Expressing Double-Stranded RNAs of Insect Hormone-Related Genes Enhances Baculovirus Insecticidal Activity

Abstract
:1. Introduction
2. Results
2.1. Construction of Recombinant Bacmid Containing dsRNA Expression Cassette
2.2. Detection of Recombinant Baculoviruses in Transfected Cells
2.3. Efficacy of Recombinant Helicoverpa Armigera Nucleopolyhedroviruses in Killing H. armigera Larvae
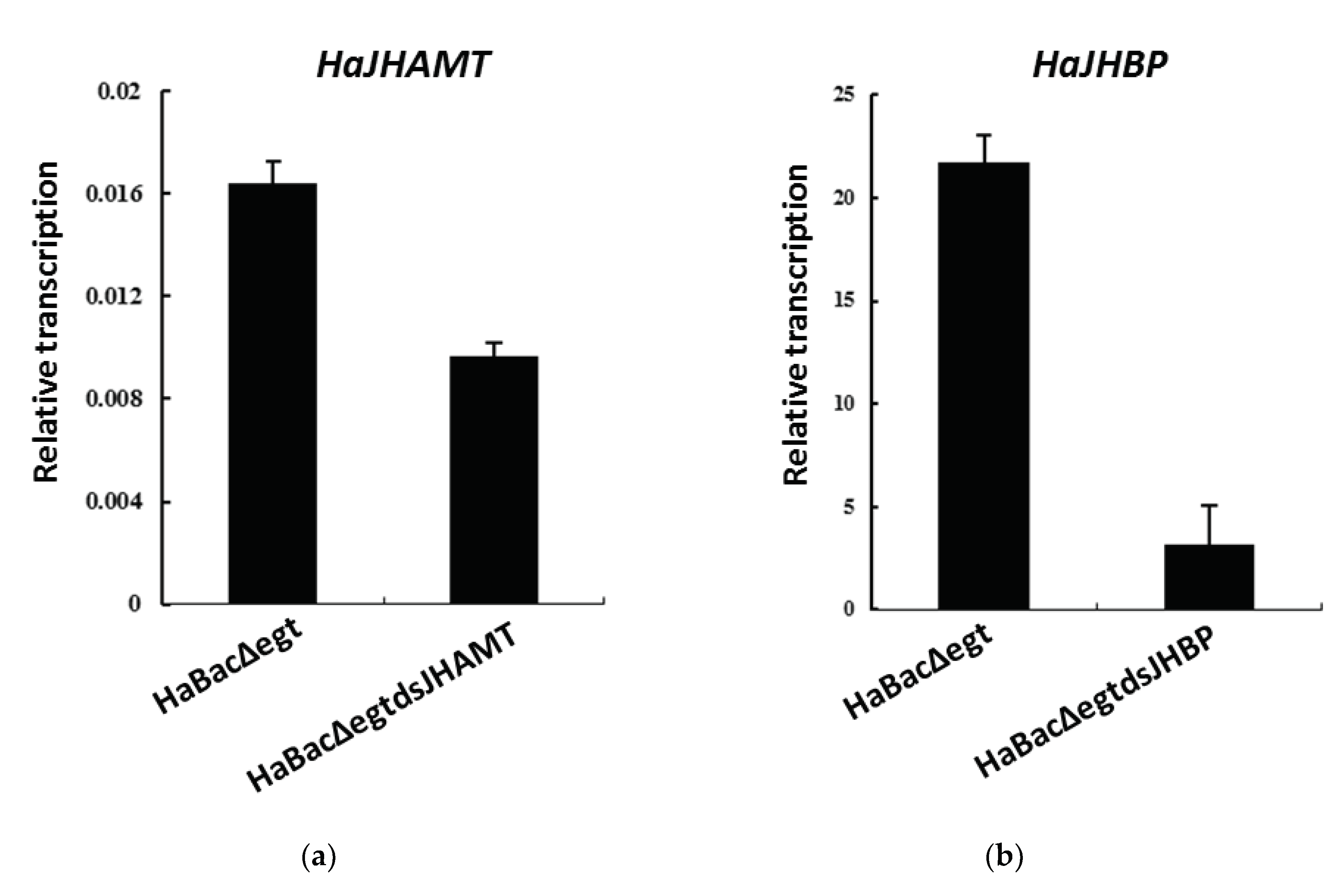
2.4. Downregulation of RNA Interference Target Gene Expression in Infected H. armigera Larvae
2.5. Effect on Hormone-Related Downstream Genes in Infected H. armigera Larvae
3. Discussion
4. Materials and Methods
4.1. Insects, Cell Line, and Viruses
4.2. Construction of Donor Plasmids
4.3. Construction of Recombinant Bacmids
4.4. Generation of Budded Viruses
4.5. Detection of dsRNA Transcripts in Infected Cells
4.6. Purification of Viral Occlusion Bodies
4.7. Transcription Analysis of Genes in Infected H. armigera Larvae
4.8. Bioassays
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| bacmid | baculovirus plasmid |
| Bt | Bacillus thuringiensis |
| eGFP | enhanced green fluorescence protein gene |
| GmR | gentamicin resistance gene |
| CmR | chloramphenicol resistance gene |
| TCID50 | 50% tissue culture infective dose |
| PIB | polyhedral inclusion body |
| UDP | uridine diphosphate |
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Beascatena, A.; Sánchezmirón, A.; Garcíacamacho, F.; Contrerasgómez, A.; Molinagrima, E. Baculovirus biopesticides: An overview. J. Anim. Plant Sci. 2014, 24, 362–373. [Google Scholar]
- Douglas, A.E. Strategies for enhanced crop resistance to insect pests. Annu. Rev. Plant Biol. 2018, 69, 637–660. [Google Scholar] [CrossRef]
- Armenta, R.; Martínez, A.M.; Chapman, J.W.; Magallanes, R.; Goulson, D.; Caballero, P.; Cave, R.D.; Cisneros, J.; Valle, J.; Castillejos, V.; et al. Impact of a nucleopolyhedrovirus bioinsecticide and selected synthetic insecticides on the abundance of insect natural enemies on maize in southern Mexico. J. Econ. Entomol. 2003, 96, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Makkonen, K.E.; Airenne, K.; Yla-Herttulala, S. Baculovirus-mediated gene delivery and RNAi applications. Viruses 2015, 7, 2099–2125. [Google Scholar] [CrossRef] [PubMed]
- Fuller, E.; Elderd, B.D.; Dwyer, G. Pathogen Persistence in the Environment and Insect-Baculovirus Interactions: Disease-Density Thresholds, Epidemic Burnout, and Insect Outbreaks. Am. Nat. 2012, 179, E70–E96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus insecticides in Latin America: Historical overview, current status and future perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef]
- Szewczyk, B.; Hoyos-Carvajal, L.; Paluszek, M.; Skrzecz, I.; Souza, M.L.D. Baculoviruses—Re-emerging biopesticides. Biotechnol. Adv. 2006, 24, 143–160. [Google Scholar] [CrossRef]
- Moscardi, F. Assessment of the application of baculoviruses for control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Stewart, L.M.D.; Hirst, M.; Ferber, M.L.P.; Merryweather, A.T.; Cayley, P.J.; Possee, R.D. Construction of an improved baculovirus insecticide containing an insect-specific toxin gene. Nature 1991, 352, 85–88. [Google Scholar] [CrossRef]
- Tomalski, M.D.; Miller, L.K. Insect paralysis by baculovirus-mediated expression of a mite neurotoxin gene. Nature 1991, 352, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Paul Pener, M. Insect Growth Regulators. In Encyclopedia of Pest Management; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar] [CrossRef]
- Maeda, S. Increased insecticidal effect by a recombinant baculovirus carrying a synthetic diuretic hormone gene. Biochem. Biophys. Res. Commun. 1989, 165, 1177–1183. [Google Scholar] [CrossRef]
- Eldridge, R.; Horodyski, F.M.; Morton, D.B.; O’Reilly, D.R.; Truman, J.W.; Riddiford, L.M.; Miller, L.K. Expression of an eclosion hormone gene in insect cells using baculovirus vectors. Insect Biochem. 1991, 21, 341–351. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Kelly, T.J.; Masler, E.P.; Thyagaraja, B.S.; Moy Robson, R.; Shaw, T.C.; Miller, L.K. Overexpression of Bombyx mori prothoracicotropic hormone using baculovirus vectors. Insect Biochem. Mol. Biol. 1995, 25, 475–485. [Google Scholar] [CrossRef]
- Hammock, B.D.; Bonning, B.C.; Possee, R.D.; Hanzlik, T.N.; Maeda, S. Expression and effects of the juvenile hormone esterase in a baculovirus vector. Nature 1990, 344, 458–461. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Miller, L.K. Improvement of a baculovirus pesticide by deletion of the EGT gene. Nat. Biotechnol. 1991, 9, 1086–1089. [Google Scholar] [CrossRef]
- O’Reilly, D.R. Baculovirus-encoded ecdysteroid UDP-glucosyltransferases. Insect Biochem. Mol. Biol. 1995, 25, 541–550. [Google Scholar] [CrossRef]
- Mello, C.C.; Conte, D. Revealing the world of RNA interference. Nature 2004, 431, 338–342. [Google Scholar] [CrossRef]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Kolliopoulou, A.; Swevers, L. Recent progress in RNAi research in Lepidoptera: Intracellular machinery, antiviral immune response and prospects for insect pest control. Curr. Opin. Insect Sci. 2014, 6, 28–34. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Ma, W.; Wang, X.; Gao, M.; Dai, Y.; Wei, X.; Zhang, L.; Peng, Y.; Chen, S.; Ding, L.; et al. Next-generation transgenic cotton: Pyramiding RNAi and Bt counters insect resistance. Plant Biotechnol. J. 2017, 15, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Hitchon, A. Environmental safety assessment considerations for plants expressing novel insecticidal RNAi traits in Canada. In Proceedings of the 2016 International Congress of Entomology, Orlando, FL, USA, 25–30 September 2016. [Google Scholar]
- Kolliopoulou, A.; Taning, C.N.T.; Smagghe, G.; Swevers, L. Viral delivery of dsRNA for control of insect agricultural pests and vectors of human disease: Prospects and challenges. Front. Physiol. 2017, 8, 399. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.T.; Soria, M.F.; Walsh, T.; Thomazoni, D.; Silvie, P.; Behere, G.T.; Anderson, C.; Downes, S. A brave new world for an old world pest: Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. PLoS ONE 2013, 8, e80134. [Google Scholar] [CrossRef] [PubMed]
- Popham, H.J.R.; Nusawardani, T.; Bonning, B.C. Introduction to the Use of Baculoviruses as Biological Insecticides, 3rd ed.; Murhammer, D.W., Ed.; Springer: New York, NY, USA, 2016; pp. 383–392. [Google Scholar] [CrossRef]
- Huang, Y.; Deng, F.; Hu, Z.; Vlak, J.; Wang, H. Baculovirus-mediated gene silencing in insect cells using intracellularly produced long double-stranded RNA. J. Biotechnol. 2007, 128, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Kontogiannatos, D.; Swevers, L.; Maenaka, K.; Park, E.Y.; Iatrou, K.; Kourti, A. Functional characterization of a juvenile hormone esterase related gene in the moth Sesamia nonagrioides through RNA interference. PLoS ONE 2013, 8, e73834. [Google Scholar] [CrossRef]
- Jindra, M.; Palli, S.R.; Riddiford, L.M. The juvenile hormone signaling pathway in insect development. Annu. Rev. Entomol. 2013, 58, 181–204. [Google Scholar] [CrossRef]
- Bellés, X.; Martín, D.; Piulachs, M.D. The mevalonate pathway and the synthesis of juvenile hormone in insects. Annu. Rev. Entomol. 2005, 50, 181–199. [Google Scholar] [CrossRef]
- Minakuchi, C.; Namiki, T.; Yoshiyama, M.; Shinoda, T. RNAi-mediated knockdown of juvenile hormone acid O-methyltransferase gene causes precocious metamorphosis in the red flour beetle Tribolium castaneum. FEBS J. 2008, 275, 2919–2931. [Google Scholar] [CrossRef]
- Trowell, S.C. High affinity juvenile hormone carrier proteins in the haemolymph of insects. Comp. Biochem. Physiol. Part B Comp. Biochem. 1992, 103, 795–807. [Google Scholar] [CrossRef]
- Hidayat, P.; Goodman, W.G. Juvenile hormone and hemolymph juvenile hormone binding protein titers and their interaction in the hemolymph of fourth stadium Manduca sexta. Insect Biochem. Mol. Biol. 1994, 24, 709–715. [Google Scholar] [CrossRef]
- Suzuki, R.; Fujimoto, Z.; Shiotsuki, T.; Tsuchiya, W.; Momma, M.; Tase, A.; Miyazawa, M.; Yamazaki, T. Structural mechanism of JH delivery in hemolymph by JHBP of silkworm, Bombyx mori. Sci. Rep. 2011, 1, 133. [Google Scholar] [CrossRef] [PubMed]
- El-Sheikh, E.-S.A.; Kamita, S.G.; Vu, K.; Hammock, B.D. Improved insecticidal efficacy of a recombinant baculovirus expressing mutated JH esterase from Manduca sexta. Biol. Control 2011, 58, 354–361. [Google Scholar] [CrossRef]
- Kamita, S.G.; Hammock, B.D. Juvenile hormone esterase: Biochemistry and structure. J. Pest. Sci. 2010, 35, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Inceoglu, A.B.; Kamita, S.G.; Hammock, B.D. Genetically modified baculoviruses: A historical overview and future outlook. Adv. Virus Res. 2006, 68, 323–360. [Google Scholar] [CrossRef]
- Chang, J.H.; Choi, J.Y.; Jin, B.R.; Roh, J.Y.; Olszewski, J.A.; Seo, S.J.; O’Reilly, D.R.; Je, Y.H. An improved baculovirus insecticide producing occlusion bodies that contain Bacillus thuringiensis insect toxin. J. Invertebr. Pathol. 2003, 84, 30–37. [Google Scholar] [CrossRef]
- Georgievska, L.; Joosten, N.; Hoover, K.; Cory, J.S.; Vlak, J.M.; van der Werf, W. Effects of single and mixed infections with wild type and genetically modified Helicoverpa armigera nucleopolyhedrovirus on movement behaviour of cotton bollworm larvae. Entomol. Exp. Appl. 2010, 135, 56–67. [Google Scholar] [CrossRef]
- Si, Y.-H.; Fang, M.-G.; Huang, Y.; Zheng, F.-L.; Li, T.; Hu, Z.-H.; Wang, H.-Z. Construction and characterization of a Helicoverpa armigera nucleopolyhedrovirus bacterial artificial chromosome with deletion of ecdysteroid UDP-glucosyltransferase gene. Biosci. Biotechnol. Biochem. 2007, 71, 2435–2441. [Google Scholar] [CrossRef]
- McIntosh, A.H.; Ignoffo, C.M. Characterization of five cell lines established from species of Heliothis. Appl. Entomol. Zool. 1983, 18, 262–269. [Google Scholar] [CrossRef]
- Wang, H.; Deng, F.; Pijlman, G.P.; Chen, X.; Sun, X.; Vlak, J.M.; Hu, Z. Cloning of biologically active genomes from a Helicoverpa armigera single-nucleocapsid nucleopolyhedrovirus isolate by using a bacterial artificial chromosome. Virus Res. 2003, 97, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tan, Y.; Yin, F.; Deng, F.; Vlak, J.M.; Hu, Z.; Wang, H. The F protein of Helicoverpa armigera single nucleopolyhedrovirus can be substituted functionally with its homologue from Spodoptera exigua multiple nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hughes, P.R.; van Beek, N.A.M.; Wood, H.A. A modified droplet feeding method for rapid assay of Bacillus thuringiensis and baculoviruses in noctuid larvae. J. Invertebr. Pathol. 1986, 48, 187–192. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, Y.; Xu, X.; Westenberg, M.; Vlak, J.M.; Wang, H.; Hu, Z.; Deng, F. Helicoverpa armigera nucleopolyhedrovirus occlusion-derived virus-associated protein, HA100, affects oral infectivity in vivo but not virus replication in vitro. J. Gen. Virol. 2011, 92, 1324–1331. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | LC50 (×104 OBs/mL) | SE | 95% Confidence Limit | |
|---|---|---|---|---|
| Upper Bound | Lower Bound | |||
| vHaBacΔegtdsJHAMT | 1.24 | 0.49 | 2.42 | 0.49 |
| vHaBacΔegtdsJHBP | 2.26 | 0.81 | 4.21 | 1.05 |
| vHaBacΔegt | 8.12 | 1.16 | 10.7 | 6.16 |
| Virus | LT50 (Hours Post Infection) | SE | 95% Confidence Limit | |
|---|---|---|---|---|
| Upper Bound | Lower Bound | |||
| vHaBacΔegtdsJHAMT | 120 | 0.89 | 118.26 | 121.74 |
| vHaBacΔegtdsJHBP | 65 | 1.08 | 62.88 | 67.12 |
| vHaBacΔegt | 120 | 1.02 | 118.01 | 122.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Wang, X.; Dai, Y.; Wei, X.; Ni, M.; Zhang, L.; Zhu, Z. Expressing Double-Stranded RNAs of Insect Hormone-Related Genes Enhances Baculovirus Insecticidal Activity. Int. J. Mol. Sci. 2019, 20, 419. https://doi.org/10.3390/ijms20020419
Liu Z, Wang X, Dai Y, Wei X, Ni M, Zhang L, Zhu Z. Expressing Double-Stranded RNAs of Insect Hormone-Related Genes Enhances Baculovirus Insecticidal Activity. International Journal of Molecular Sciences. 2019; 20(2):419. https://doi.org/10.3390/ijms20020419
Chicago/Turabian StyleLiu, Zheming, Xiaofang Wang, Yan Dai, Xiaoli Wei, Mi Ni, Lei Zhang, and Zhen Zhu. 2019. "Expressing Double-Stranded RNAs of Insect Hormone-Related Genes Enhances Baculovirus Insecticidal Activity" International Journal of Molecular Sciences 20, no. 2: 419. https://doi.org/10.3390/ijms20020419