MicroRNA-766-3p Contributes to Anti-Inflammatory Responses through the Indirect Inhibition of NF-κB Signaling

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Changes in Plasma miRNA Levels before and after Abatacept Treatment in Patients with RA
2.2. Suppression of TNF-Induced Inflammatory Responses by hsa-miR-766-3p
2.3. Blunted Induction of Inflammatory Responses in miR-766-3p-Treated MH7A Cells
2.4. Involvement of miR-766-3p in the Suppression of Cytokine-Induced NF-κB Activation
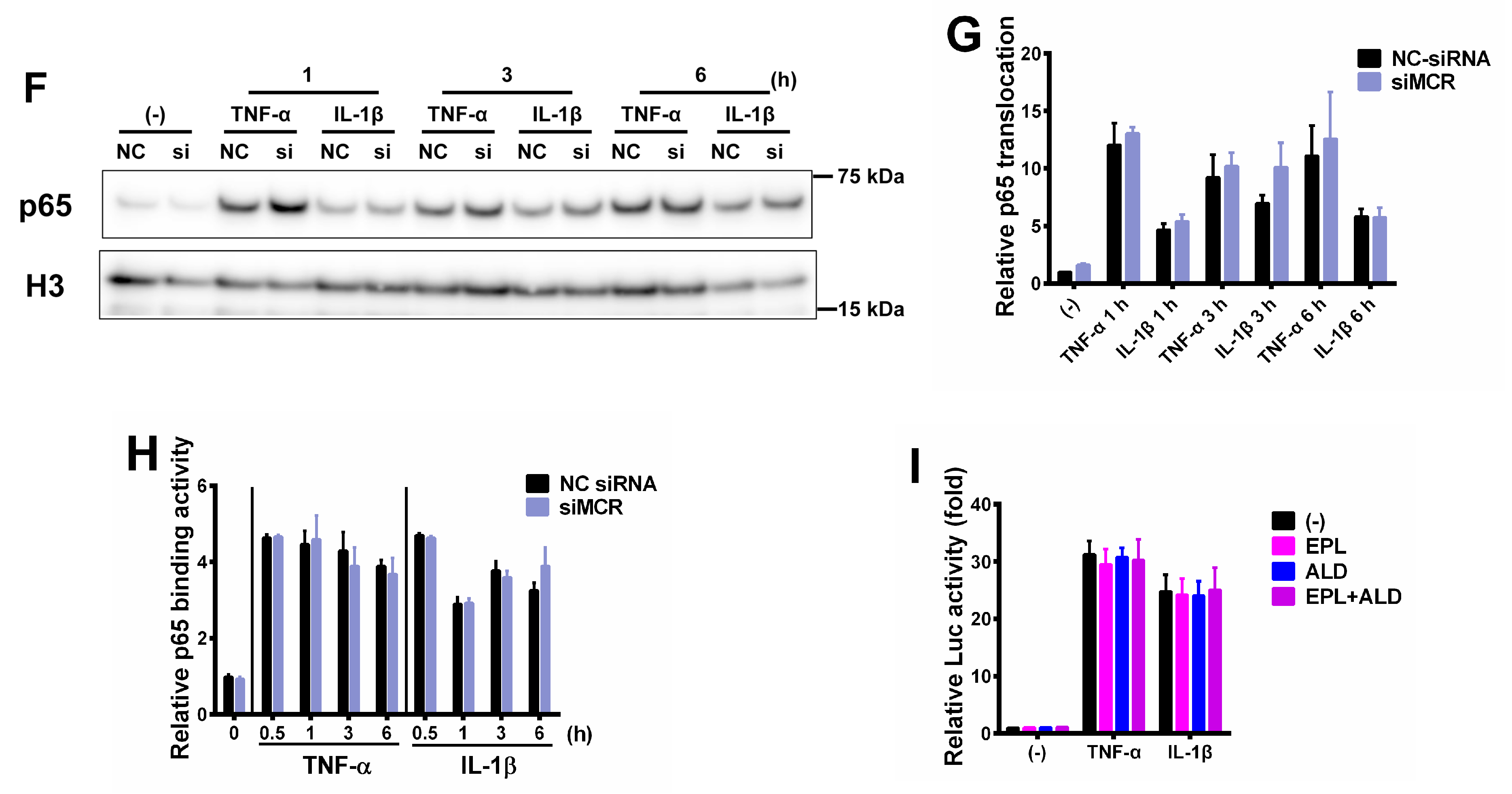
2.5. Mineralocorticoid Receptor Contributes to the miR-766-3p-Induced Anti-Inflammatory Response
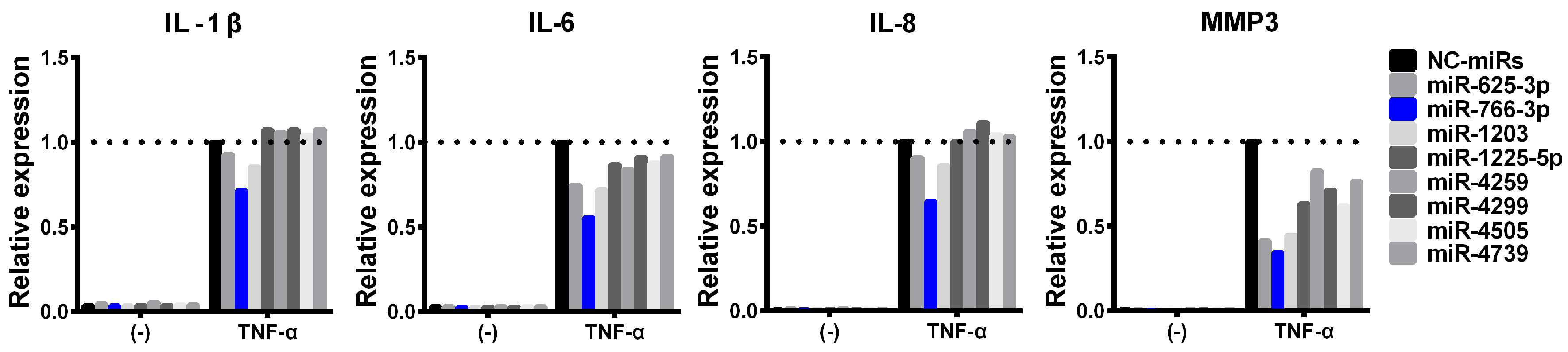
2.6. Generalizability of the Suppression of Inflammatory Cytokines in mR-766-3p-Transfected Cells
3. Discussion
4. Materials and Methods
4.1. Clinical Specimens
4.2. miRNA Array Analyses
4.3. Reagents
4.4. Cells
4.5. Transient Transfectants
4.6. Co-Culture of MH7A with PBMCs
4.7. RNA Extraction and qPCR
4.8. Western Blotting
4.9. Formazan Assays
4.10. Evaluation of DNA Binding Activity of NF-κB.
4.11. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| miRNA | microRNA |
| RA | rheumatoid arthritis |
| TNF-α | tumor necrosis factor-α |
| MMP | matrix metalloproteinase |
| IL | Interleukin |
| qPCR | quantitative polymerase chain reaction |
| PBMC | peripheral blood mononuclear cell |
| LPS | lipopolysaccharide |
| NC | negative control |
| NF-κB | nuclear factor-κB |
| MCR | Mineralocorticoid receptor |
| siRNA | small interfering RNA |
| ALD | aldosterone |
| EPL | eplerenone |
| NHMC | normal human mesangial cells |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
References
- Alevizos, I.; Illei, G.G. MicroRNAs as biomarkers in rheumatic diseases. Nat. Rev. Rheumatol. 2010, 6, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating microRNAs: Novel biomarkers and extracellular communicators in cardiovascular disease? Circ. Res. 2012, 110, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Hirai, T.; Ikeda, K.; Tsushima, H.; Fujishiro, M.; Hayakawa, K.; Yoshida, Y.; Morimoto, S.; Yamaji, K.; Takasaki, Y.; Takamori, K.; et al. Circulating plasma microRNA profiling in patients with polymyositis/dermatomyositis before and after treatment: miRNA may be associated with polymyositis/dermatomyositis. Inflamm. Regen. 2018, 38, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, H.L.; Moots, R.J.; Edwards, S.W. The multifactorial role of neutrophils in rheumatoid arthritis. Nat. Rev. Rheumatol. 2014, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Morimoto, S.; Fujishiro, M.; Hayakawa, K.; Suzuki, S.; Ikeda, K.; Miyazawa, K.; Morioka, M.; Takamori, K.; Ogawa, H.; et al. Inhibition of each module of connective tissue growth factor as a potential therapeutic target for rheumatoid arthritis. Autoimmunity 2016, 49, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Noss, E.H.; Brenner, M.B. The role and therapeutic implications of fibroblast-like synoviocytes in inflammation and cartilage erosion in rheumatoid arthritis. Immunol. Rev. 2008, 223, 252–270. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Yamada, T.; Takahashi, K.; Munakata, Y.; Hosaka, S.; Takahashi, H.; Gao, J.; Shirai, Y.; Kodama, S.; Asai, Y.; et al. MicroRNAs 106b and 222 Improve Hyperglycemia in a Mouse Model of Insulin-Deficient Diabetes via Pancreatic beta-Cell Proliferation. EBioMedicine 2017, 15, 163–172. [Google Scholar] [CrossRef]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, F.; Convertino, R.; Molino, N.; Provenzano, C.; Marino, M.; Zoli, A.; Bartoccioni, E. Effect of pro-inflammatory/anti-inflammatory agents on cytokine secretion by peripheral blood mononuclear cells in rheumatoid arthritis and systemic lupus erythematosus. Autoimmunity 2003, 36, 71–77. [Google Scholar] [CrossRef]
- Xu, J.; Itoh, Y.; Hayashi, H.; Takii, T.; Miyazawa, K.; Onozaki, K. Dihydrotestosterone inhibits interleukin-1alpha or tumor necrosis factor alpha-induced proinflammatory cytokine production via androgen receptor-dependent inhibition of nuclear factor-kappaB activation in rheumatoid fibroblast-like synovial cell line. Biol. Pharm. Bull. 2011, 34, 1724–1730. [Google Scholar] [CrossRef] [PubMed]
- Whitmarsh, A.J.; Davis, R.J. Transcription factor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. 1996, 74, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ma, X.; Guan, G.; Liu, H.; Yang, Y.; Niu, Q.; Wu, Z.; Jiang, Y.; Bian, C.; Zang, Y.; et al. MicroRNA-766 promotes cancer progression by targeting NR3C2 in hepatocellular carcinoma. FASEB J. 2019, 33, 1456–1467. [Google Scholar] [CrossRef] [PubMed]
- Leroy, V.; De Seigneux, S.; Agassiz, V.; Hasler, U.; Rafestin-Oblin, M.E.; Vinciguerra, M.; Martin, P.Y.; Feraille, E. Aldosterone activates NF-kappaB in the collecting duct. J. Am. Soc. Nephrol. 2009, 20, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.Y.; Li, C.; Shen, Z.X.; Zhang, W.C.; Ai, T.J.; Du, L.J.; Zhang, Y.Y.; Yao, G.F.; Liu, Y.; Sun, S.; et al. Mineralocorticoid Receptor Deficiency in Macrophages Inhibits Neointimal Hyperplasia and Suppresses Macrophage Inflammation Through SGK1-AP1/NF-kappaB Pathways. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Meng, Y.; Hiramatsu, N.; Kasai, A.; Yamauchi, K.; Yao, J.; Kitamura, M. Priming of glomerular mesangial cells by activated macrophages causes blunted responses to proinflammatory stimuli. J. Immunol. 2006, 176, 2529–2537. [Google Scholar] [CrossRef] [PubMed]
- Lucio-Cazana, J.; Nakayama, K.; Xu, Q.; Konta, T.; Moreno-Manzano, V.; Furusu, A.; Kitamura, M. Suppression of constitutive but not Il-1beta-inducible expression of monocyte chemoattractant protein-1 in mesangial cells by retinoic acids: Intervention in the activator protein-1 pathway. J. Am. Soc. Nephrol. 2001, 12, 688–694. [Google Scholar] [PubMed]
- Hummel, R.; Wang, T.; Watson, D.I.; Michael, M.Z.; Van der Hoek, M.; Haier, J.; Hussey, D.J. Chemotherapy-induced modification of microRNA expression in esophageal cancer. Oncol. Rep. 2011, 26, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Skrygan, M.; Georgas, D.; Sand, D.; Hahn, S.A.; Gambichler, T.; Altmeyer, P.; Bechara, F.G. Microarray analysis of microRNA expression in cutaneous squamous cell carcinoma. J. Dermatol. Sci. 2012, 68, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Li, C.F.; Chen, L.B.; Li, D.D.; Yang, L.; Jin, J.P.; Zhang, B. MicroRNA-766 targeting regulation of SOX6 expression promoted cell proliferation of human colorectal cancer. Oncol. Targets Ther. 2015, 8, 2981–2988. [Google Scholar] [CrossRef]
- Milagro, F.I.; Miranda, J.; Portillo, M.P.; Fernandez-Quintela, A.; Campion, J.; Martinez, J.A. High-throughput sequencing of microRNAs in peripheral blood mononuclear cells: Identification of potential weight loss biomarkers. PLoS ONE 2013, 8, e54319. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.; Akerstrom, T.; Rinnov, A.; Yfanti, C.; Scheele, C.; Pedersen, B.K.; Laye, M.J. The miRNA plasma signature in response to acute aerobic exercise and endurance training. PLoS ONE 2014, 9, e87308. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Xue, S.; Zhang, J.; Chen, W.; Gong, D.; Zheng, J.; Ma, J.; Xue, W.; Chen, Y.; Zhai, W.; et al. DNA-methylation-mediated repression of miR-766-3p promotes cell proliferation via targeting SF2 expression in renal cell carcinoma. Int. J. Cancer 2017, 141, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.; Lee, D.S. In vivo validation of metastasis-regulating microRNA-766 in human triple-negative breast cancer cells. Lab. Anim. Res. 2017, 33, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Li, X.; Wang, L.; Yu, S.; Xu, Z.; Gu, Y.; Pan, Z.; Li, T.; Hu, M.; Cui, H.; et al. MicroRNAs contribute to promyelocyte apoptosis in As2O3-treated APL cells. Cell. Physiol. Biochem. 2013, 32, 1818–1829. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Diecke, S.; Zhang, W.Y.; Lan, F.; He, C.; Mordwinkin, N.M.; Chua, K.F.; Wu, J.C. The role of SIRT6 protein in aging and reprogramming of human induced pluripotent stem cells. J. Biol. Chem. 2013, 288, 18439–18447. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Zhang, S.; Bair, E.; Nares, S.; Khan, A.A. Differential expression of microRNAs in normal and inflamed human pulps. J. Endod. 2012, 38, 746–752. [Google Scholar] [CrossRef]
- Jafari Ghods, F.; Topal Sarikaya, A.; Arda, N.; Hamuryudan, V. MiRNA and mRNA Profiling in Systemic Lupus Reveals a Novel Set of Cytokine—Related miRNAs and their Target Genes in Cases with and Without Renal Involvement. Kidney Blood Press. Res. 2017, 42, 1322–1337. [Google Scholar] [CrossRef]
- Alam, H.; Gu, B.; Lee, M.G. Histone methylation modifiers in cellular signaling pathways. Cell. Mol. Life Sci. 2015, 72, 4577–4592. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic Potential of Mesenchymal Cell-Derived miRNA-150-5p-Expressing Exosomes in Rheumatoid Arthritis Mediated by the Modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef]
- Zhang, S.G.; Liu, C.Y.; Li, L.; Sun, T.W.; Luo, Y.G.; Yun, W.J.; Zhang, J.Y. Examination of artificial MiRNA mimics with centered-site complementarity for gene targeting. PLoS ONE 2013, 8, e72062. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Mori, A.; Okudaira, H. Establishment and characterization of a novel human rheumatoid fibroblast-like synoviocyte line, MH7A, immortalized with SV40 T antigen. J. Biochem. 1998, 124, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Meng, Y.; Hiramatsu, N.; Kasai, A.; Yao, J.; Kitamura, M. Spontaneous activation of the NF-kappaB signaling pathway in isolated normal glomeruli. Am. J. Physiol. Renal. Physiol. 2006, 291, F1169–F1176. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA Name (miRBase ver.17) | ID | Detection Number (Before) | Detection Number (After) | p Value | Fold |
|---|---|---|---|---|---|
| Before/After | |||||
| Down-regulated miRNAs compered to before treatment | |||||
| hsa-miR-4459 | MIMAT0018981 | 10 | 10 | 0.043 | −1.99 |
| hsa-miR-625* | MIMAT0004808 | 10 | 9 | 0.002 | −1.80 |
| hsa-miR-146a | MIMAT0000449 | 5 | 8 | 0.041 | −1.73 |
| hsa-miR-4505 | MIMAT0019041 | 10 | 10 | 0.021 | −1.72 |
| hsa-miR-4520a-5p, hsa-miR-4520b-5p | MIMAT0019235, MIMAT0020299 | 4 | 4 | 0.020 | −1.66 |
| hsa-miR-4739 | MIMAT0019868 | 10 | 10 | 0.004 | −1.58 |
| hsa-miR-3616-3p | MIMAT0017996 | 7 | 8 | 0.038 | −1.42 |
| hsa-miR-766 | MIMAT0003888 | 10 | 10 | 0.009 | −1.38 |
| hsa-miR-4442 | MIMAT0018960 | 10 | 10 | 0.043 | −1.28 |
| hsa-miR-149 | MIMAT0000450 | 7 | 5 | 0.026 | −1.28 |
| hsa-miR-4675 | MIMAT0019757 | 9 | 10 | 0.033 | −1.27 |
| hsa-miR-3189-3p | MIMAT0015071 | 5 | 3 | 0.031 | −1.23 |
| hsa-miR-3197 | MIMAT0015082 | 10 | 10 | 0.050 | −1.22 |
| hsa-miR-4716-5p | MIMAT0019826 | 9 | 7 | 0.050 | −1.19 |
| hsa-miR-4508 | MIMAT0019045 | 10 | 10 | 0.039 | −1.16 |
| hsa-miR-409-3p | MIMAT0001639 | 4 | 4 | 0.025 | −1.03 |
| hsa-miR-1288 | MIMAT0005942 | 3 | 8 | 0.031 | −1.03 |
| Up-regulated miRNAs compered to before treatment | |||||
| hsa-miR-3663-5p | MIMAT0018084 | 8 | 6 | 0.038 | 1.05 |
| hsa-miR-3713 | MIMAT0018164 | 5 | 5 | 0.048 | 1.08 |
| hsa-miR-760 | MIMAT0004957 | 10 | 10 | 0.037 | 1.16 |
| hsa-miR-3202 | MIMAT0015089 | 5 | 5 | 0.006 | 1.17 |
| hsa-miR-4317 | MIMAT0016872 | 4 | 5 | 0.038 | 1.20 |
| hsa-miR-657 | MIMAT0003335 | 7 | 6 | 0.043 | 1.20 |
| hsa-miR-4728-3p | MIMAT0019850 | 9 | 9 | 0.014 | 1.24 |
| hsa-miR-4651 | MIMAT0019715 | 10 | 10 | 0.031 | 1.33 |
| hsa-miR-4640-5p | MIMAT0019699 | 10 | 10 | 0.027 | 1.37 |
| hsa-miR-3648 | MIMAT0018068 | 10 | 10 | 0.048 | 1.42 |
| hsa-miR-762 | MIMAT0010313 | 10 | 10 | 0.040 | 1.42 |
| hsa-miR-4669 | MIMAT0019749 | 9 | 9 | 0.037 | 1.43 |
| hsa-miR-1193 | MIMAT0015049 | 9 | 10 | 0.046 | 1.46 |
| hsa-miR-1915 | MIMAT0007892 | 10 | 10 | 0.048 | 1.47 |
| hsa-miR-1225-5p | MIMAT0005572 | 9 | 10 | 0.015 | 1.49 |
| hsa-miR-124 | MIMAT0000422 | 8 | 5 | 0.026 | 1.49 |
| hsa-miR-4299 | MIMAT0016851 | 10 | 10 | 0.004 | 1.58 |
| hsa-miR-3942-3p | MIMAT0019230 | 4 | 3 | 0.037 | 1.62 |
| hsa-miR-3622a-3p | MIMAT0018004 | 7 | 5 | 0.025 | 1.66 |
| hsa-miR-1203 | MIMAT0005866 | 9 | 10 | 0.032 | 1.79 |
| hsa-miR-1231 | MIMAT0005586 | 6 | 7 | 0.047 | 1.80 |
| hsa-miR-4326 | MIMAT0016888 | 6 | 10 | 0.047 | 1.98 |
| hsa-miR-2276 | MIMAT0011775 | 6 | 6 | 0.025 | 2.00 |
| hsa-miR-4664-3p | MIMAT0019738 | 6 | 8 | 0.044 | 2.26 |
| hsa-miR-4259 | MIMAT0016880 | 10 | 9 | 0.012 | 2.43 |
| Characteristics | Before | 3 Months After * | 1 Year After |
|---|---|---|---|
| Number of patients | 10 | 9 | 10 |
| Sex, male/female | 2/8 | 2/7 | 2/8 |
| Age (years) | 54.7 ± 13.4 | 55.9 ± 13.7 | 54.7 ± 13.4 |
| Disease duration (months) | 11.6 ± 7.2 | 12.0 ± 7.5 | 11.6 ± 7.2 |
| ESR (mm/h) | 39.4 ± 24.1 | 24.3 ± 16.4 | 20.8 ± 12.4 |
| C-reactive protein (mg/L) | 1.19 ± 1.45 | 0.30 ± 0.61 | 0.23 ± 0.39 |
| MMP3 (ng/mL) | 166 ± 137 | 212 ± 152 | 143 ± 97 |
| DAS28-CRP | 4.20 ± 1.15 | 2.66 ± 0.86 | 1.94 ± 0.67 |
| Remission (<2.3) | 0 | 3 | 7 |
| low disease activity (<2.7) | 0 | 2 | 2 |
| moderate disease activity (2.7–4.1) | 5 | 4 | 1 |
| high disease activity (>4.1) | 5 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayakawa, K.; Kawasaki, M.; Hirai, T.; Yoshida, Y.; Tsushima, H.; Fujishiro, M.; Ikeda, K.; Morimoto, S.; Takamori, K.; Sekigawa, I. MicroRNA-766-3p Contributes to Anti-Inflammatory Responses through the Indirect Inhibition of NF-κB Signaling. Int. J. Mol. Sci. 2019, 20, 809. https://doi.org/10.3390/ijms20040809
Hayakawa K, Kawasaki M, Hirai T, Yoshida Y, Tsushima H, Fujishiro M, Ikeda K, Morimoto S, Takamori K, Sekigawa I. MicroRNA-766-3p Contributes to Anti-Inflammatory Responses through the Indirect Inhibition of NF-κB Signaling. International Journal of Molecular Sciences. 2019; 20(4):809. https://doi.org/10.3390/ijms20040809
Chicago/Turabian StyleHayakawa, Kunihiro, Mikiko Kawasaki, Takuya Hirai, Yuko Yoshida, Hiroshi Tsushima, Maki Fujishiro, Keigo Ikeda, Shinji Morimoto, Kenji Takamori, and Iwao Sekigawa. 2019. "MicroRNA-766-3p Contributes to Anti-Inflammatory Responses through the Indirect Inhibition of NF-κB Signaling" International Journal of Molecular Sciences 20, no. 4: 809. https://doi.org/10.3390/ijms20040809