Identification of Gain and Loss of Function Missense Variants in MRGPRX2’s Transmembrane and Intracellular Domains for Mast Cell Activation by Substance P

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MRGPRX2 Mediates SP-Induced MC Activation via Both Gαi and Gαq
2.2. Mutations of the Highly Conserved Residues 3x46, 6x37, and 7x53 in MRGPRX2 Lead to a Significant Reduction in SP-Induced MC Activation
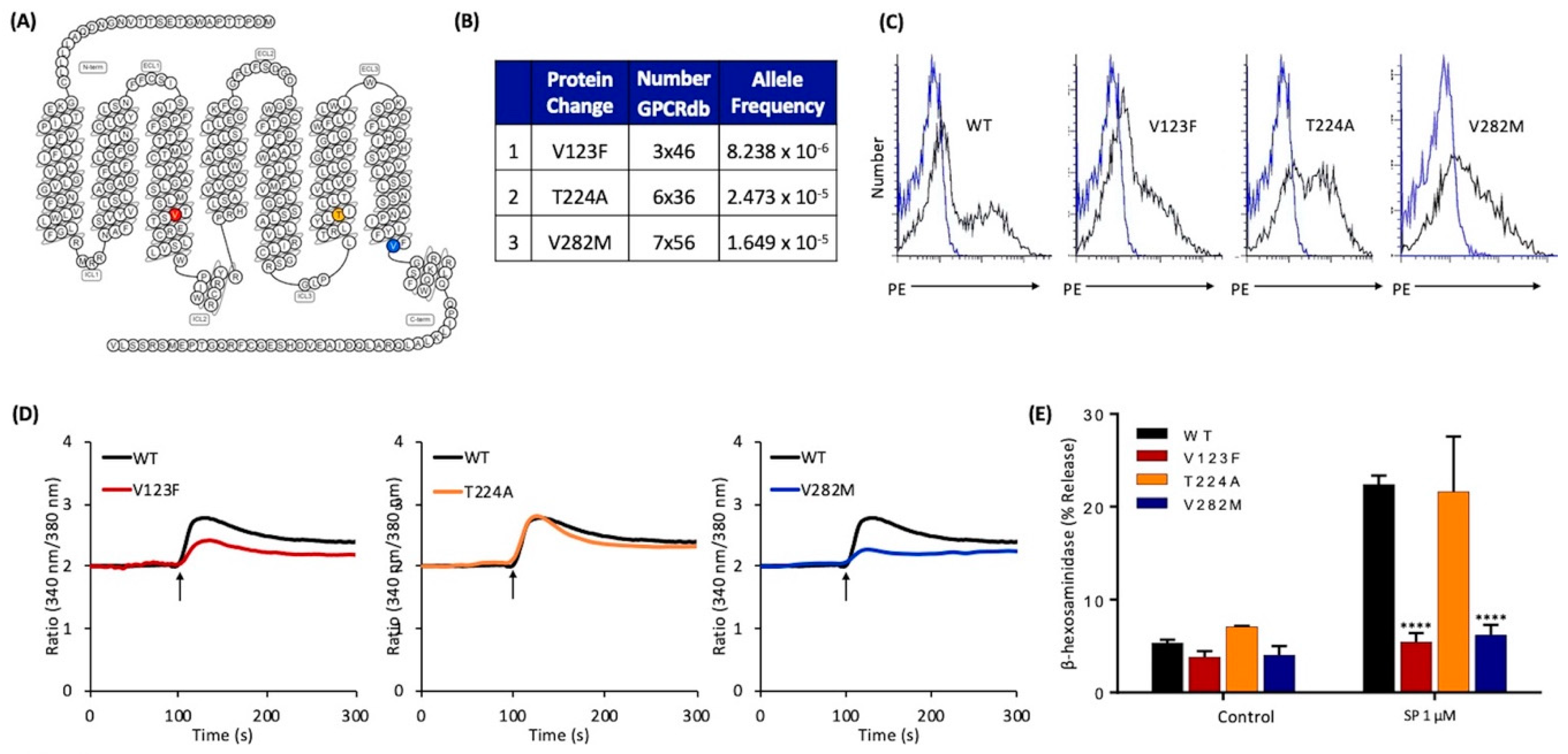
2.3. Naturally Occurring Missense MRGPRX2 Variants at or Near the Conserved Residues, V123F and V282M, Display Loss of Function Phenotype for SP-Induced MC Activation
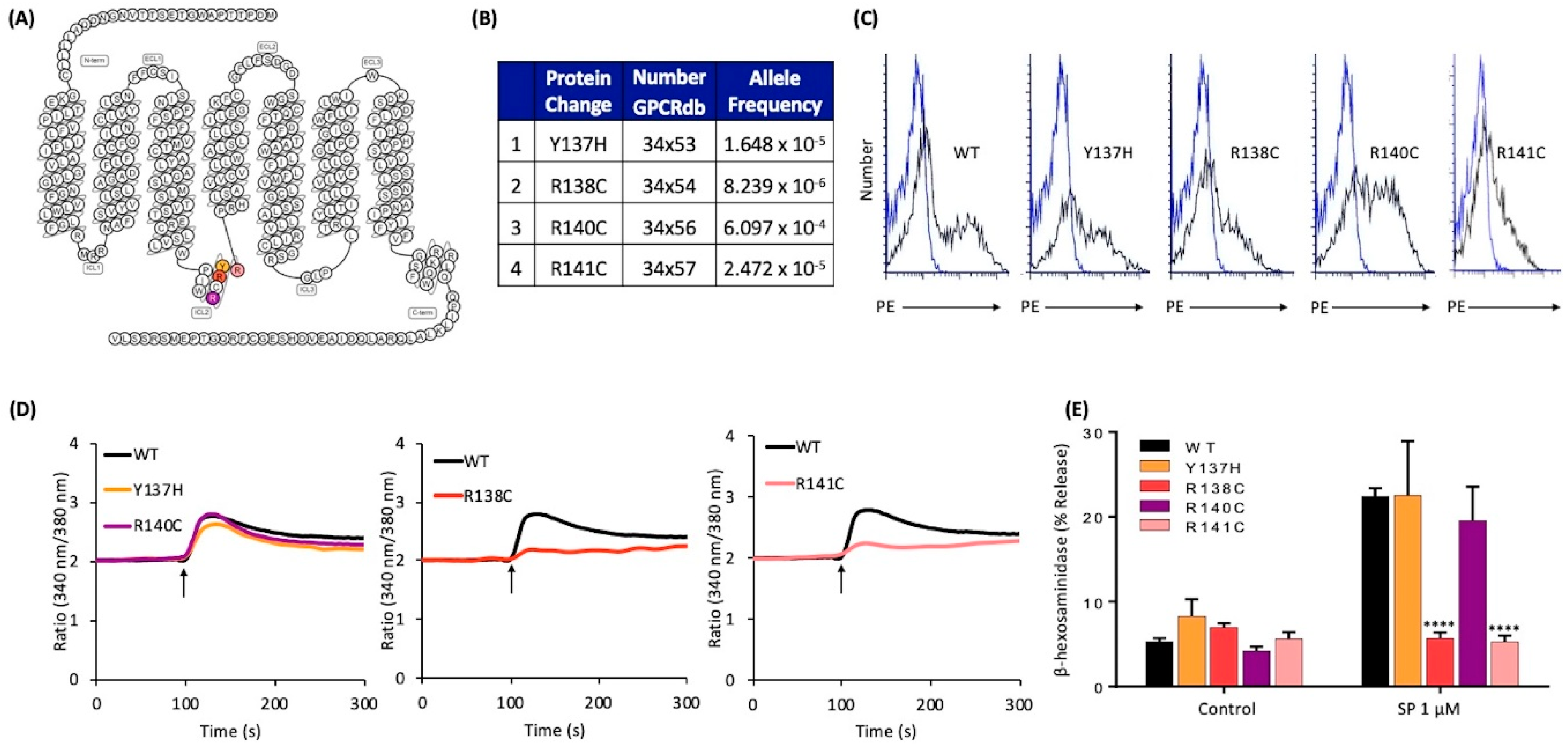
2.4. Naturally Occurring Missense MRGPRX2 Variants at the Second Intracellular Loop, R138C and R141C, Display Loss of Function Phenotype for SP-Induced MC Activation
2.5. Mutations in Potential Phosphorylation Sites of MRGPRX2 Leads to Enhanced MC Activation in Response to SP
2.6. Naturally Occurring Missense MRGPRX2 Variants at its Carboxyl-Terminus, S325L and L329Q, Display Gain of Function Phenotype for SP-Induced MC Activation
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Construction of MRGPRX2 Variants
4.4. Generation of Cells Transiently Expressing MRGPRX2 and its Variants
4.5. Degranulation Assay
4.6. Calcium Mobilization Assay
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MC | Mast cell |
| SP | Substance P |
| NK-1R | Neurokinin-1 receptor |
| MRGPRX2 | Mas-related G protein-coupled receptor-X2 |
| GPCR | G protein-coupled receptor |
| TM | Transmembrane |
| ICL | Intracellular loop |
| PTx | Pertussis toxin |
| RBL-2H3 | Rat basophilic leukemia-2H3 cells |
| RBL-MRGPRX2 | RBL-2H3 cells stably expressing MRGPRX2 |
| GPCRdb | GPCR database |
| WT | Wild-type |
| PE | Phycoerythrin |
| CRAC | Cholesterol recognition amino acid consensus |
| NHERF | Na+/H+ exchanger regulatory factor |
References
- Theoharides, T.C.; Alysandratos, K.D.; Angelidou, A.; Delivanis, D.A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast Cells and Inflammation. Biochim. Biophys. Acta 2012, 1822, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Suwanpradid, J.; Kim, I.H.; Staats, H.F.; Haniffa, M.; MacLeod, A.S.; Abraham, S.N. Perivascular Dendritic Cells Elicit Anaphylaxis by Relaying Allergens to Mast Cells Via Microvesicles. Science 2018, 362. [Google Scholar] [CrossRef] [PubMed]
- Karhausen, J.; Abraham, S.N. How Mast Cells make Decisions. J. Clin. Invest. 2016, 126, 3735–3738. [Google Scholar] [CrossRef] [PubMed]
- Kleij, H.P.; Bienenstock, J. Significance of Conversation between Mast Cells and Nerves. Allergy Asthma Clin. Immunol. 2005, 1, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsythe, P. Mast Cells in Neuroimmune Interactions. Trends Neurosci. 2019, 42, 43–55. [Google Scholar] [CrossRef]
- Gupta, K.; Harvima, I.T. Mast Cell-Neural Interactions Contribute to Pain and Itch. Immunol. Rev. 2018, 282, 168–187. [Google Scholar] [CrossRef]
- Shim, W.S.; Oh, U. Histamine-Induced Itch and its Relationship with Pain. Mol. Pain 2008, 4, 2–29. [Google Scholar] [CrossRef]
- Kulka, M.; Sheen, C.H.; Tancowny, B.P.; Grammer, L.C.; Schleimer, R.P. Neuropeptides Activate Human Mast Cell Degranulation and Chemokine Production. Immunology 2008, 123, 398–410. [Google Scholar] [CrossRef]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide Substance P and the Immune Response. Cell Mol. Life Sci. 2016, 73, 4249–4264. [Google Scholar] [CrossRef]
- Ren, K.; Dubner, R. Interactions between the Immune and Nervous Systems in Pain. Nat. Med. 2010, 16, 1267–1276. [Google Scholar] [CrossRef]
- Sagi, V.; Mittal, A.; Gupta, M.; Gupta, K. Immune Cell Neural Interactions and their Contributions to Sickle Cell Disease. Neurosci. Lett. 2019, 699, 167–171. [Google Scholar] [CrossRef]
- Serhan, N.; Basso, L.; Sibilano, R.; Petitfils, C.; Meixiong, J.; Bonnart, C.; Reber, L.L.; Marichal, T.; Starkl, P.; Cenac, N.; et al. House Dust Mites Activate Nociceptor-Mast Cell Clusters to Drive Type 2 Skin Inflammation. Nat. Immunol. 2019. [CrossRef] [PubMed]
- Fujisawa, D.; Kashiwakura, J.; Kita, H.; Kikukawa, Y.; Fujitani, Y.; Sasaki-Sakamoto, T.; Kuroda, K.; Nunomura, S.; Hayama, K.; Terui, T.; et al. Expression of Mas-Related Gene X2 on Mast Cells is Upregulated in the Skin of Patients with Severe Chronic Urticaria. J. Allergy Clin. Immunol. 2014, 134, 62–633.e9. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Recio, S.; Gascon, P. Biological and Pharmacological Aspects of the NK1-Receptor. Biomed. Res. Int. 2015, 2015, 495704. [Google Scholar]
- Borsook, D.; Upadhyay, J.; Klimas, M.; Schwarz, A.J.; Coimbra, A.; Baumgartner, R.; George, E.; Potter, W.Z.; Large, T.; Bleakman, D.; et al. Decision-Making using fMRI in Clinical Drug Development: Revisiting NK-1 Receptor Antagonists for Pain. Drug Discov. Today 2012, 17, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Tatemoto, K.; Nozaki, Y.; Tsuda, R.; Konno, S.; Tomura, K.; Furuno, M.; Ogasawara, H.; Edamura, K.; Takagi, H.; Iwamura, H.; et al. Immunoglobulin E-Independent Activation of Mast Cell is Mediated by Mrg Receptors. Biochem. Biophys. Res. Commun. 2006, 349, 1322–1328. [Google Scholar]
- McNeil, B.D.; Pundir, P.; Meeker, S.; Han, L.; Undem, B.J.; Kulka, M.; Dong, X. Identification of a Mast-Cell-Specific Receptor Crucial for Pseudo-Allergic Drug Reactions. Nature 2015, 519, 237–241. [Google Scholar] [CrossRef]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A Mast-Cell-Specific Receptor Mediates Neurogenic Inflammation and Pain. Neuron 2019, 101, 41–420.e3. [Google Scholar] [CrossRef]
- Navratilova, E.; Porreca, F. Substance P and Inflammatory Pain: Getting it Wrong and Right Simultaneously. Neuron 2019, 101, 353–355. [Google Scholar] [CrossRef]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Diversity and Modularity of G Protein-Coupled Receptor Structures. Trends Pharmacol. Sci. 2012, 33, 17–27. [Google Scholar] [CrossRef]
- Venkatakrishnan, A.J.; Deupi, X.; Lebon, G.; Tate, C.G.; Schertler, G.F.; Babu, M.M. Molecular Signatures of G-Protein-Coupled Receptors. Nature 2013, 494, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, A.J.; Deupi, X.; Lebon, G.; Heydenreich, F.M.; Flock, T.; Miljus, T.; Balaji, S.; Bouvier, M.; Veprintsev, D.B.; Tate, C.G.; et al. Diverse Activation Pathways in Class A GPCRs Converge Near the G-Protein-Coupling Region. Nature 2016, 536, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Isberg, V.; de Graaf, C.; Bortolato, A.; Cherezov, V.; Katritch, V.; Marshall, F.H.; Mordalski, S.; Pin, J.P.; Stevens, R.C.; Vriend, G.; et al. Generic GPCR Residue Numbers-Aligning Topology Maps while Minding the Gaps. Trends Pharmacol. Sci. 2015, 36, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Munk, C.; Isberg, V.; Mordalski, S.; Harpsoe, K.; Rataj, K.; Hauser, A.S.; Kolb, P.; Bojarski, A.J.; Vriend, G.; Gloriam, D.E. GPCRdb: The G Protein-Coupled Receptor Database-an Introduction. Br. J. Pharmacol. 2016, 173, 2195–2207. [Google Scholar] [CrossRef]
- Wootten, D.; Christopoulos, A.; Marti-Solano, M.; Babu, M.M.; Sexton, P.M. Mechanisms of Signalling and Biased Agonism in G Protein-Coupled Receptors. Nat. Rev. Mol. Cell Biol. 2018, 19, 638–653. [Google Scholar]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and Dynamics of GPCR Signaling Complexes. Nat. Struct. Mol. Biol. 2018, 25, 4–12. [Google Scholar] [CrossRef]
- Rasmussen, S.G.; DeVree, B.T.; Zou, Y.; Kruse, A.C.; Chung, K.Y.; Kobilka, T.S.; Thian, F.S.; Chae, P.S.; Pardon, E.; Calinski, D.; et al. Crystal Structure of the Beta2 Adrenergic Receptor-Gs Protein Complex. Nature 2011, 477, 549–555. [Google Scholar] [CrossRef]
- Gupta, K.; Subramanian, H.; Klos, A.; Ali, H. Phosphorylation of C3a Receptor at Multiple Sites Mediates Desensitization, Beta-Arrestin-2 Recruitment and Inhibition of NF-kappaB Activity in Mast Cells. PLoS ONE 2012, 7, e46369. [Google Scholar] [CrossRef]
- Guo, Q.; Subramanian, H.; Gupta, K.; Ali, H. Regulation of C3a Receptor Signaling in Human Mast Cells by G Protein Coupled Receptor Kinases. PLoS ONE 2011, 6, e22559. [Google Scholar] [CrossRef]
- Cahill, T.J., 3rd; Thomsen, A.R.; Tarrasch, J.T.; Plouffe, B.; Nguyen, A.H.; Yang, F.; Huang, L.Y.; Kahsai, A.W.; Bassoni, D.L.; Gavino, B.J.; et al. Distinct Conformations of GPCR-Beta-Arrestin Complexes Mediate Desensitization, Signaling, and Endocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, 2562–2567. [Google Scholar] [CrossRef]
- Gurevich, V.V.; Gurevich, E.V. GPCR Signaling Regulation: The Role of GRKs and Arrestins. Front. Pharmacol. 2019, 10, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Ganguly, A.; Haque, M.; Ali, H. Angiogenic Host Defense Peptide AG-30/5C and Bradykinin B2 Receptor Antagonist Icatibant are G Protein Biased Agonists for MRGPRX2 in Mast Cells. J. Immunol. 2019, 202, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, H.; Gupta, K.; Guo, Q.; Price, R.; Ali, H. Mas-Related Gene X2 (MrgX2) is a Novel G Protein-Coupled Receptor for the Antimicrobial Peptide LL-37 in Human Mast Cells: Resistance to Receptor Phosphorylation, Desensitization, and Internalization. J. Biol. Chem. 2011, 286, 44739–44749. [Google Scholar] [CrossRef] [PubMed]
- Lansu, K.; Karpiak, J.; Liu, J.; Huang, X.P.; McCorvy, J.D.; Kroeze, W.K.; Che, T.; Nagase, H.; Carroll, F.I.; Jin, J.; et al. In Silico Design of Novel Probes for the Atypical Opioid Receptor MRGPRX2. Nat. Chem. Biol. 2017, 13, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.B.; Graham, T.A.; Azimi, E.; Lerner, E.A. A Single Amino Acid in MRGPRX2 Necessary for Binding and Activation by Pruritogens. J. Allergy Clin. Immunol. 2017, 140, 1726–1728. [Google Scholar] [CrossRef]
- Alkanfari, I.; Gupta, K.; Jahan, T.; Ali, H. Naturally Occurring Missense MRGPRX2 Variants Display Loss of Function Phenotype for Mast Cell Degranulation in Response to Substance P, Hemokinin-1, Human Beta-Defensin-3, and Icatibant. J. Immunol. 2018, 201, 343–349. [Google Scholar] [CrossRef]
- Subramanian, H.; Gupta, K.; Lee, D.; Bayir, A.K.; Ahn, H.; Ali, H. Beta-Defensins Activate Human Mast Cells Via Mas-Related Gene X2. J. Immunol. 2013, 191, 345–352. [Google Scholar] [CrossRef]
- Takasaki, J.; Saito, T.; Taniguchi, M.; Kawasaki, T.; Moritani, Y.; Hayashi, K.; Kobori, M. A Novel Galphaq/11-Selective Inhibitor. J. Biol. Chem. 2004, 279, 47438–47445. [Google Scholar] [CrossRef]
- Gupta, K.; Subramanian, H.; Ali, H. Modulation of Host Defense Peptide-Mediated Human Mast Cell Activation by LPS. Innate Immun. 2016, 22, 21–30. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, B.; Feng, D.; Hu, H.; Chu, M.; Qu, Q.; Tarrasch, J.T.; Li, S.; Sun Kobilka, T.; Kobilka, B.K.; et al. Cryo-EM Structure of the Activated GLP-1 Receptor in Complex with a G Protein. Nature 2017, 546, 248–253. [Google Scholar] [CrossRef]
- Meixiong, J.; Anderson, M.; Limjunyawong, N.; Sabbagh, M.F.; Hu, E.; Mack, M.R.; Oetjen, L.K.; Wang, F.; Kim, B.S.; Dong, X. Activation of Mast-Cell-Expressed Mas-Related G-Protein-Coupled Receptors Drives Non-Histaminergic Itch. Immunity 2019, 50, 116–1171.e5. [Google Scholar] [CrossRef]
- Pundir, P.; Liu, R.; Vasavda, C.; Serhan, N.; Limjunyawong, N.; Yee, R.; Zhan, Y.; Dong, X.; Wu, X.; Zhang, Y.; et al. A Connective Tissue Mast-Cell-Specific Receptor Detects Bacterial Quorum-Sensing Molecules and Mediates Antibacterial Immunity. Cell. Host Microbe. 2019, 26, 11–122.e8. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, H.; Gupta, K.; Ali, H. Roles of Mas-Related G Protein-Coupled Receptor X2 on Mast Cell-Mediated Host Defense, Pseudoallergic Drug Reactions, and Chronic Inflammatory Diseases. J. Allergy Clin. Immunol. 2016, 138, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Civciristov, S.; Ellisdon, A.M.; Suderman, R.; Pon, C.K.; Evans, B.A.; Kleifeld, O.; Charlton, S.J.; Hlavacek, W.S.; Canals, M.; Halls, M.L. Preassembled GPCR Signaling Complexes Mediate Distinct Cellular Responses to Ultralow Ligand Concentrations. Sci. Signal. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Moro, O.; Lameh, J.; Hogger, P.; Sadee, W. Hydrophobic Amino Acid in the I2 Loop Plays a Key Role in Receptor-G Protein Coupling. J. Biol. Chem. 1993, 268, 22273–22276. [Google Scholar] [PubMed]
- Villar, V.A.; Cuevas, S.; Zheng, X.; Jose, P.A. Localization and Signaling of GPCRs in Lipid Rafts. Methods Cell Biol. 2016, 132, 3–23. [Google Scholar]
- Nobles, K.N.; Xiao, K.; Ahn, S.; Shukla, A.K.; Lam, C.M.; Rajagopal, S.; Strachan, R.T.; Huang, T.Y.; Bressler, E.A.; Hara, M.R.; et al. Distinct Phosphorylation Sites on the Beta(2)-Adrenergic Receptor Establish a Barcode that Encodes Differential Functions of Beta-Arrestin. Sci. Signal. 2011, 4, ra51. [Google Scholar] [CrossRef]
- Ardura, J.A.; Friedman, P.A. Regulation of G Protein-Coupled Receptor Function by Na+/H+ Exchange Regulatory Factors. Pharmacol. Rev. 2011, 63, 882–900. [Google Scholar] [CrossRef]
- Wang, B.; Yang, Y.; Abou-Samra, A.B.; Friedman, P.A. NHERF1 Regulates Parathyroid Hormone Receptor Desensitization: Interference with Beta-Arrestin Binding. Mol. Pharmacol. 2009, 75, 1189–1197. [Google Scholar] [CrossRef]
- Subramanian, H.; Gupta, K.; Ali, H. Roles for NHERF1 and NHERF2 on the Regulation of C3a Receptor Signaling in Human Mast Cells. PLoS ONE 2012, 7, e51355. [Google Scholar] [CrossRef]
- Ali, H.; Richardson, R.M.; Tomhave, E.D.; DuBose, R.A.; Haribabu, B.; Snyderman, R. Regulation of Stably Transfected Platelet Activating Factor Receptor in RBL-2H3 Cells. Role of Multiple G Proteins and Receptor Phosphorylation. J. Biol. Chem. 1994, 269, 24557–24563. [Google Scholar] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chompunud Na Ayudhya, C.; Roy, S.; Alkanfari, I.; Ganguly, A.; Ali, H. Identification of Gain and Loss of Function Missense Variants in MRGPRX2’s Transmembrane and Intracellular Domains for Mast Cell Activation by Substance P. Int. J. Mol. Sci. 2019, 20, 5247. https://doi.org/10.3390/ijms20215247
Chompunud Na Ayudhya C, Roy S, Alkanfari I, Ganguly A, Ali H. Identification of Gain and Loss of Function Missense Variants in MRGPRX2’s Transmembrane and Intracellular Domains for Mast Cell Activation by Substance P. International Journal of Molecular Sciences. 2019; 20(21):5247. https://doi.org/10.3390/ijms20215247
Chicago/Turabian StyleChompunud Na Ayudhya, Chalatip, Saptarshi Roy, Ibrahim Alkanfari, Anirban Ganguly, and Hydar Ali. 2019. "Identification of Gain and Loss of Function Missense Variants in MRGPRX2’s Transmembrane and Intracellular Domains for Mast Cell Activation by Substance P" International Journal of Molecular Sciences 20, no. 21: 5247. https://doi.org/10.3390/ijms20215247