In Silico Approach for the Definition of radiomiRNomic Signatures for Breast Cancer Differential Diagnosis

 , , and
, , and
Abstract
:1. Introduction
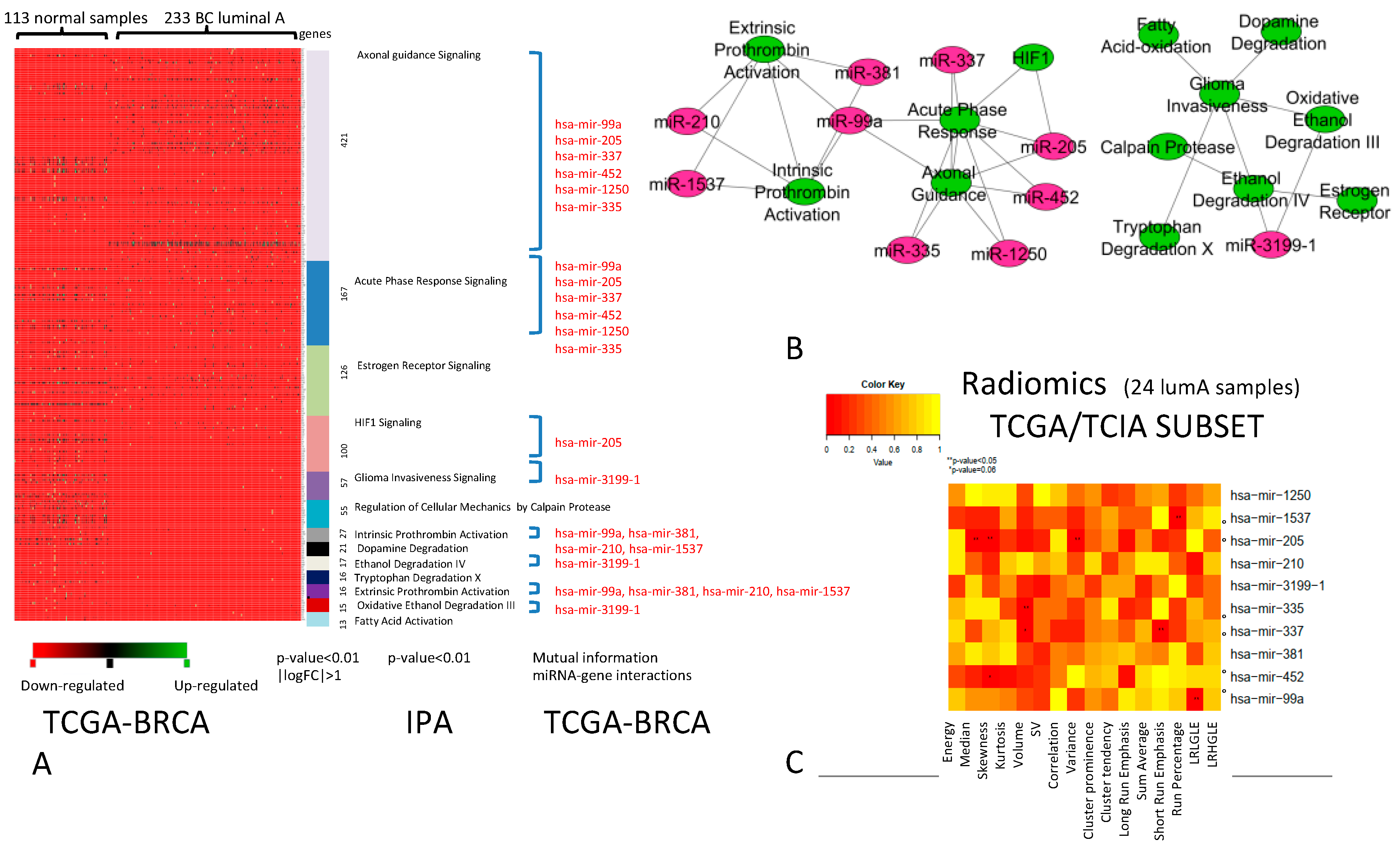
2. Results
2.1. Association Between miRNAs and Imaging Features
2.2. Diagnostic Role of RadiomiRNomic Signature
2.3. Validation of the Three miRNA Signature for the Differential Diagnosis of Human BC Samples
3. Discussion
4. Materials and Methods
4.1. Genomic and Imaging Datasets
- (1)
- TCGA-BRCA: Gene and miRNA expression levels derived by The Cancer Genome Atlas (TCGA) were downloaded including the profiles of 233 BC Luminal A, 103 BC Luminal B, 43 BC HER2+, and 74 BC basal patients, according to PAM50 classification. BC subtypes were compared with 113 normal samples, allowing the identification of the DEGs of each subtype.
- (2)
- TCGA/TCIA-BRCA_1 dataset: We used miRNA expression profiles of the TCGA-BRCA’s subset to explore the relationships between miRNAs and radiological phenotype. The subset of TCGA-BRCA (TCGA/TCIA-BRCA) was selected to include patients acquired on the same MR scanner to avoid the impact of scanner on imaging features and it contains the genomic profiles and images of 24 Luminal A, 4 Luminal B, 3 HER2+, and 6 basal samples. For these MRI samples, miRNA expression levels and PAM50 classification were available. This analysis allowed the identification of radiomic features of each BC subtype.
4.2. TCGA/TCIA-BRCA_1 Dataset: Imaging Data Analysis
4.3. TCGA-BRCA Dataset: Analysis of miRNAs Regulating Pathway Cross-Talk in BC Subtypes
4.4. TCGA/TCIA-BRCA_1 Dataset: Association Between miRNAs and IFs
4.5. TCGA/TCIA-BRCA_1, TCGA/TCIA-BRCA_2, and GSE81000 Datasets: Diagnostic Role of RadiomiRNomic Signature
4.6. Validation on BC Human Tissue Samples
Author Contributions
Funding
Conflicts of Interest
References
- Van ‘t Veer, L.J.; Dai, H.; van de Vijver, M.J.; He, Y.D.; Hart, A.A.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar] [CrossRef]
- Van de Vijver, M.J.; He, Y.D.; van‘t Veer, L.J.; Dai, H.; Hart, A.A.; Voskuil, D.W.; Schreiber, G.J.; Peterse, J.L.; Roberts, C.; Marton, M.J.; et al. A gene-expression signature as a predictor of survival in breast cancer. N. Engl. J. Med. 2002, 347, 1999–2009. [Google Scholar] [CrossRef]
- Paik, S.; Shak, S.; Tang, G.; Kim, C.; Baker, J.; Cronin, M.; Baehner, F.L.; Walker, M.G.; Watson, D.; Park, T.; et al. A multigene assay to predict recurrence of tamoxifen-treated, node-negative breast cancer. N. Engl. J. Med. 2004, 351, 2817–2826. [Google Scholar] [CrossRef]
- Paik, S.; Tang, G.; Shak, S.; Kim, C.; Baker, J.; Kim, W.; Cronin, M.; Baehner, F.L.; Watson, D.; Bryant, J.; et al. Gene expression and benefit of chemotherapy in women with node-negative, estrogen receptor-positive breast cancer. J. Clin. Oncol. 2006, 24, 3726–3734. [Google Scholar] [CrossRef]
- Cronin, M.; Sangli, C.; Liu, M.L.; Pho, M.; Dutta, D.; Nguyen, A.; Jeong, J.; Wu, J.; Langone, K.C.; Watson, D. Analytical validation of the Oncotype DX genomic diagnostic test for recurrence prognosis and therapeutic response prediction in node-negative, estrogen receptor-positive breast cancer. Clin. Chem. 2007, 53, 1084–1091. [Google Scholar] [CrossRef]
- Parker, J.S.; Mullins, M.; Cheang, M.C.; Leung, S.; Voduc, D.; Vickery, T.; Davies, S.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 2009, 27, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Parker, J.S.; Fan, C.; Perou, C.M. PAM50 assay and the three-gene model for identifying the major and clinically relevant molecular subtypes of breast cancer. Breast Cancer Res. Treat. 2012, 135, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, G.; Cava, C.; Castiglioni, I. The potential of miRNAs for diagnosis, treatment and monitoring of breast cancer. Scand. J. Clin. Lab. Investig. Suppl. 2016, 245, S34–S39. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, G.; Cava, C.; Diceglie, C.; Martelli, C.; Rizzo, G.; Piccotti, F.; Ottobrini, L.; Castiglioni, I. MicroRNA-567 dysregulation contributes to carcinogenesis of breast cancer, targeting tumor cell proliferation, and migration. Breast Cancer Res. Treat. 2017, 161, 605–616. [Google Scholar] [CrossRef]
- Castiglioni, I.; Gallivanone, F.; Soda, P.; Avanzo, M.; Stancanello, J.; Aiello, M.; Interlenghi, M.; Salvatore, M. AI-based applications in hybrid imaging: How to build smart and truly multi-parametric decision models for radiomics. Eur. J. Nucl. Med. Mol. Imaging 2019. [Google Scholar] [CrossRef]
- Aerts, H.J.; Velazquez, E.R.; Leijenaar, R.T.; Parmar, C.; Grossmann, P.; Carvalho, S.; Bussink, J.; Monshouwer, R.; Haibe-Kains, B.; Rietveld, D.; et al. Decoding tumour phenotype by noninvasive imaging using a quantitative radiomics approach. Nat. Commun. 2014, 5, 4006. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, G.; Bargallo, X.; Ganau, S.; Alonso, I.; Munoz, M.; Molla, M.; Fernandez, P.L.; Prat, A. Multiparametric MR imaging to assess response following neoadjuvant systemic treatment in various breast cancer subtypes: Comparison between different definitions of pathologic complete response. Eur. J. Radiol. 2019, 117, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Heacock, L.; Lewin, A.; Ayoola, A.; Moccaldi, M.; Babb, J.S.; Kim, S.G.; Moy, L. Dynamic Contrast-Enhanced MRI Evaluation of Pathologic Complete Response in Human Epidermal Growth Factor Receptor 2 (HER2)-Positive Breast Cancer After HER2-Targeted Therapy. Acad. Radiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Tabouret-Viaud, C.; Botsikas, D.; Delattre, B.M.; Mainta, I.; Amzalag, G.; Rager, O.; Vinh-Hung, V.; Miralbell, R.; Ratib, O. PET/MR in Breast Cancer. Semin. Nucl. Med. 2015, 45, 304–321. [Google Scholar] [CrossRef]
- Gillies, R.J.; Kinahan, P.E.; Hricak, H. Radiomics: Images Are More than Pictures, They Are Data. Radiology 2016, 278, 563–577. [Google Scholar] [CrossRef]
- Pinker, K.; Shitano, F.; Sala, E.; Do, R.K.; Young, R.J.; Wibmer, A.G.; Hricak, H.; Sutton, E.J.; Morris, E.A. Background, current role, and potential applications of radiogenomics. J. Magn. Reson. Imaging 2018, 47, 604–620. [Google Scholar] [CrossRef]
- Segal, E.; Sirlin, C.B.; Ooi, C.; Adler, A.S.; Gollub, J.; Chen, X.; Chan, B.K.; Matcuk, G.R.; Barry, C.T.; Chang, H.Y.; et al. Decoding global gene expression programs in liver cancer by noninvasive imaging. Nat. Biotechnol. 2007, 25, 675–680. [Google Scholar] [CrossRef]
- Diehn, M.; Nardini, C.; Wang, D.S.; McGovern, S.; Jayaraman, M.; Liang, Y.; Aldape, K.; Cha, S.; Kuo, M.D. Identification of noninvasive imaging surrogates for brain tumor gene-expression modules. Proc. Natl. Acad. Sci. USA 2008, 105, 5213–5218. [Google Scholar] [CrossRef]
- Gevaert, O.; Xu, J.; Hoang, C.D.; Leung, A.N.; Xu, Y.; Quon, A.; Rubin, D.L.; Napel, S.; Plevritis, S.K. Non-small cell lung cancer: Identifying prognostic imaging biomarkers by leveraging public gene expression microarray data--methods and preliminary results. Radiology 2012, 264, 387–396. [Google Scholar] [CrossRef]
- Lambin, P.; Rios-Velazquez, E.; Leijenaar, R.; Carvalho, S.; van Stiphout, R.G.; Granton, P.; Zegers, C.M.; Gillies, R.; Boellard, R.; Dekker, A.; et al. Radiomics: Extracting more information from medical images using advanced feature analysis. Eur. J. Cancer 2012, 48, 441–446. [Google Scholar] [CrossRef]
- Mahrooghy, M.; Ashraf, A.B.; Daye, D.; Mies, C.; Feldman, M.; Rosen, M.; Kontos, D. Heterogeneity wavelet kinetics from DCE-MRI for classifying gene expression based breast cancer recurrence risk. Med. Image Comput. Comput. Assist. Interv. 2013, 16, 295–302. [Google Scholar] [PubMed]
- Ha, S.; Park, S.; Bang, J.I.; Kim, E.K.; Lee, H.Y. Metabolic Radiomics for Pretreatment (18)F-FDG PET/CT to Characterize Locally Advanced Breast Cancer: Histopathologic Characteristics, Response to Neoadjuvant Chemotherapy, and Prognosis. Sci. Rep. 2017, 7, 1556. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Tamboli, P.; Vikram, R.; Rao, A. Imaging-genomic pipeline for identifying gene mutations using three-dimensional intra-tumor heterogeneity features. J. Med. Imaging (Bellingham) 2015, 2, 041009. [Google Scholar] [CrossRef] [PubMed]
- Antunovic, L.; Gallivanone, F.; Sollini, M.; Sagona, A.; Invento, A.; Manfrinato, G.; Kirienko, M.; Tinterri, C.; Chiti, A.; Castiglioni, I. [(18)F] FDG PET/CT features for the molecular characterization of primary breast tumors. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 1945–1954. [Google Scholar] [CrossRef] [PubMed]
- Leithner, D.; Bernard-Davila, B.; Martinez, D.F.; Horvat, J.V.; Jochelson, M.S.; Marino, M.A.; Avendano, D.; Ochoa-Albiztegui, R.E.; Sutton, E.J.; Morris, E.A.; et al. Radiomic Signatures Derived from Diffusion-Weighted Imaging for the Assessment of Breast Cancer Receptor Status and Molecular Subtypes. Mol. Imaging Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; He, B.; Zhuang, X.; Gao, X.; Wang, D.; Li, M.; Lin, Z.; Luo, R. CT-based radiomic signatures for prediction of pathologic complete response in esophageal squamous cell carcinoma after neoadjuvant chemoradiotherapy. J. Radiat. Res. 2019, 60, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, H.; Mofid, B.; Shiri, I.; Razzaghdoust, A.; Saadipoor, A.; Mahdavi, A.; Galandooz, H.M.; Mahdavi, S.R. Machine learning-based radiomic models to predict intensity-modulated radiation therapy response, Gleason score and stage in prostate cancer. Radiol. Med. 2019, 124, 555–567. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, Z.; Gu, D.; Tian, J.; Zhao, P.; Wei, J.; Yang, X.; Hao, X.; Dong, D.; He, N.; et al. Prediction early recurrence of hepatocellular carcinoma eligible for curative ablation using a Radiomics nomogram. Cancer Imaging 2019, 19, 21. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network; Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Mills Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Clark, K.; Vendt, B.; Smith, K.; Freymann, J.; Kirby, J.; Koppel, P.; Moore, S.; Phillips, S.; Maffitt, D.; Pringle, M.; et al. The Cancer Imaging Archive (TCIA): Maintaining and operating a public information repository. J. Digit. Imaging. 2013, 26, 1045–1057. [Google Scholar] [CrossRef]
- Pensa, S.; Watson, C.J.; Poli, V. Stat3 and the inflammation/acute phase response in involution and breast cancer. J. Mammary Gland Biol. Neoplasia 2009, 14, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Asegaonkar, S.B.; Asegaonkar, B.N.; Takalkar, U.V.; Advani, S.; Thorat, A.P. C-Reactive Protein and Breast Cancer: New Insights from Old Molecule. Int. J. Breast Cancer 2015, 2015, 145647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harburg, G.C.; Hinck, L. Navigating breast cancer: Axon guidance molecules as breast cancer tumor suppressors and oncogenes. J. Mammary Gland Biol. Neoplasia 2011, 16, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, F.; Yang, M.; Tong, N.; Fang, J.; Pan, Y.; Li, J.; Zhang, X. Identification of six key miRNAs associated with breast cancer through screening large-scale microarray data. Oncol. Lett. 2018, 16, 4159–4168. [Google Scholar] [CrossRef]
- Nguyen, B.P.; Tay, W.L.; Chui, C.K. Robust biometric recognition from palm depth images for gloved hands. IEEE Trans. Hum. Mach. Syst. 2015, 45, 799–804. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Colaprico, A.; Cava, C.; Bertoli, G.; Bontempi, G.; Castiglioni, I. Integrative Analysis with Monte Carlo Cross-Validation Reveals miRNAs Regulating Pathways Cross-Talk in Aggressive Breast Cancer. BioMed Res. Int. 2015, 2015, 831314. [Google Scholar] [CrossRef] [Green Version]
- Cava, C.; Colaprico, A.; Bertoli, G.; Bontempi, G.; Mauri, G.; Castiglioni, I. How interacting pathways are regulated by miRNAs in breast cancer subtypes. BMC Bioinform. 2016, 17, 348. [Google Scholar] [CrossRef] [Green Version]
- Meyer, D.; Dimitriadou, E.; Hornik, K.; Weingessel, A.; Leisch, F.; Chang, C.-C.; Lin, C.-C. Functions for Latent Class Analysis, Short Time Fourier Transform, Fuzzy Clustering, Support Vector Machines, Shortest Path Computation, Bagged Clustering, Naive Bayes Classifier. Available online: https://cran.r-project.org/web/packages/e1071/index.html (accessed on 5 June 2019).
- Cava, C.; Novello, C.; Martelli, C.; Lodico, A.; Ottobrsini, L.; Piccotti, F.; Truffi, M.; Corsi, F.; Bertoli, G.; Castiglioni, I. Theranostics application of miR-429 in HER2+ breast cancer. Theranostics 2019, 10, 50–61. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | AUC-TCGA/TCIA-BRCA_1 | AUC-GSE81000 |
|---|---|---|
| miR-190b | 0.92 | 0.56 |
| miR-155 | 0.88 | 0.74 |
| miR-337 | 0.87 | 0.65 |
| miR-135b | 0.73 | 0.69 |
| miR-99a | 0.72 | 0.8 |
| miR-365-2 | 0.68 | 0.55 |
| miR-335 | 0.66 | 0.45 |
| miR-452 | 0.64 | 0.51 |
| miR-429 | 0.62 | 0.54 |
| miR-190 | 0.61 | - |
| Imaging Features | AUC-TCGA/TCIA-BRCA_1 | AUC-TCGA/TCIA-BRCA_2 |
|---|---|---|
| Correlation (Corr.) | 0.84 | 0.63 |
| Short Run Emphasis (SRE) | 0.76 | 0.66 |
| Long Run High Gray Level Emphasis (LRHGLE) | 0.7 | 0.47 |
| Volume (V) | 0.6 | 0.78 |
| Sum Average (SumA) | 0.6 | 0.5 |
| miRNA | AUC-TCGA/TCIA-BRCA_1 |
|---|---|
| miR-135b; LRHGLE | 0.94 |
| miR-135b; V | 0.92 |
| miR-135b; SumA | 0.88 |
| miR-155; V | 0.86 |
| miR-135b; Corr | 0.84 |
| miR-99a; SRE | 0.82 |
| miR-155; Corr | 0.8 |
| miR-135b; SRE | 0.8 |
| miR-99a; SumA | 0.74 |
| miR-155; SumA | 0.7 |
| miR-155; LRHGLE | 0.7 |
| miRNA | AUC-TCGA/TCIA-BRCA_1 | AUC-(GSE81000) |
|---|---|---|
| miR-135b; miR-99a; SRE | 0.94 | |
| miR-135b; miR-155; miR-99a | 0.94 | 0.85 |
| miR-155; miR-99a; SRE | 0.92 | |
| miR-135b; miR-155; SRE | 0.9 | |
| miR-135b; miR-99a; Corr | 0.88 |
| Type of Feature | Feature |
|---|---|
| Histogram-based features | Energy [(g/cc)2] – [(mm2/sec)2] |
| Skewness | |
| Kurtosis | |
| Median [g/cc] – [mm2/sec] | |
| Shape-and-size based features | Volume [cm3] (V) |
| Surface to volume ratio [cm−1] (SV) | |
| Gray-Level Co-Occurrence Matrix based features (IFGLCM) | Correlation (Corr.) |
| Variance (VAR) | |
| Cluster prominence (CP) | |
| Cluster tendency (CT) | |
| Sum Average (SumA) | |
| Gray-Level Run-Length Matrix based features (IFGLRLM) | Long Run Emphasis (LRE) |
| Short Run Emphasis (SRE) | |
| Run Percentage (RP) | |
| Long Run Low Gray Level Emphasis (LRLGLE) | |
| Long Run High Gray Level Emphasis (LRHGLE) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallivanone, F.; Cava, C.; Corsi, F.; Bertoli, G.; Castiglioni, I. In Silico Approach for the Definition of radiomiRNomic Signatures for Breast Cancer Differential Diagnosis. Int. J. Mol. Sci. 2019, 20, 5825. https://doi.org/10.3390/ijms20235825
Gallivanone F, Cava C, Corsi F, Bertoli G, Castiglioni I. In Silico Approach for the Definition of radiomiRNomic Signatures for Breast Cancer Differential Diagnosis. International Journal of Molecular Sciences. 2019; 20(23):5825. https://doi.org/10.3390/ijms20235825
Chicago/Turabian StyleGallivanone, Francesca, Claudia Cava, Fabio Corsi, Gloria Bertoli, and Isabella Castiglioni. 2019. "In Silico Approach for the Definition of radiomiRNomic Signatures for Breast Cancer Differential Diagnosis" International Journal of Molecular Sciences 20, no. 23: 5825. https://doi.org/10.3390/ijms20235825