Glucagon-Like Peptide-1 (GLP-1) and 5-Hydroxytryptamine 2c (5-HT2c) Receptor Agonists in the Ventral Tegmental Area (VTA) Inhibit Ghrelin-Stimulated Appetitive Reward

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
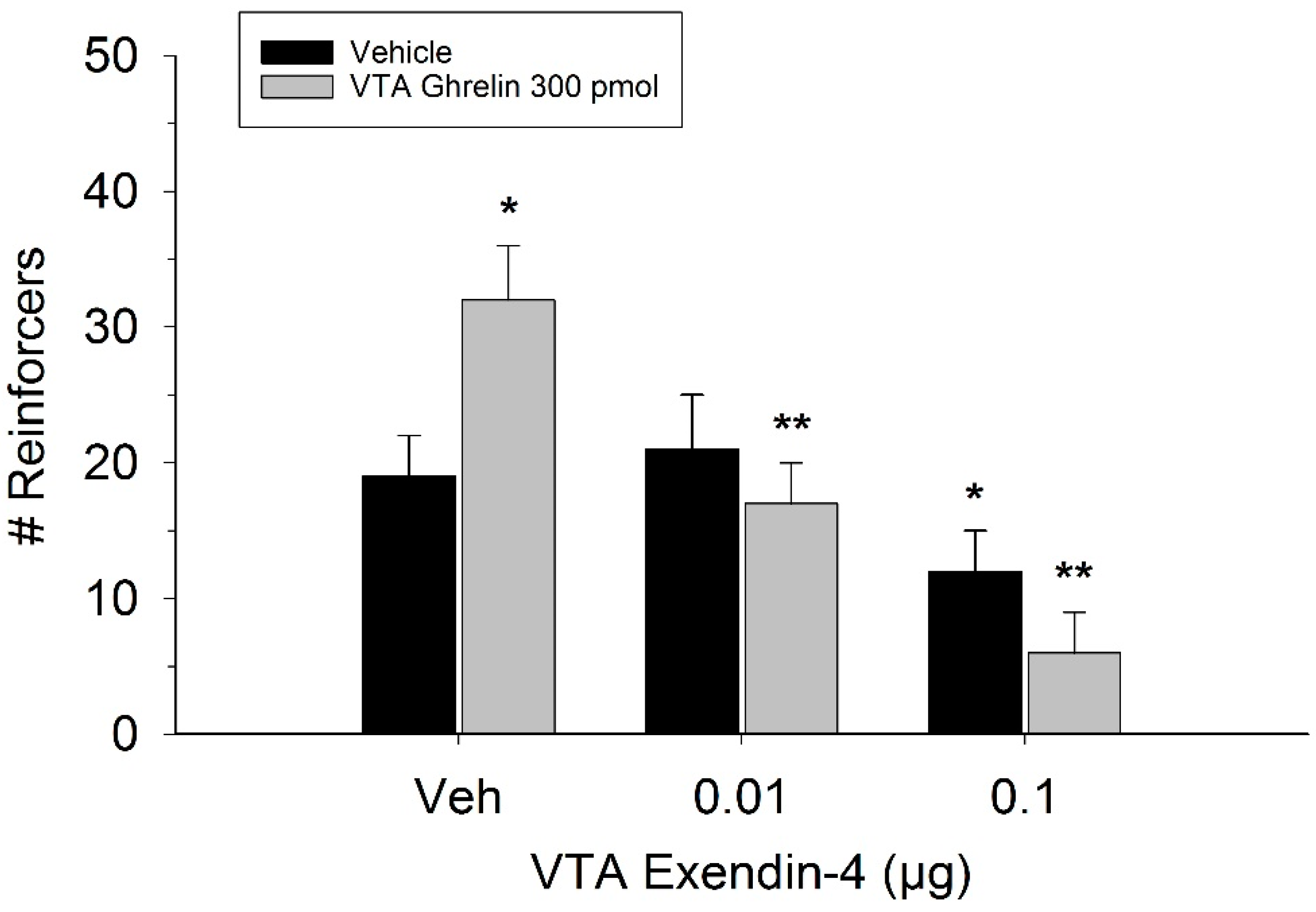
2. Results
3. Discussion
4. Methods
4.1. Animals
4.2. Peptide and Drug
4.3. Stereotaxic Surgery
4.4. Design and Procedure
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abtahi, S.; VanderJagt, H.L.; Currie, P.J. The Glucagon-Like Peptide-1 Analog Exendin-4 Antagonizes the effect of Acyl Ghrelin on the Respiratory Exchange Ratio. Neuroreport 2016, 27, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Cepko, L.C.S.; Selva, J.A.; Merfeld, E.B.; Fimmel, A.I.; Goldberg, S.A.; Currie, P.J. Ghrelin Alters the Stimulatory Effect of Cocaine on Ethanol Intake Following Mesolimbic or Systemic Administration. Neuropharmacology 2014, 85, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a Growth-Hormone-Releasing Acylated Peptide from Stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Edwards, A.; Abizaid, A. Clarifying the Ghrelin System’s Ability to Regulate Feeding Behaviours Despite Enigmatic Spatial Separation of the GHSR and Its Endogenous Ligand. Int. J. Mol. Sci. 2017, 18, 859. [Google Scholar] [CrossRef] [PubMed]
- Abtahi, S.; Howell, E.; Currie, P.J. Accumbal Ghrelin and Glucagon-Like Peptide 1 Signaling in Alcohol Reward in Female Rats. Neuroreport 2018, 29, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, V.; Watts, A.; Abizaid, A. Ghrelin Enhances Cue-Induced Bar Pressing for High Fat Food. Horm. Behav. 2016, 78, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Suchankova, P.; Steensland, P.; Fredriksson, I.; Engel, J.A.; Jerlhag, E. Ghrelin Receptor (GHS-R1A) Antagonism Suppresses Both Alcohol Consumption and the Alcohol Deprivation Effect in Rats following Long-Term Voluntary Alcohol Consumption. PLoS ONE 2013, 8, e71284. [Google Scholar] [CrossRef]
- Skibicka, K.P.; Hansson, C.; Alvarez-Crespo, M.; Friberg, P.A.; Dickson, S.L. Ghrelin Directly Targets the Ventral Tegmental Area to Increase Food Motivation. Neuroscience 2011, 180, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Skibicka, K.P.; Shirazi, R.H.; Rabasa-Papio, C.; Alvarez-Crespo, M.; Neuber, C.; Vogel, H.; Dickson, S.L. Divergent Circuitry Underlying Food Reward and Intake Effects of Ghrelin: Dopaminergic VTA-Accumbens Projection Mediates Ghrelin’s Effect on Food Reward but Not Food Intake. Neuropharmacology 2013, 73, 274–283. [Google Scholar] [CrossRef]
- Jerlhag, E.; Janson, A.C.; Waters, S.; Engel, J.A. Concomitant Release of Ventral Tegmental Acetylcholine and Accumbal Dopamine by Ghrelin in Rats. PLoS ONE 2012, 7, e49557. [Google Scholar] [CrossRef] [PubMed]
- Engel, J.A.; Nylander, I.; Jerlhag, E. A Ghrelin Receptor (GHS-R1A) Antagonist Attenuates the Rewarding Properties of Morphine and Increases Opioid Peptide Levels in Reward Areas in Mice. Eur. Neuropsychopharmacol. 2015, 25, 2364–2371. [Google Scholar] [CrossRef] [PubMed]
- Zallar, L.J.; Beurmann, S.; Tunstall, B.J.; Fraser, C.M.; Koob, G.F.; Vendruscolo, L.F.; Leggio, L. Ghrelin Receptor Deletion Reduces Binge-Like Alcohol Drinking in Rats. J. Neuroendocrinol. 2018, e12663. [Google Scholar] [CrossRef] [PubMed]
- Schuette, L.M.; Gray, C.C.; Currie, P.J. Microinjection of Ghrelin into the Ventral Tegmental Area Potentiates Cocaine-Induced Conditioned Place Preference. J. Behav. Brain Sci. 2013, 3, 576–580. [Google Scholar] [CrossRef] [PubMed]
- Abtahi, S.; Howell, E.; Salvucci, J.T.; Bastacky, J.M.R.; Dunn, D.P.; Currie, P.J. Exendin-4 Antagonizes the Metabolic Action of Acylated Ghrelinergic Signaling in the Hypothalamic Paraventricular Nucleus. Gen. Comp. Endocrinol. 2019, 270, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Currie, P.J.; John, C.S.; Nicholson, M.L.; Chapman, C.D.; Loera, K.E. Hypothalamic Paraventricular 5-Hydroxytryptamine Inhibits the Effects of Ghrelin on Eating and Energy Substrate Utilization. Pharmacol. Biochem. Behav. 2010, 97, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Holst, J.J. The Physiology of Glucagon-like Peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef] [Green Version]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 Neurons in the Nucleus of the Solitary Tract Project Directly to the Ventral Tegmental Area and Nucleus Accumbens to Control for Food Intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Merchenthaler, I.; Lane, M.; Shughrue, P. Distribution of Pre-Pro-Glucagon and Glucagon-Like Peptide-1 Receptor Messenger RNAs in the Rat Central Nervous System. J. Comp. Neurol. 1999, 403, 261–280. [Google Scholar] [CrossRef]
- Egecioglu, E.; Steensland, P.; Fredriksson, I.; Feltmann, K.; Engel, J.A.; Jerlhag, E. The Glucagon-Like Peptide 1 Analogue Exendin-4 Attenuates Alcohol Mediated behaviors in Rodents. Psychoneuroendocrinology 2013, 38, 1259–1270. [Google Scholar] [CrossRef]
- Sirohi, S.; Schurdak, J.D.; Seeley, R.J.; Benoit, S.C.; Davis, J.F. Central & Peripheral Glucagon-Like Peptide-1 Receptor Signaling Differentially Regulate Addictive Behaviors. Physiol. Behav. 2016, 161, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, N.S.; Ige, K.Y.; Mietlicki-Baase, E.G.; Molina-Castro, G.C.; Turner, C.A.; Hayes, M.R.; Schmidt, H.D. Glucagon-Like Peptide-1 Receptor Activation in the Ventral Tegmental Area Attenuates Cocaine Seeking in Rats. Neuropsychopharmacology 2018, 10, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Browne, C.J.; Fletcher, P.J. Decreased Incentive Motivation Following Knockout or Acute Blockade of the Serotonin Transporter: Role of the 5-HT2C Receptor. Neuropsychopharmacology 2016, 41, 2566–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, P.J.; Chintoh, A.F.; Sinyard, J.; Higgins, G.A. Injection of the 5-HT2C Receptor Agonist Ro60-0175 into the Ventral Tegmental Area Reduces Cocaine-Induced Locomotor Activity and Cocaine Self-Administration. Neuropsychopharmacology 2004, 29, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Price, A.E.; Anastasio, N.C.; Stutz, S.J.; Hommel, J.D.; Cunningham, K.A. Serotonin 5-HT2C Receptor Activation Suppresses Binge Intake and the Reinforcing and Motivational Properties of High-Fat Food. Front. Pharmacol. 2018, 9, 821. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; He, Y.; Cao, X.; Valencia-Torres, L.; Yan, X.; Saito, K.; Wang, C.; Yang, Y.; Hinton, A., Jr.; Zhu, L.; et al. Activation of Serotonin 2C Receptors in Dopamine Neurons Inhibits Binge-like Eating in Mice. Biol. Psychiatry 2017, 81, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Valencia-Torres, L.; Olarte-Sánchez, C.M.; Lyons, D.J.; Georgescu, T.; Greenwald-Yarnell, M.; Myers, M.G.; Bradshaw, C.M.; Heisler, L.K. Activation of Ventral Tegmental Area 5-HT2C Receptors Reduces Incentive Motivation. Neuropsychopharmacology 2017, 42, 1511–1521. [Google Scholar] [CrossRef]
- Bubar, M.J.; Stutz, S.J.; Cunningham, K.A. 5-HT2C Receptors Localize to Dopamine and GABA Neurons in the Rat Mesoaccumbens Pathway. PLoS ONE 2011, 6, e20508. [Google Scholar] [CrossRef]
- Browne, C.J.; Abela, A.R.; Chu, D.; Li, Z.; Ji, X.; Lambe, E.K.; Fletcher, P.J. Dorsal Raphe Serotonin Neurons Inhibit Operant Responding for Reward via Inputs to the Ventral Tegmental Area but Not the Nucleus Accumbens: Evidence From Studies Combining Optogenetic Stimulation and Serotonin Reuptake Inhibition. Neuropsychopharmacology 2018, 1–12. [Google Scholar] [CrossRef]
- Bake, T.; Edvardsson, C.E.; Cummings, C.J.; Dickson, S.L. Ghrelin’s Effects on Food Motivation in Rats Are Not Limited to Palatable Foods. J. Neuroendocrinol. 2018, e12665. [Google Scholar] [CrossRef]
- King, S.J.; Isaacs, A.M.; O’Farrell, E.; Abizaid, A. Motivation to Obtain Preferred Foods is Enhanced by Ghrelin in the Ventral Tegmental Area. Horm. Behav. 2011, 60, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Skov, L.J.; Jensen, M.; Christiansen, S.H.; Ratner, C.; Woldbye, D.P.D.; Holst, B. Exploring the Behavioral and Metabolic Phenotype Generated by Re-Introduction of the Ghrelin Receptor in the Ventral Tegmental Area. Int. J. Mol. Sci. 2017, 18, 914. [Google Scholar] [CrossRef] [PubMed]
- Dailey, M.J.; Moran, T.H.; Holland, P.C.; Johnson, A.W. The Antagonism of Ghrelin Alters the Appetitive Response to Learned Cues Associated with Food. Behav. Brain Res. 2016, 303, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, R.H.; Hansson, C.; Fenander, M.; Richard, J.E.; Dickson, S.L.; Nissbrandt, H.; Bergquist, F.; Skibicka, K.P. The Stomach-Derived Hormone Ghrelin Increases Impulsive Behavior. Neuropsychopharmacology 2016, 41, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, M.P.; Barrile, F.; De Francesco, P.N.; Portiansky, E.L.; Reynaldo, M.; Perello, M. Ghrelin Recruits Specific Subsets of Dopamine and GABA Neurons of Different Ventral Tegmental Area Sub-nuclei. Neuroscience 2018, 392, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E. GLP-1 Signaling and Alcohol-Mediated Behaviors; Preclinical and Clinical Evidence. Neuropharmacology 2018, 136, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.D.; Mietlicki-Baase, E.G.; Ige, K.Y.; Maurer, J.J.; Reiner, D.J.; Zimmer, D.J.; Van Nest, D.S.; Guercio, L.A.; Wimmer, M.E.; Olivos, D.R.; et al. Glucagon-Like Peptide-1 Receptor Activation in the Ventral Tegmental Area Decreases the Reinforcing Efficacy of Cocaine. Neuropsychopharmacology 2016, 41, 1917–1928. [Google Scholar] [CrossRef]
- Gagnon, J.; Baggio, L.L.; Drucker, D.J.; Brubaker, P.L. Ghrelin Is a Novel Regulator of GLP-1 Secretion. Diabetes 2015, 64, 1513–1521. [Google Scholar] [CrossRef]
- Lindqvist, A.; Shcherbina, L.; Fischer, A.-H.T.; Wierup, N. Ghrelin Is a Regulator of Glucagon-Like Peptide 1 Secretion and Transcription in Mice. Front. Endocrinol. 2017, 8, 135. [Google Scholar] [CrossRef]
- Dalvi, P.S.; Nazarians-Armavil, A.; Purser, M.J.; Belsham, D.D. Glucagon-Like Peptide-1 Receptor Agonist, Exendin-4, Regulates Feeding-Associated Neuropeptides in Hypothalamic Neurons in Vivo and in Vitro. Endocrinology 2012, 153, 2208–2222. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, P.J.; Sinyard, J.; Higgins, G.A. Genetic and Pharmacological Evidence That 5-HT2C Receptor Activation, but Not Inhibition, Affects Motivation to Feed Under a Progressive Ratio Schedule of Reinforcement. Pharmacol. Biochem. Behav. 2010, 97, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, R.H.; Richard, J.E.; Eerola, K.; Ferreras, L.L.; Nordbeck, E.B.; Hansson, C.; Nissbrandt, H.; Bergquist, F.; Gribble, F.M.; Reimann, F.; et al. Glucagon-Like Peptide-1 and its Analogues Act in the Dorsal Raphe and Modulate Central Serotonin to Reduce Appetite and Body Weight. Diabetes 2017, 66, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.Y.; Nicholson, M.L.; Currie, P.J. 6-Hydroxydopamine Lesions of the Ventral Tegmental Area Suppress Ghrelin’s Ability to Elicit Food-Reinforced Behavior. Neurosci. Lett. 2011, 499, 70–73. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Gao, Y.; Alhadeff, A.L.; Castorena, C.M.; Huang, Y.; Lieu, L.; Afrin, S.; Sun, J.; Betley, J.N.; Guo, H.; et al. Cellular and Synaptic Reorganization of Arcuate NPY/AgRP and POMC Neurons after Exercise. Mol. Metab. 2018, 18, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Zagmutt, S.; Mera, P.; Soler-Vázquez, M.C.; Herrero, L.; Serra, D. Targeting AgRP Neurons to Maintain Energy Balance: Lessons from Animal Models. Biochem. Pharmacol. 2018, 155, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-R.; Chen, H.; Zhou, J.-J.; Pradhan, G.; Sun, Y.; Pan, H.-L.; Li, D.-P. Ghrelin Receptors Mediate Ghrelin-Induced Excitation of Agouti-Related Protein/Neuropeptide Y but Not pro-Opiomelanocortin Neurons. J. Neurochem. 2017, 142, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Cowley, M.A.; Smith, R.G.; Diano, S.; Tschöp, M.; Pronchuk, N.; Grove, K.L.; Strasburger, C.J.; Bidlingmaier, M.; Esterman, M.; Heiman, M.L.; et al. The Distribution and Mechanism of Action of Ghrelin in the CNS Demonstrates a Novel Hypothalamic Circuit Regulating Energy Homeostasis. Neuron 2003, 37, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Hashiguchi, H.; Sheng, Z.; Routh, V.; Gerzanich, V.; Simard, J.M.; Bryan, J. Direct versus Indirect Actions of Ghrelin on Hypothalamic NPY Neurons. PLoS ONE 2017, 12, e0184261. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Liang, T.; Wang, G.; Li, Z. Ghrelin, a Gastrointestinal Hormone, Regulates Energy Balance and Lipid Metabolism. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed]
- Martin-Gronert, M.S.; Stocker, C.J.; Wargent, E.T.; Cripps, R.L.; Garfield, A.S.; Jovanovic, Z.; D’Agostino, G.; Yeo, G.S.H.; Cawthorne, M.A.; Arch, J.R.S.; et al. 5-HT2A and 5-HT2C Receptors as Hypothalamic Targets of Developmental Programming in Male Rats. Dis. Models Mech. 2016, 9, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Romanova, I.V.; Derkach, K.V.; Mikhrina, A.L.; Sukhov, I.B.; Mikhailova, E.V.; Shpakov, A.O. The Leptin, Dopamine and Serotonin Receptors in Hypothalamic POMC-Neurons of Normal and Obese Rodents. Neurochem. Res. 2018, 43, 821–837. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.R.; Baik, J.-H. Melanocortin 4 Receptor and Dopamine D2 Receptor Expression in Brain Areas Involved in Food Intake. Endocrinol. Metab. 2015, 30, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C. POMC Neurons: Feeding, Energy Metabolism, and Beyond. Adv. Exp. Med. Biol. 2018, 1090, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Currie, P.J.; Coiro, C.D.; Niyomchai, T.; Lira, A.; Farahmand, F. Hypothalamic Paraventricular 5-Hydroxytryptamine: Receptor-Specific Inhibition of NPY-Stimulated Eating and Energy Metabolism. Pharmacol. Biochem. Behav. 2002, 71, 709–716. [Google Scholar] [CrossRef]
- Currie, P.J.; Saxena, N.; Tu, A.Y. 5-HT(2A/2C) Receptor Antagonists in the Paraventricular Nucleus Attenuate the Action of DOI on NPY-Stimulated Eating. Neuroreport 1999, 10, 3033–3036. [Google Scholar] [CrossRef] [PubMed]
- Borkar, C.D.; Upadhya, M.A.; Shelkar, G.P.; Subhedar, N.K.; Kokare, D.M. Neuropeptide Y System in Accumbens Shell Mediates Ethanol Self-Administration in Posterior Ventral Tegmental Area. Addict. Biol. 2016, 21, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.J.; Upadhya, M.A.; Subhedar, N.K.; Kokare, D.M. NPY Mediates Reward Activity of Morphine, via NPY Y1 Receptors, in the Nucleus Accumbens Shell. Behav. Brain Res. 2013, 247, 79–91. [Google Scholar] [CrossRef]
- Skibicka, K.P.; Hansson, C.; Egecioglu, E.; Dickson, S.L. Role of Ghrelin in Food Reward: Impact of Ghrelin on Sucrose Self-Administration and Mesolimbic Dopamine and Acetylcholine Receptor Gene Expression. Addict. Biol. 2012, 17, 95–107. [Google Scholar] [CrossRef]
- Skibicka, K.P.; Shirazi, R.H.; Hansson, C.; Dickson, S.L. Ghrelin Interacts with Neuropeptide Y Y1 and Opioid Receptors to Increase Food Reward. Endocrinology 2012, 153, 1194–1205. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, G.; Jensen, M.; Weikop, P.; Dencker, D.; Christiansen, S.H.; Loland, C.J.; Bengtsen, C.H.; Petersen, J.H.; Fink-Jensen, A.; Wörtwein, G.; et al. Neuropeptide Y Y5 Receptor Antagonism Attenuates Cocaine-Induced Effects in Mice. Psychopharmacology 2012, 222, 565–577. [Google Scholar] [CrossRef]
- West, K.S.; Roseberry, A.G. Neuropeptide-Y alters VTA Dopamine Neuron Activity Through Both Pre- and Post-Synaptic Mechanisms. J. Neurophysiol. 2017, 118, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Sustkova-Fiserova, M.; Charalambous, C.; Havlickova, T.; Lapka, M.; Jerabek, P.; Puskina, N.; Syslova, K. Alterations in Rat Accumbens Endocannabinoid and GABA Content during Fentanyl Treatment: The Role of Ghrelin. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Jiang, C.; Liu, P.; Wang, F.; Ma, L. Mesolimbic Leptin Signaling Negatively Regulates Cocaine-Conditioned Reward. Transl. Psychiatry 2016, 6, e972. [Google Scholar] [CrossRef] [PubMed]
- Van der Plasse, G.; van Zessen, R.; Luijendijk, M.C.M.; Erkan, H.; Stuber, G.D.; Ramakers, G.M.J.; Adan, R.A.H. Modulation of Cue-Induced Firing of Ventral Tegmental Area Dopamine Neurons by Leptin and Ghrelin. Int. J. Obes. 2015, 39, 1742–1749. [Google Scholar] [CrossRef]
- You, Z.-B.; Wang, B.; Liu, Q.-R.; Wu, Y.; Otvos, L.; Wise, R.A. Reciprocal Inhibitory Interactions Between the Reward-Related Effects of Leptin and Cocaine. Neuropsychopharmacology 2016, 41, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.; Omrani, A.; Luijendijk, M.C.M.; de Vrind, V.A.J.; Van Rozen, A.J.; Ophuis, R.J.O.; Garner, K.; Kallo, I.; Ghanem, A.; Liposits, Z.; et al. Melanocortin 3 Receptor Signaling in Midbrain Dopamine Neurons Increases the Motivation for Food Reward. Neuropsychopharmacology 2016, 41, 2241–2251. [Google Scholar] [CrossRef] [PubMed]
- Hsu, R.; Taylor, J.R.; Newton, S.S.; Alvaro, J.D.; Haile, C.; Han, G.; Hruby, V.J.; Nestler, E.J.; Duman, R.S. Blockade of Melanocortin Transmission Inhibits Cocaine Reward. Eur. J. Neurosci. 2005, 21, 2233–2242. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 7th ed.; Academic Press: New York, NY, USA, 2014. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howell, E.; Baumgartner, H.M.; Zallar, L.J.; Selva, J.A.; Engel, L.; Currie, P.J. Glucagon-Like Peptide-1 (GLP-1) and 5-Hydroxytryptamine 2c (5-HT2c) Receptor Agonists in the Ventral Tegmental Area (VTA) Inhibit Ghrelin-Stimulated Appetitive Reward. Int. J. Mol. Sci. 2019, 20, 889. https://doi.org/10.3390/ijms20040889
Howell E, Baumgartner HM, Zallar LJ, Selva JA, Engel L, Currie PJ. Glucagon-Like Peptide-1 (GLP-1) and 5-Hydroxytryptamine 2c (5-HT2c) Receptor Agonists in the Ventral Tegmental Area (VTA) Inhibit Ghrelin-Stimulated Appetitive Reward. International Journal of Molecular Sciences. 2019; 20(4):889. https://doi.org/10.3390/ijms20040889
Chicago/Turabian StyleHowell, Erin, Hannah M. Baumgartner, Lia J. Zallar, Joaquín A. Selva, Liv Engel, and Paul J. Currie. 2019. "Glucagon-Like Peptide-1 (GLP-1) and 5-Hydroxytryptamine 2c (5-HT2c) Receptor Agonists in the Ventral Tegmental Area (VTA) Inhibit Ghrelin-Stimulated Appetitive Reward" International Journal of Molecular Sciences 20, no. 4: 889. https://doi.org/10.3390/ijms20040889