Natural Flavonol, Myricetin, Enhances the Function and Survival of Cryopreserved Hepatocytes In Vitro and In Vivo

 , and
, and 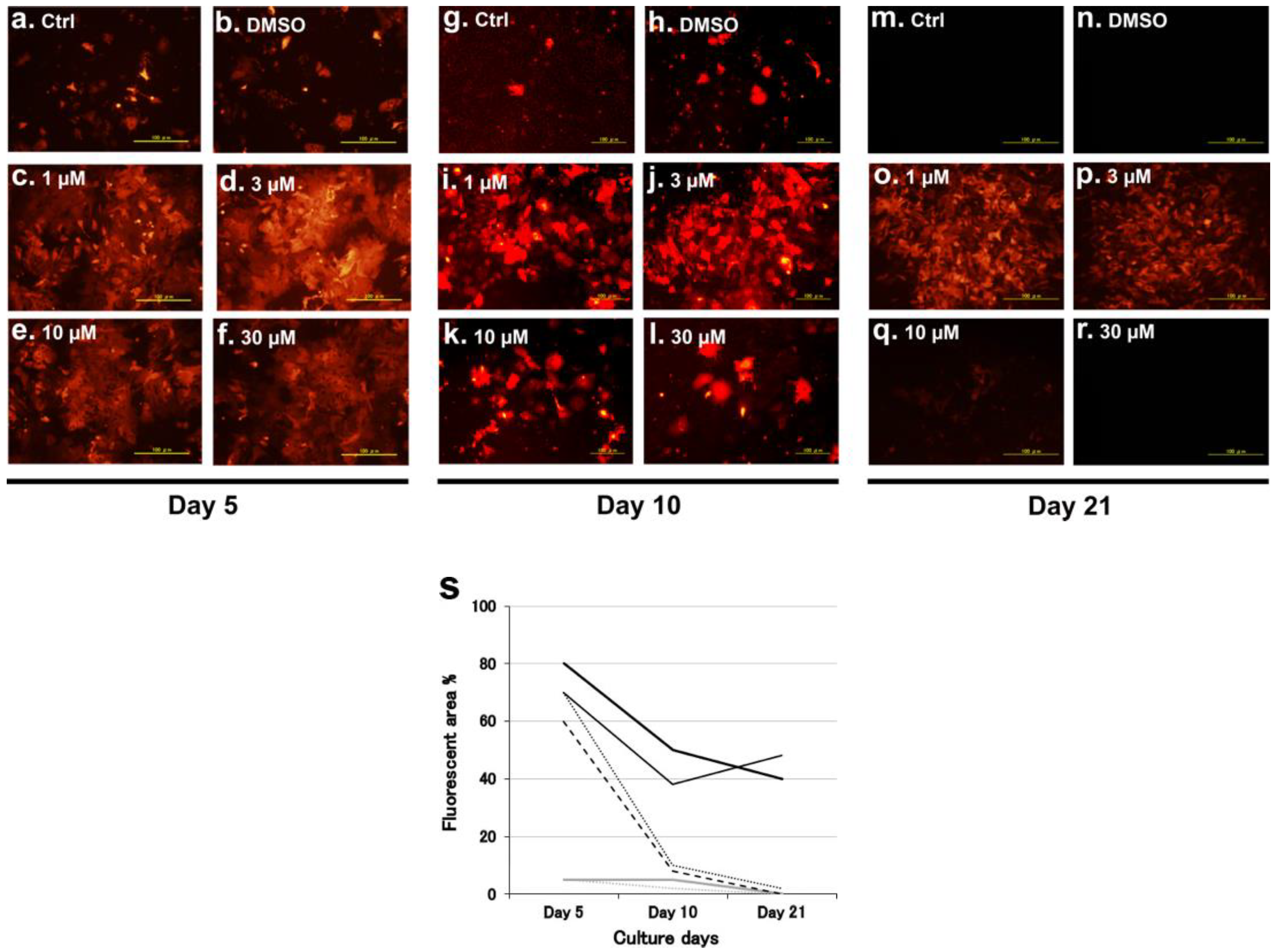{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Hepatocyte Culture
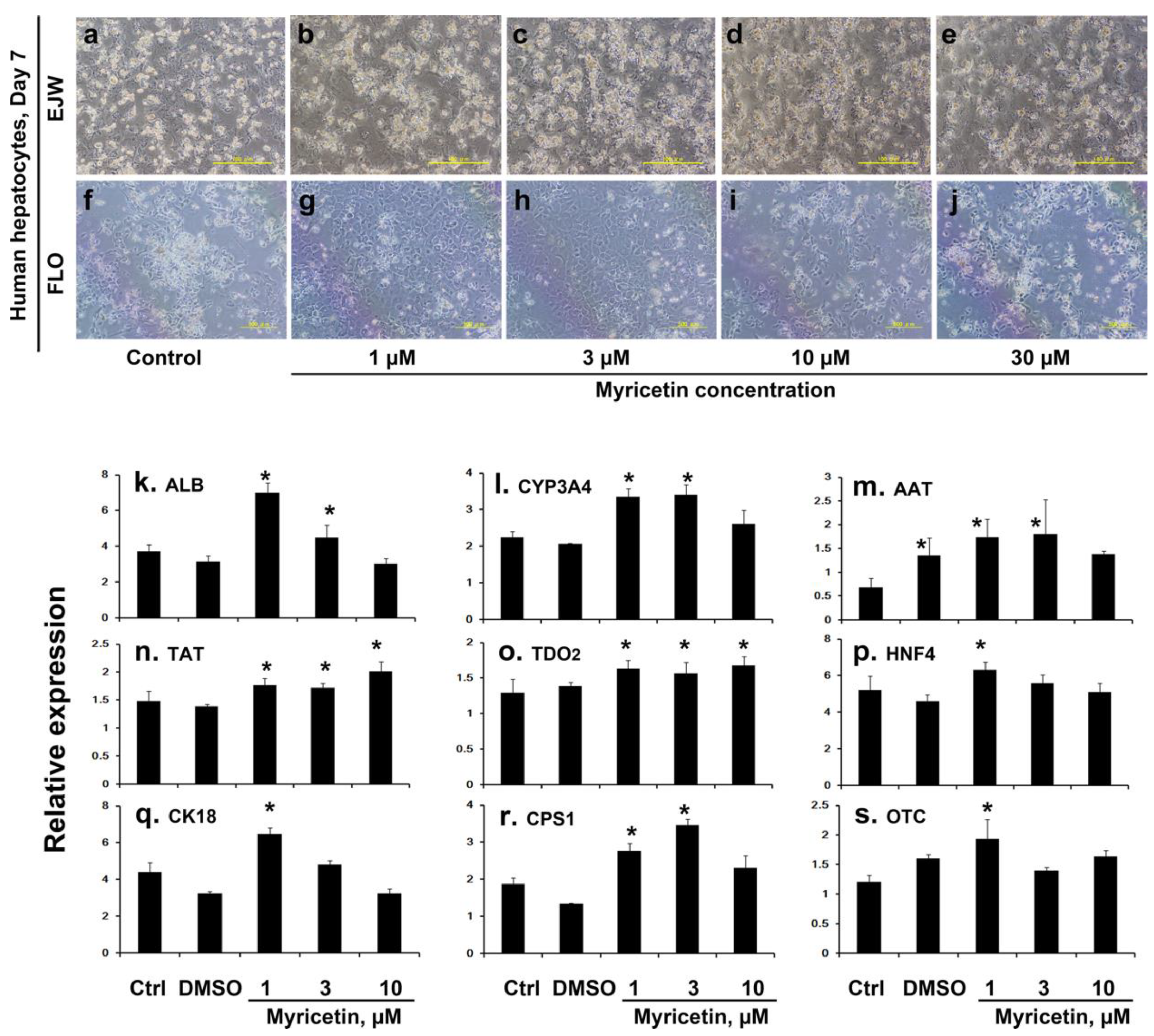
2.1.1. Porcine Hepatocyte Culture with Myricetin
2.1.2. Human Hepatocyte Culture with Myricetin
2.1.3. Ammonia Removal Activity of Myricetin Treated Hepatocytes
2.2. Hepatocyte Transplantation into Mouse Liver
2.2.1. Fate of Hepatocytes Injected Directly into the Liver of Neonatal SCID Mice
2.2.2. Survival of Human Hepatocytes in SCID Mouse Livers and Semi-Quantitative Estimation of the Advantage of Myricetin Pretreatment
2.2.3. Therapeutic Potential of Myricetin Treated Human Hepatocytes in Mutant Mice
2.3. Effect of Myricetin on the Phosphorylation of MKK4
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cells and Chemicals
4.3. Hepatocyte Culture
4.4. Hepatocyte Transplantation
4.5. Quantitative PCR Analysis
4.6. Immunohistochemical Analyses
4.7. Human Serum Albumin Assays in the Mouse Plasma
4.8. Western Blotting
4.9. Ethical Considerations
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MKK4 | Mitogen-activated protein kinase 4 |
| SCID | Severe combined immunodeficiency |
| OTC | Ornithine transcarbamylase |
| OTCD | Ornithine transcarbamylase-deficient |
| DMSO | Dimethylsulfoxide |
| JNK | c-Jun N-terminal kinase |
References
- Kim, W.R.; Lake, J.R.; Smith, J.M.; Skeans, M.A.; Schladt, D.P.; Edwards, E.B.; Harper, A.M.; Wainright, J.L.; Snyder, J.J.; Israni, A.K.; et al. OPTN/SRTR 2013 Annual Data Report: Liver. Am. J. Transplant. 2015, 15 (Suppl. 2), 1–28. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.; Karam, V.; Cailliez, V.; O Grady, J.G.; Mirza, D.; Cherqui, D.; Klempnauer, J.; Salizzoni, M.; Pratschke, J.; Jamieson, N.; et al. 2018 Annual Report of the European Liver Transplant Registry (ELTR) - 50-year evolution of liver transplantation. Transpl. Int. 2018, 31, 1293–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, R.A.; Strom, S.C. Human hepatocyte transplantation: Worldwide results. Transplantation 2006, 82, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Iansante, V.; Chandrashekran, A.; Dhawan, A. Cell-based liver therapies: Past, present and future. Philos. Trans. R. Soc. Lond B Biol. Sci. 2018, 373, 20170229. [Google Scholar] [CrossRef] [PubMed]
- Najimi, M.; Defresne, F.; Sokal, E.M. Concise Review: Updated advances and current challenges in cell therapy for inborn liver metabolic defects. Stem Cells Transl. Med. 2016, 5, 1117–1125. [Google Scholar] [CrossRef]
- Meyburg, J.; Das, A.M.; Hoerster, F.; Lindner, M.; Kriegbaum, H.; Engelmann, G.; Schmidt, J.; Ott, M.; Pettenazzo, A.; Luecke, T.; et al. One liver for four children: First clinical series of liver cell transplantation for severe neonatal urea cycle defects. Transplantation 2009, 87, 636–641. [Google Scholar] [CrossRef]
- Meyburg, J.; Hoffmann, G.F. Liver, liver cell and stem cell transplantation for the treatment of urea cycle defects. Mol. Genet. Metab. 2010, 100 (Suppl. 1), S77–S83. [Google Scholar] [CrossRef]
- Enosawa, S.; Horikawa, R.; Yamamoto, A.; Sakamoto, S.; Shigeta, T.; Nosaka, S.; Fujimoto, J.; Nakazawa, A.; Tanoue, A.; Nakamura, K.; et al. Hepatocyte transplantation using a living donor reduced graft in a baby with ornithine transcarbamylase deficiency: A novel source of hepatocytes. Liver Transplant. 2014, 20, 391–393. [Google Scholar] [CrossRef]
- Ribes-Koninckx, C.; Ibars, E.P.; Calzado Agrasot, M.Á.; Bonora-Centelles, A.; Miquel, B.P.; Vila Carbó, J.J.; Aliaga, E.D.; Pallardó, J.M.; Gómez-Lechón, M.J.; Castell, J.V. Clinical outcome of hepatocyte transplantation in four pediatric patients with inherited metabolic diseases. Cell Transplant. 2012, 21, 2267–2282. [Google Scholar] [CrossRef] [Green Version]
- Soltys, K.A.; Setoyama, K.; Tafaleng, E.N.; Soto Gutiérrez, A.; Fong, J.; Fukumitsu, K.; Nishikawa, T.; Nagaya, M.; Sada, R.; Haberman, K.; et al. Host conditioning and rejection monitoring in hepatocyte transplantation in humans. J. Hepatol. 2017, 66, 987–1000. [Google Scholar] [CrossRef] [Green Version]
- Harnly, J.M.; Doherty, R.F.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Bhagwat, S.; Gebhardt, S. Flavonoid content of U.S. fruits, vegetables, and nuts. J. Agric. Food Chem. 2006, 54, 9966–9977. [Google Scholar] [CrossRef] [PubMed]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florêncio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Shimmyo, Y.; Kihara, T.; Akaike, A.; Niidome, T.; Sugimoto, H. Three distinct neuroprotective functions of myricetin against glutamate-induced neuronal cell death: Involvement of direct inhibition of caspase-3. J. Neurosci Res. 2008, 86, 1836–1845. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.E.; Haza, A.I.; Arranz, N.; García, A.; Morales, P. Dietary polyphenols protect against N-nitrosamines and benzo(a)pyrene-induced DNA damage (strand breaks and oxidized purines/pyrimidines) in HepG2 human hepatoma cells. Eur. J. Nutr. 2008, 47, 479–490. [Google Scholar] [CrossRef]
- Kang, K.A.; Wang, Z.H.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kang, S.S.; Kim, Y.W.; Lee, J.; Park, D.; Hyun, J.W. Myricetin protects cells against oxidative stress-induced apoptosis via regulation of PI3K/Akt and MAPK signaling pathways. Int. J. Mol. Sci. 2010, 11, 4348–4360. [Google Scholar] [CrossRef]
- Zhang, K.; Ma, Z.; Wang, J.; Xie, A.; Xie, J. Myricetin attenuated MPP(+)-induced cytotoxicity by anti-oxidation and inhibition of MKK4 and JNK activation in MES23.5 cells. Neuropharmacology 2011, 61, 329–335. [Google Scholar] [CrossRef]
- Haeusgen, W.; Herdegen, T.; Waetzig, V. The bottleneck of JNK signaling: Molecular and functional characteristics of MKK4 and MKK7. Eur. J. Cell Biol. 2011, 90, 536–544. [Google Scholar] [CrossRef]
- Wuestefeld, T.; Pesic, M.; Rudalska, R.; Dauch, D.; Longerich, T.; Kang, T.W.; Yevsa, T.; Heinzmann, F.; Hoenicke, L.; Hohmeyer, A.; et al. A direct in vivo RNAi screen identifies MKK4 as a key regulator of liver regeneration. Cell 2013, 153, 389–401. [Google Scholar] [CrossRef] [Green Version]
- Matsunari, H.; Nagashima, H. Application of genetically modified and cloned pigs in translational research. J. Reprod. Dev. 2009, 55, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Shigeta, T.; Hsu, H.C.; Enosawa, S.; Matsuno, N.; Kasahara, M.; Matsunari, H.; Umeyama, K.; Watanabe, M.; Nagashima, H. Transgenic pig expressing the red fluorescent protein kusabira-orange as a novel tool for preclinical studies on hepatocyte transplantation. Transplant. Proc. 2013, 45, 1808–1810. [Google Scholar] [CrossRef]
- Matsunari, H.; Nakano, K.; Kanai, T.; Sakai, R.; Watanabe, M.; Umeyama, K.; Kobayashi, T.; Yamaguchi, T.; Shiota, A.; Nagaya, M.; et al. Generation of a double-transgenic pig with pancreas-specific green and liver-specific red fluorescence. Reprod. Fertil. Dev. 2012, 25, 311. [Google Scholar] [CrossRef]
- Cui, L.L.; Golubczyk, D.; Tolppanen, A.M.; Boltze, J.; Jolkkonen, J. Cell therapy for ischemic stroke: Are differences in preclinical and clinical study design responsible for the translational loss of efficacy? Ann. Neurol. 2019, 86, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Enosawa, S. Isolation of GMP grade human hepatocytes from remnant liver tissue of living donor liver transplantation. In Hepatocyte Transplantation: Methods and Protocols. Methods in Molecular Biology; Stock, P., Christ, B., Eds.; Springer Science+Bussiness Media: New York, NY, USA, 2017; Volume 1506, pp. 231–245. [Google Scholar]
- Jitraruch, S.; Dhawan, A.; Hughes, R.D.; Filippi, C.; Lehec, S.C.; Glover, L.; Mitry, R.R. Cryopreservation of hepatocyte microbeads for clinical transplantation. Cell Transplant. 2017, 26, 1341–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilbride, P.; Lamb, S.; Gibbons, S.; Bundy, J.; Erro, E.; Selden, C.; Fuller, B.; Morris, J. Cryopreservation and re-culture of a 2.3 litre biomass for use in a bioartificial liver device. PLoS ONE 2017, 12, e0183385. [Google Scholar] [CrossRef] [Green Version]
- De Vries, R.J.; Banik, P.D.; Nagpal, S.; Weng, L.; Ozer, S.; van Gulik, T.M.; Toner, M.; Tessier, S.N.; Uygun, K. Bulk droplet vitrification: An approach to improve large-scale hepatocyte cryopreservation outcome. Langmuir 2019, 35, 7354–7363. [Google Scholar] [CrossRef] [PubMed]
- Jitraruch, S.; Dhawan, A.; Hughes, R.D.; Filippi, C.; Soong, D.; Philippeos, C.; Lehec, S.C.; Heaton, N.D.; Longhi, M.S.; Mitry, R.R. Alginate microencapsulated hepatocytes optimised for transplantation in acute liver failure. PLoS ONE 2014, 9, e113609. [Google Scholar] [CrossRef]
- Lee, C.A.; Dhawan, A.; Iansante, V.; Lehec, S.; Khorsandi, S.E.; Filippi, C.; Walker, S.; Fernandez-Dacosta, R.; Heaton, N.; Bansal, S.; et al. Cryopreserved neonatal hepatocytes may be a source for transplantation: Evaluation of functionality toward clinical use. Liver Transplant. 2018, 24, 394–406. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.C.; Khoo, H.E. Effects of myricetin on glycemia and glycogen metabolism in diabetic rats. Life Sci. 2000, 67, 1695–1705. [Google Scholar] [CrossRef]
- Liu, I.M.; Liou, S.S.; Lan, T.W.; Hsu, F.L.; Cheng, J.T. Myricetin as the active principle of Abelmoschus moschatus to lower plasma glucose in streptozotocin-induced diabetic rats. Planta Med. 2005, 71, 617–621. [Google Scholar] [CrossRef]
- Liu, I.M.; Tzeng, T.F.; Liou, S.S.; Lan, T.W. Myricetin, a naturally occurring flavonol, ameliorates insulin resistance induced by a high-fructose diet in rats. Life Sci. 2007, 81, 1479–1488. [Google Scholar] [CrossRef]
- Kang, N.J.; Jung, S.K.; Lee, K.W.; Lee, H.J. Myricetin is a potent chemopreventive phytochemical in skin carcinogenesis. Ann. N. Y. Acad. Sci. 2011, 1229, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Lescano, C.H.; de Lima, F.F.; Mendes-Silvério, C.B.; Justo, A.F.O.; da Silva Baldivia, D.; Vieira, C.P.; Sanjinez-Argandoña, E.J.; Cardoso, C.A.L.; Mónica, F.Z.; Pires de Oliveira, I. Effect of polyphenols from campomanesia adamantium on platelet aggregation and inhibition of cyclooxygenases: Molecular docking and in vitro analysis. Front. Pharmacol. 2018, 12, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, C.; Suemizu, H.; Tamamushi, S.; Kimoto, S.; Tamaoki, N.; Ohnishi, Y. Genotyping the mouse severe combined immunodeficiency mutation using the polymerase chain reaction with confronting two-pair primers (PCR-CTPP). Exp. Anim. 2002, 51, 391–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ImageJ. Available online: https://imagej.nih.gov/ij/index.html (accessed on 16 November 2019).
- Cui, C.-H.; Uyama, T.; Miyado, K.; Terai, M.; Kyo, S.; Kiyono, T.; Umezawa, A. Menstrual blood-derived cells confer human dystrophin expression in the murine model of duchenne muscular dystrophy via cell fusion and myogenic transdifferentiation. Mol. Biol. Cell 2007, 18, 1586–1594. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, C.; Enosawa, S.; Matsunari, H.; Nagashima, H.; Umezawa, A. Natural Flavonol, Myricetin, Enhances the Function and Survival of Cryopreserved Hepatocytes In Vitro and In Vivo. Int. J. Mol. Sci. 2019, 20, 6123. https://doi.org/10.3390/ijms20246123
Cui C, Enosawa S, Matsunari H, Nagashima H, Umezawa A. Natural Flavonol, Myricetin, Enhances the Function and Survival of Cryopreserved Hepatocytes In Vitro and In Vivo. International Journal of Molecular Sciences. 2019; 20(24):6123. https://doi.org/10.3390/ijms20246123
Chicago/Turabian StyleCui, Changhao, Shin Enosawa, Hitomi Matsunari, Hiroshi Nagashima, and Akihiro Umezawa. 2019. "Natural Flavonol, Myricetin, Enhances the Function and Survival of Cryopreserved Hepatocytes In Vitro and In Vivo" International Journal of Molecular Sciences 20, no. 24: 6123. https://doi.org/10.3390/ijms20246123