef]
- Wong, C.Y.; O’Moore-Sullivan, T.; Leano, R.; Hukins, C.; Jenkins, C.; Marwick, T.H. Association of subclinical right ventricular dysfunction with obesity. J. Am. Coll. Cardiol. 2006, 47, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Taraseviciute, A.; Voelkel, N.F. Severe pulmonary hypertension in postmenopausal obese women. Eur. J. Med. Res. 2006, 11, 198–202. [Google Scholar]
- Poms, A.D.; Turner, M.; Farber, H.W.; Meltzer, L.A.; McGoon, M.D. Comorbid conditions and outcomes in patients with pulmonary arterial hypertension: A REVEAL registry analysis. Chest 2013, 144, 169–176. [Google Scholar] [CrossRef]
- de Carvalho, M.H.C.; Colaco, A.L.; Fortes, Z.B. Cytokines, endothelial dysfunction, and insulin resistance. Arq. Bras. Endocrinol. Metabol. 2006, 50, 304–312. [Google Scholar] [CrossRef]
- Perros, F.; Montani, D.; Dorfmuller, P.; Durand-Gasselin, I.; Tcherakian, C.; Le Pavec, J.; Mazmanian, M.; Fadel, E.; Mussot, S.; Mercier, O.; et al. Platelet-derived growth factor expression and function in idiopathic pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2008, 178, 81–88. [Google Scholar] [CrossRef]
- Moschen, A.R.; Kaser, A.; Enrich, B.; Mosheimer, B.; Theurl, M.; Niederegger, H.; Tilg, H. Visfatin, an adipocytokine with proinflammatory and immunomodulating properties. J. Immunol. 2007, 178, 1748–1758. [Google Scholar] [CrossRef]
- Quarck, R.; Nawrot, T.; Meyns, B.; Delcroix, M. C-reactive protein: A new predictor of adverse outcome in pulmonary arterial hypertension. J. Am. Coll. Cardiol. 2009, 53, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Galvez-Prieto, B.; Somoza, B.; Gil-Ortega, M.; García-Prieto, C.F.; de Las Heras, A.I.; González, M.C.; Arribas, S.; Aranguez, I.; Bolbrinker, J.; Kreutz, R.; et al. Anticontractile Effect of Perivascular Adipose Tissue and Leptin are Reduced in Hypertension. Front. Pharmacol. 2012, 3, 103. [Google Scholar] [CrossRef] [PubMed]
- Huertas, A.; Tu, L.; Gambaryan, N.; Girerd, B.; Perros, F.; Montani, D.; Fabre, D.; Fadel, E.; Eddahibi, S.; Cohen-Kaminsky, S.; et al. Leptin and regulatory T-lymphocytes in idiopathic pulmonary arterial hypertension. Eur. Respir. J. 2012, 40, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Huertas, A.; Tu, L.; Thuillet, R.; Le Hiress, M.; Phan, C.; Ricard, N.; Nadaud, S.; Fadel, E.; Humbert, M.; Guignabert, C. Leptin signaling system as a target for pulmonary arterial hypertension therapy. Eur. Respir. J. 2015, 45, 1066–1080. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, A.R.; Aytekin, M.; Feldstein, A.E.; Dweik, R.A. Leptin levels predict survival in pulmonary arterial hypertension. Pulm. Circ. 2012, 2, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Korman, B.D.; Marangoni, R.G.; Hinchcliff, M.; Shah, S.J.; Carns, M.; Hoffmann, A.; Ramsey-Goldman, R.; Varga, J. Brief Report: Association of Elevated Adipsin Levels With Pulmonary Arterial Hypertension in Systemic Sclerosis. Arthritis Rheumatol. 2017, 69, 2062–2068. [Google Scholar] [CrossRef]
- Nagaya, N.; Uematsu, M.; Satoh, T.; Kyotani, S.; Sakamaki, F.; Nakanishi, N.; Yamagishi, M.; Kunieda, T.; Miyatake, K. Serum uric acid levels correlate with the severity and the mortality of primary pulmonary hypertension. Am. J. Respir. Crit. Care Med. 1999, 160, 487–492. [Google Scholar] [CrossRef]
- Zharikov, S.I.; Swenson, E.R.; Lanaspa, M.; Block, E.R.; Patel, J.M.; Johnson, R.J. Could uric acid be a modifiable risk factor in subjects with pulmonary hypertension? Med. Hypotheses 2010, 74, 1069–1074. [Google Scholar] [CrossRef]
- Friedman, S.E.; Andrus, B.W. Obesity and pulmonary hypertension: A review of pathophysiologic mechanisms. J. Obes. 2012, 2012, 505274. [Google Scholar] [CrossRef]
- Senaratna, C.V.; Perret, J.L.; Lodge, C.J.; Lowe, A.J.; Campbell, B.E.; Matheson, M.C.; Hamilton, G.S.; Dharmage, S.C. Prevalence of obstructive sleep apnea in the general population: A systematic review. Sleep Med. Rev. 2017, 34, 70–81. [Google Scholar] [CrossRef]
- Warricker, F.; Islam, Z.; Shah, B.N. Lesson of the month 1: Obesity hypoventilation (Pickwickian) syndrome: A reversible cause of severe pulmonary hypertension. Clin. Med. 2017, 17, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, W.-Q.; Zhang, H.; Yang, X.; Fan, Q.; Christopher, T.A.; Lopez, B.L.; Tao, L.; Goldstein, B.J.; Gao, F.; et al. Adiponectin improves endothelial function in hyperlipidemic rats by reducing oxidative/nitrative stress and differential regulation of eNOS/iNOS activity. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1703–E1708. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Wang, H.; Farmer, S.R. Adiponectin secretion is regulated by SIRT1 and the endoplasmic reticulum oxidoreductase Ero1-L alpha. Mol. Cell. Biol. 2007, 27, 4698–4707. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhou, L.; Xu, A.; Lam, K.S.; Wetzel, M.D.; Xiang, R.; Zhang, J.; Xin, X.; Dong, L.Q.; Liu, F. A disulfide-bond A oxidoreductase-like protein (DsbA-L) regulates adiponectin multimerization. Proc. Natl. Acad. Sci. USA 2008, 105, 18302–18307. [Google Scholar] [CrossRef]
- Magkos, F.; Sidossis, L.S. Recent advances in the measurement of adiponectin isoform distribution. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 571–575. [Google Scholar] [CrossRef]
- Fruebis, J.; Tsao, T.S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 2005–2010. [Google Scholar] [CrossRef]
- Daniele, A.; De Rosa, A.; Nigro, E.; Scudiero, O.; Capasso, M.; Masullo, M.; de Laurentiis, G.; Oriani, G.; Sofia, M.; Bianco, A. Adiponectin oligomerization state and adiponectin receptors airway expression in chronic obstructive pulmonary disease. Int. J. Biochem. Cell Biol. 2012, 44, 563–569. [Google Scholar] [CrossRef]
- Chinetti, G.; Zawadski, C.; Fruchart, J.C.; Staels, B. Expression of adiponectin receptors in human macrophages and regulation by agonists of the nuclear receptors PPARalpha, PPARgamma, and LXR. Biochem. Biophys. Res. Commun. 2004, 314, 151–158. [Google Scholar] [CrossRef]
- Kharroubi, I.; Rasschaert, J.; Eizirik, D.L.; Cnop, M. Expression of adiponectin receptors in pancreatic beta cells. Biochem. Biophys. Res. Commun. 2003, 312, 1118–1122. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M.; Bullen, J.W.J.; Lee, J.H.; Kralisch, S.; Fasshauer, M.; Klöting, N.; Niebauer, J.; Schön, M.R.; Williams, C.J.; Mantzoros, C.S. Circulating adiponectin and expression of adiponectin receptors in human skeletal muscle: Associations with metabolic parameters and insulin resistance and regulation by physical training. J. Clin. Endocrinol. Metab. 2006, 91, 2310–2316. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; Murakami, K.; Ohteki, T.; Uchida, S.; et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 2003, 423, 762–769. [Google Scholar] [CrossRef] [PubMed]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.-S.; Lodish, H.F. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef]
- Hebbard, L.W.; Garlatti, M.; Young, L.J.T.; Cardiff, R.D.; Oshima, R.G.; Ranscht, B. T-cadherin supports angiogenesis and adiponectin association with the vasculature in a mouse mammary tumor model. Cancer Res. 2008, 68, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Lacedonia, D.; Nigro, E.; Matera, M.G.; Scudiero, O.; Monaco, M.L.; Polito, R.; Carpagnano, G.E.; Foschino Barbaro, M.P.; Mazzarella, G.; Bianco, A.; et al. Evaluation of adiponectin profile in Italian patients affected by obstructive sleep apnea syndrome. Pulm. Pharmacol. Ther. 2016, 40, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.; Mazzarella, G.; Turchiarelli, V.; Nigro, E.; Corbi, G.; Scudiero, O.; Sofia, M.; Daniele, A. Adiponectin: An attractive marker for metabolic disorders in Chronic Obstructive Pulmonary Disease (COPD). Nutrients 2013, 5, 4115–4125. [Google Scholar] [CrossRef]
- Nigro, E.; Daniele, A.; Scudiero, O.; Ludovica Monaco, M.; Roviezzo, F.; D’Agostino, B.; Mazzarella, G.; Bianco, A. Adiponectin in asthma: Implications for phenotyping. Curr. Protein Pept. Sci. 2015, 16, 182–187. [Google Scholar] [CrossRef]
- Bianco, A.; Nigro, E.; Monaco, M.L.; Matera, M.G.; Scudiero, O.; Mazzarella, G.; Daniele, A. The burden of obesity in asthma and COPD: Role of adiponectin. Pulm. Pharmacol. Ther. 2017, 43, 20–25. [Google Scholar] [CrossRef]
- Nigro, E.; Scudiero, O.; Monaco, M.L.; Palmieri, A.; Mazzarella, G.; Costagliola, C.; Bianco, A.; Daniele, A. New insight into adiponectin role in obesity and obesity-related diseases. BioMed Res. Int. 2014, 2014, 658913. [Google Scholar] [CrossRef]
- Okamoto, Y.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Libby, P. Adiponectin: A key adipocytokine in metabolic syndrome. Clin. Sci. 2006, 110, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Chan, J.; Rovin, B.H. Induction of chemokine expression by adiponectin in vitro is isoform dependent. Transl. Res. 2009, 154, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Prins, J.; Venkatesh, B. Clinical review: Adiponectin biology and its role in inflammation and critical illness. Crit. Care 2011, 15, 221. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Argueta, J.G.M.; Masuhiro, Y.; Kagishita, M.; Nonaka, K.; Saito, T.; Hanazawa, S.; Yamashita, Y. Adiponectin inhibits Toll-like receptor family-induced signaling. FEBS Lett. 2005, 579, 6821–6826. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Ouchi, N.; Shibata, R.; Aprahamian, T.; Kirber, M.T.; Summer, R.S.; Kihara, S.; Walsh, K. Adiponectin modulates inflammatory reactions via calreticulin receptor-dependent clearance of early apoptotic bodies. J. Clin. Investig. 2007, 117, 375–386. [Google Scholar] [CrossRef]
- Wolf, A.M.; Wolf, D.; Rumpold, H.; Enrich, B.; Tilg, H. Adiponectin induces the anti-inflammatory cytokines IL-10 and IL-1RA in human leukocytes. Biochem. Biophys. Res. Commun. 2004, 323, 630–635. [Google Scholar] [CrossRef]
- Yokota, T.; Oritani, K.; Takahashi, I.; Ishikawa, J.; Matsuyama, A.; Ouchi, N.; Kihara, S.; Funahashi, T.; Tenner, A.J. Adiponectin, a new member of the family of soluble defense collagens, negatively regulates the growth of myelomonocytic progenitors and the functions of macrophages. Blood 2000, 96, 1723–1732. [Google Scholar]
- Lovren, F.; Pan, Y.; Quan, A.; Szmitko, P.E.; Singh, K.K.; Shukla, P.C.; Gupta, M.; Chan, L.; Al-Omran, M.; Teoh, H.; et al. Adiponectin primes human monocytes into alternative anti-inflammatory M2 macrophages. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H656–H663. [Google Scholar] [CrossRef]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; et al. Adiponectin promotes macrophage polarization toward an anti-inflammatory phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef]
- Masaie, H.; Oritani, K.; Yokota, T.; Takahashi, I.; Shirogane, T.; Ujiie, H.; Ichii, M.; Saitoh, N.; Maeda, T.; Tanigawa, R.; et al. Adiponectin binds to chemokines via the globular head and modulates interactions between chemokines and heparan sulfates. Exp. Hematol. 2007, 35, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, R.; Gong, Y.; Berzins, B.; Wu, X.; Mahadev, K.; Hough, K.; Chan, L.; Goldstein, B.J.; Scalia, R. Adiponectin deficiency increases leukocyte-endothelium interactions via upregulation of endothelial cell adhesion molecules in vivo. J. Clin. Investig. 2007, 117, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, V.; Le Cras, T.D.; Ivy, D.D.; Grover, T.R.; Kinsella, J.P.; Abman, S.H. Role of platelet-derived growth factor in vascular remodeling during pulmonary hypertension in the ovine fetus. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 284, L826–L833. [Google Scholar] [CrossRef] [PubMed]
- Christou, H.; Yoshida, A.; Arthur, V.; Morita, T.; Kourembanas, S. Increased vascular endothelial growth factor production in the lungs of rats with hypoxia-induced pulmonary hypertension. Am. J. Respir. Cell Mol. Biol. 1998, 18, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Partovian, C.; Adnot, S.; Raffestin, B.; Louzier, V.; Levame, M.; Mavier, I.M.; Lemarchand, P.; Eddahibi, S. Adenovirus-mediated lung vascular endothelial growth factor overexpression protects against hypoxic pulmonary hypertension in rats. Am. J. Respir. Cell Mol. Biol. 2000, 23, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Schermuly, R.T.; Dony, E.; Ghofrani, H.A.; Pullamsetti, S.; Savai, R.; Roth, M.; Sydykov, A.; Lai, Y.J.; Weissmann, N.; Seeger, W.; et al. Reversal of experimental pulmonary hypertension by PDGF inhibition. J. Clin. Investig. 2005, 115, 2811–2821. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; et al. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nat. Med. 2002, 8, 731–737. [Google Scholar] [CrossRef]
- Shibata, R.; Sato, K.; Pimentel, D.R.; Takemura, Y.; Kihara, S.; Ohashi, K.; Funahashi, T.; Ouchi, N.; Walsh, K. Adiponectin protects against myocardial ischemia-reperfusion injury through AMPK- and COX-2-dependent mechanisms. Nat. Med. 2005, 11, 1096–1103. [Google Scholar] [CrossRef]
- Ding, M.; Xie, Y.; Wagner, R.J.; Jin, Y.; Carrao, A.C.; Liu, L.S.; Guzman, A.K.; Powell, R.J.; Hwa, J.; Rzucidlo, E.M.; et al. Adiponectin induces vascular smooth muscle cell differentiation via repression of mammalian target of rapamycin complex 1 and FoxO4. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1403–1410. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; Maeda, K.; Kuriyama, H.; Okamoto, Y.; Kumada, M.; Hotta, K.; Nishida, M.; Takahashi, M.; et al. Adipocyte-derived plasma protein adiponectin acts as a platelet-derived growth factor-BB-binding protein and regulates growth factor-induced common postreceptor signal in vascular smooth muscle cell. Circulation 2002, 105, 2893–2898. [Google Scholar] [CrossRef]
- Schmid, P.M.; Resch, M.; Steege, A.; Fredersdorf-Hahn, S.; Stoelcker, B.; Birner, C.; Schach, C.; Buechler, C.; Riegger, G.A.; Luchner, A.; et al. Globular and full-length adiponectin induce NO-dependent vasodilation in resistance arteries of Zucker lean but not Zucker diabetic fatty rats. Am. J. Hypertens. 2011, 24, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, S.; Lind, L.; Soderberg, S.; Ingelsson, E. Associations of circulating adiponectin with measures of vascular function and morphology. J. Clin. Endocrinol. Metab. 2010, 95, 2927–2934. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.C.B.; Xu, A.; Chow, W.S.; Lam, M.C.; Ai, V.H.; Tam, S.C.; Lam, K.S. Hypoadiponectinemia is associated with impaired endothelium-dependent vasodilation. J. Clin. Endocrinol. Metab. 2004, 89, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Summer, R.; Walsh, K.; Medoff, B.D. Obesity and pulmonary arterial hypertension: Is adiponectin the molecular link between these conditions? Pulm. Circ. 2011, 1, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Ouchi, N.; Matsuzawa, Y. Adiponectin and hypertension. Am. J. Hypertens. 2011, 24, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Nigro, E.; Scudiero, O.; Sarnataro, D.; Mazzarella, G.; Sofia, M.; Bianco, A.; Daniele, A. Adiponectin affects lung epithelial A549 cell viability counteracting TNFalpha and IL-1ss toxicity through AdipoR1. Int. J. Biochem. Cell Biol. 2013, 45, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Summer, R.; Fiack, C.A.; Ikeda, Y.; Sato, K.; Dwyer, D.; Ouchi, N.; Fine, A.; Farber, H.W.; Walsh, K. Adiponectin deficiency: A model of pulmonary hypertension associated with pulmonary vascular disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L432–L438. [Google Scholar] [CrossRef]
- Weng, M.; Raher, M.J.; Leyton, P.; Combs, T.P.; Scherer, P.E.; Bloch, K.D.; Medoff, B.D. Adiponectin decreases pulmonary arterial remodeling in murine models of pulmonary hypertension. Am. J. Respir. Cell Mol. Biol. 2011, 45, 340–347. [Google Scholar] [CrossRef]
- Civelek, M.; Lusis, A.J. Systems genetics approaches to understand complex traits. Nat. Rev. Genet. 2014, 15, 34–48. [Google Scholar] [CrossRef]
- Moral-Sanz, J.; Lopez-Lopez, J.G.; Menendez, C.; Moreno, E.; Barreira, B.; Morales-Cano, D.; Escolano, L.; Fernandez-Segoviano, P.; Villamor, E.; Cogolludo, A.; et al. Different patterns of pulmonary vascular disease induced by type 1 diabetes and moderate hypoxia in rats. Exp. Physiol. 2012, 97, 676–686. [Google Scholar] [CrossRef]
- West, J.; Niswender, K.D.; Johnson, J.A.; Pugh, M.E.; Gleaves, L.; Fessel, J.P.; Hemnes, A.R. A potential role for insulin resistance in experimental pulmonary hypertension. Eur. Respir. J. 2013, 41, 861–871. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, A.; Yamauchi, T.; Ito, Y.; Hada, Y.; Maki, T.; Takekawa, S.; Kamon, J.; Kobayashi, M.; Suzuki, R.; Hara, K.; et al. Insulin/Foxo1 Pathway Regulates Expression Levels of Adiponectin Receptors and Adiponectin Sensitivity. J. Biol. Chem. 2004, 279, 30817–30822. [Google Scholar] [CrossRef]
- Santos, M.; Reis, A.; Goncalves, F.; Ferreira-Pinto, M.J.; Cabral, S.; Torres, S.; Leite-Moreira, A.F.; Henriques-Coelho, T. Adiponectin levels are elevated in patients with pulmonary arterial hypertension. Clin. Cardiol. 2014, 37, 21–25. [Google Scholar] [CrossRef]
- Kochetkova, E.A.; Ugai, L.G.; Maistrovskaia, Y.V.; Nevzorova, V.A. Adipokines: A Possible Contribution to Vascular and Bone Remodeling in Idiopathic Pulmonary Arterial Hypertension. Calcif. Tissue Int. 2017, 100, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Guo, X.; Cui, Q.; Liu, X.; Su, G.; Zhang, J. Sildenafil improves the function of endothelial cells in patients suffering from congenital heart disease with pulmonary hypertension. Pharmazie 2016, 71, 570–574. [Google Scholar] [PubMed]
- Rabinovitch, M. PPARgamma and the pathobiology of pulmonary arterial hypertension. Adv. Exp. Med. Biol. 2010, 661, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Hansmann, G.; Zamanian, R.T. PPARgamma activation: A potential treatment for pulmonary hypertension. Sci. Transl. Med. 2009, 1, 12ps14. [Google Scholar] [CrossRef] [PubMed]
- Astapova, O.; Leff, T. Adiponectin and PPARgamma: Cooperative and interdependent actions of two key regulators of metabolism. Vitam. Horm. 2012, 90, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, O.; Fujita, M.; Kato, M.; Yamazaki, S.; Asano, Y.; Ogai, A.; Okazaki, H.; Asai, M.; Nagamachi, Y.; Maeda, N.; et al. Natriuretic peptides enhance the production of adiponectin in human adipocytes and in patients with chronic heart failure. J. Am. Coll. Cardiol. 2009, 53, 2070–2077. [Google Scholar] [CrossRef] [PubMed]
- Sam, F.; Duhaney, T.-A.S.; Sato, K.; Wilson, R.M.; Ohashi, K.; Sono-Romanelli, S.; Higuchi, A.; De Silva, D.S.; Qin, F.; Walsh, K.; et al. Adiponectin deficiency, diastolic dysfunction, and diastolic heart failure. Endocrinology 2010, 151, 322–331. [Google Scholar] [CrossRef]
- Domagała-Kulawik, J.; Osińska, I.; Piechuta, A.; Bielicki, P.; Skirecki, T. T, B, and NKT Cells in Systemic Inflammation in Obstructive Sleep Apnoea. Mediators Inflamm. 2015, 2015, 161579. [Google Scholar] [CrossRef]
- Isobe, S.; Kataoka, M.; Kawakami, T.; Fukuda, K. Adiponectin in Chronic Thromboembolic Pulmonary Hypertension. Circ. J. 2018, 82, 1466–1468. [Google Scholar] [CrossRef] [PubMed]
- Shore, S.A.; Terry, R.D.; Flynt, L.; Xu, A.; Hug, C. Adiponectin attenuates allergen-induced airway inflammation and hyperresponsiveness in mice. J. Allergy Clin. Immunol. 2006, 118, 389–395. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Kishida, K.; Kihara, S.; Funahashi, T.; Shimomura, I. Adiponectin ameliorates hypoxia-induced pulmonary arterial remodeling. Biochem. Biophys. Res. Commun. 2009, 382, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Kubota, N.; Sato, H.; Inoue, M.; Kumagai, H.; Iwamura, T.; Takamoto, I.; Kobayashi, T.; Moroi, M.; Terauchi, Y.; et al. Pioglitazone Ameliorates Smooth Muscle Cell Proliferation in Cuff-Induced Neointimal Formation by Both Adiponectin-Dependent and -Independent Pathways. Sci. Rep. 2016, 6, 34707. [Google Scholar] [CrossRef]
- Li, J.; Wang, P.; Chen, Z.; Yu, S.; Xu, H. Fenofibrate Ameliorates Oxidative Stress-Induced Retinal Microvascular Dysfunction in Diabetic Rats. Curr. Eye Res. 2018, 43, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Wang, Q.; Jiang, S.; Wang, Q.; Hu, W.; Zhou, S.; Zhao, L.; Xie, L.; Chen, J.; Wellstein, A.; Lai, E.Y. Fenofibrate improves vascular endothelial function and contractility in diabetic mice. Redox Biol. 2018, 20, 87–97. [Google Scholar] [CrossRef]
- Lee, J.M.; Kim, J.H.; Son, H.S.; Hong, E.G.; Yu, J.M.; Han, K.A.; Min, K.W.; Chang, S.A. Valsartan increases circulating adiponectin levels without changing HOMA-IR in patients with type 2 diabetes mellitus and hypertension. J. Int. Med. Res. 2010, 38, 234–241. [Google Scholar] [CrossRef]
- Yenicesu, M.; Yilmaz, M.I.; Caglar, K.; Sonmez, A.; Eyileten, T.; Acikel, C.; Kilic, S.; Bingol, N.; Bingol, S.; Vural, A. Blockade of the renin-angiotensin system increases plasma adiponectin levels in type-2 diabetic patients with proteinuria. Nephron Clin. Pract. 2005, 99, c115–c121. [Google Scholar] [CrossRef]
- Schupp, M.; Janke, J.; Clasen, R.; Unger, T.; Kintscher, U. Angiotensin Type 1 Receptor Blockers Induce Peroxisome Proliferator-Activated Receptor-γ Activity. Circulation 2004, 109, 2054–2057. [Google Scholar] [CrossRef]
- Lim, S.; Quon, M.J.; Koh, K.K. Modulation of adiponectin as a potential therapeutic strategy. Atherosclerosis 2014, 233, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, Y.; Zhou, K.; Zhang, G.; Wang, Y.; Liu, T.; Zhang, Y.; Guo, A.; An, Y. Mineralocorticoid Receptor Blockade Improves Insulin Sensitivity in the Rat Heart and a Possible Molecular Mechanism. Cell. Physiol. Biochem. 2016, 39, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zheng, W.; Lian, G.; Chen, H.; Li, L.; Xu, C.; Xie, L. Combination treatment of adipose-derived stem cells and adiponectin attenuates pulmonary arterial hypertension in rats by inhibiting pulmonary arterial smooth muscle cell proliferation and regulating the AMPK/BMP/Smad pathway. Int. J. Mol. Med. 2018, 41, 51–60. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Molecule/Drug | Mechanism of Action | Authors |
|---|---|---|
| Thiazolidinediones | Synthetic ligands of PPARγ: ↑ production and secretion of adiponectin: ↓ PASMCs proliferation | Kubota et al. [85] |
| Fenofibrate | PPARγ agonist: ↑ levels of adiponectin: ↓ levels of inflammation ↑ endothelial functions | Li et al. [86], Xu et al. [87] |
| RAS blockers ARBs | ↑ levels of adiponectin ↑ PPARγ activity | Yenicesu et al. [89] Schupp et al. [90] |
| MR blockade | ↑ heart and adipose tissue adiponectin; ↑ PPARα protein expression; ↓ p38 MAPK; ↓ inflammation; ↓ insulin resistance | Wang et al. [92] |
| ADSCs and adiponectin | ↓AMPK/BMP/Smad signaling pathway ↓ PASMCs proliferation | Luo et al. [93] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrotta, F.; Nigro, E.; Mollica, M.; Costigliola, A.; D’Agnano, V.; Daniele, A.; Bianco, A.; Guerra, G. Pulmonary Hypertension and Obesity: Focus on Adiponectin. Int. J. Mol. Sci. 2019, 20, 912. https://doi.org/10.3390/ijms20040912
Perrotta F, Nigro E, Mollica M, Costigliola A, D’Agnano V, Daniele A, Bianco A, Guerra G. Pulmonary Hypertension and Obesity: Focus on Adiponectin. International Journal of Molecular Sciences. 2019; 20(4):912. https://doi.org/10.3390/ijms20040912
Chicago/Turabian StylePerrotta, Fabio, Ersilia Nigro, Mariano Mollica, Adriano Costigliola, Vito D’Agnano, Aurora Daniele, Andrea Bianco, and Germano Guerra. 2019. "Pulmonary Hypertension and Obesity: Focus on Adiponectin" International Journal of Molecular Sciences 20, no. 4: 912. https://doi.org/10.3390/ijms20040912
APA StylePerrotta, F., Nigro, E., Mollica, M., Costigliola, A., D’Agnano, V., Daniele, A., Bianco, A., & Guerra, G. (2019). Pulmonary Hypertension and Obesity: Focus on Adiponectin. International Journal of Molecular Sciences, 20(4), 912. https://doi.org/10.3390/ijms20040912