The Biophysical Interaction of the Danger-Associated Molecular Pattern (DAMP) Calreticulin with the Pattern-Associated Molecular Pattern (PAMP) Lipopolysaccharide

Abstract
:1. Introduction
2. Results
2.1. Native Human and Recombinant Calreticulin and Calreticulin Domains are Associated with Endotoxin (LPS)
2.2. Calreticulin Associates With Certain Gram-Negative and Gram-Positive Bacteria
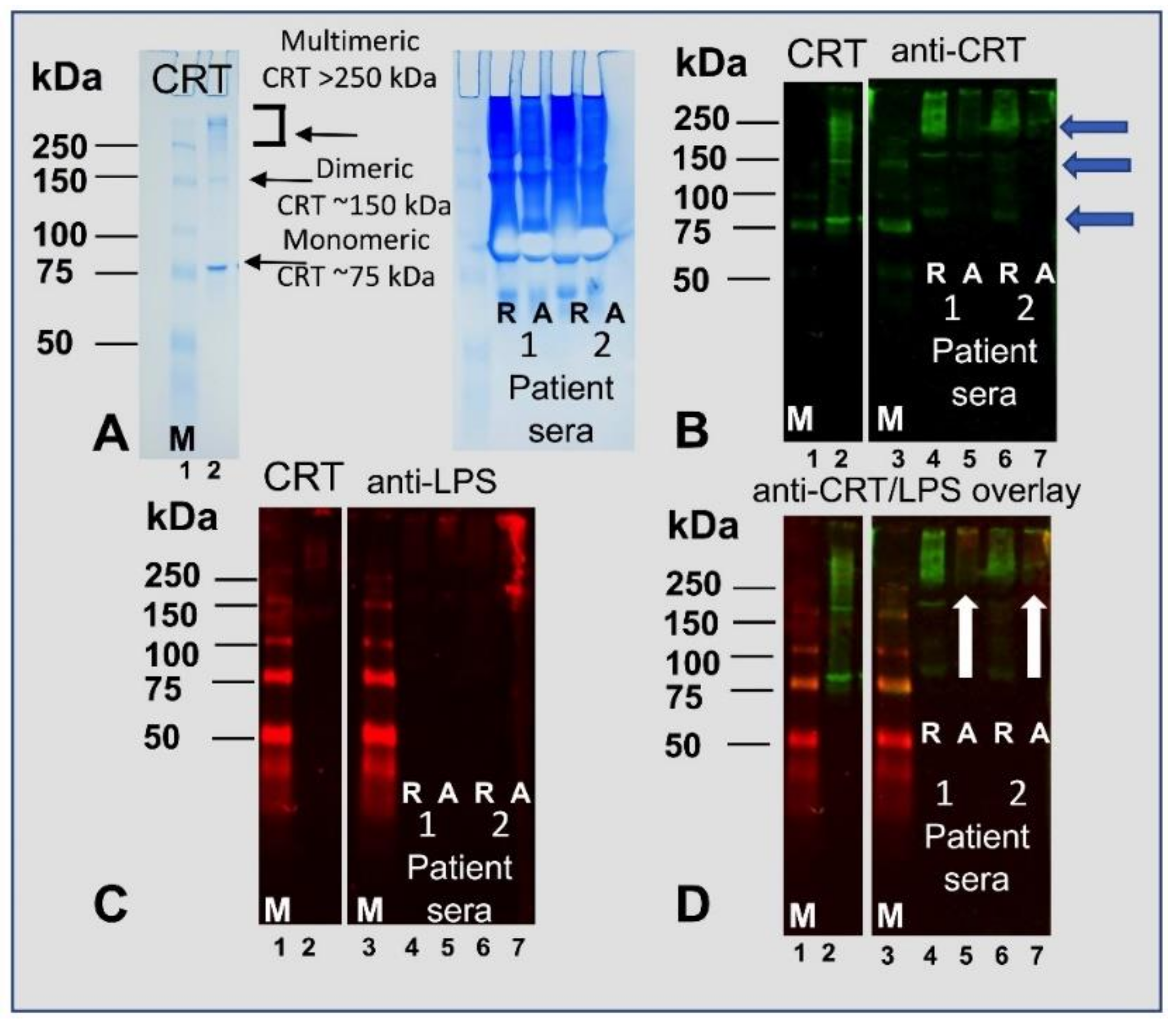
2.3. Lipopolysaccharide Induces Oilgomerization of Calreticulin and Simultaneously Decreases the Amount of the Monomeric Form
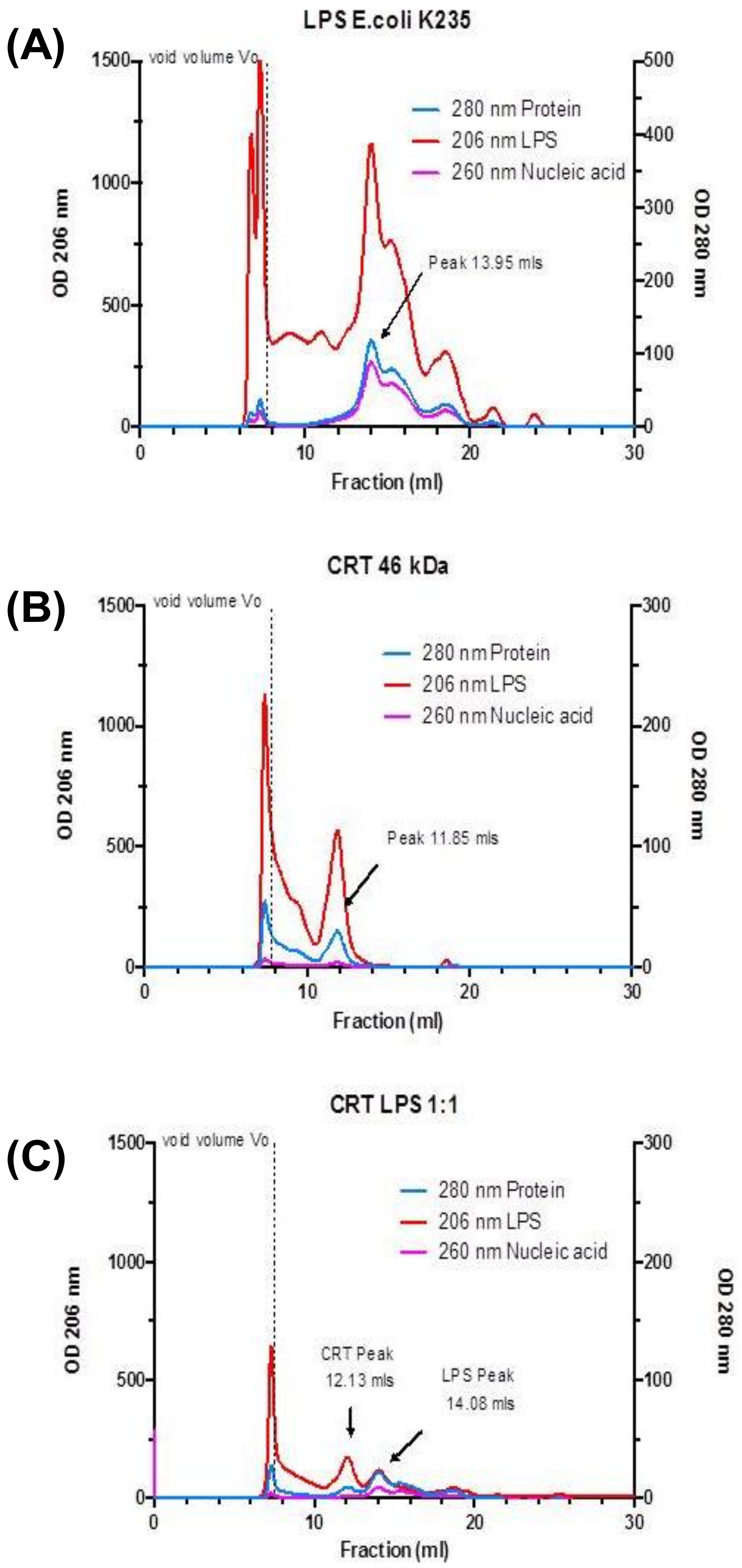
2.4. Multimeric Forms of Human Recombinant Calreticulin are Demonstrated by Ion Exchange and Size Exclusion Chromatography
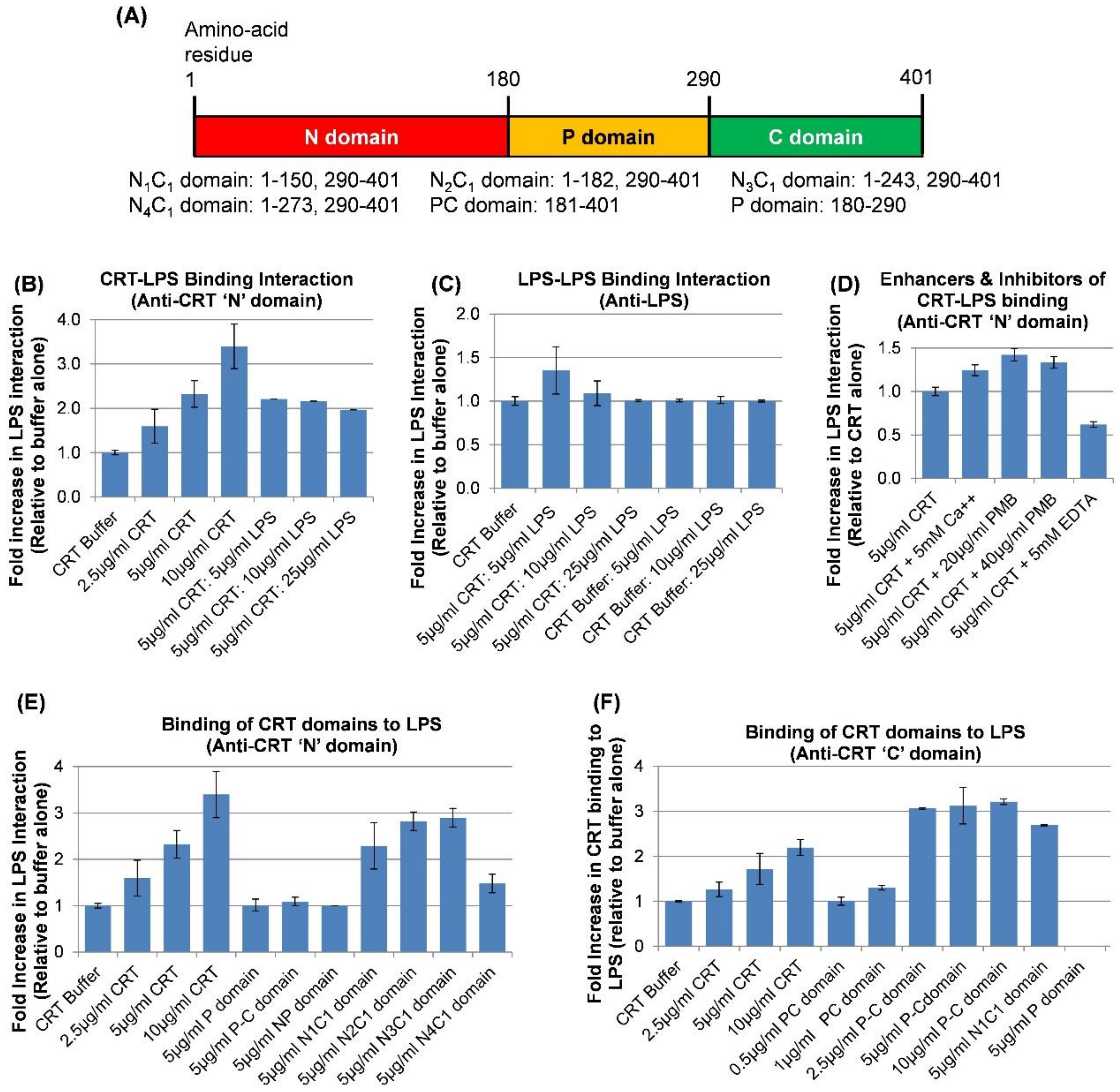
2.5. Direct Physical Binding of CRT to LPS through the N-C Globular Head Region and the C-domain Determined by Solid Phase Assay
3. Discussion
4. Materials and Methods
4.1. Antibodies, Recombinant Proteins and Reagents
4.2. Limulus Amebocyte Lysate (LAL) Endotoxin Detection Assay
4.3. Calreticulin and Calreticulin Domains
4.4. Binding of Calreticulin to Bacteria
4.5. Binding of Lipopolysaccharide to Calreticulin Detected by Immunoblotting
4.6. Anion Exchange Chromatography and ELISA of Oligomeric CRT
4.7. Size Exclusion Chromatographic Analysis of CRT
4.8. Binding of CRT to LPS by Solid Phase Assay (ELISA)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CRT | Calreticulin |
| DAMP | Damage-associated molecular pattern |
| ER | Endoplasmic reticulum |
| HMGB1 | High mobility group box 1 |
| ICD | Immunogenic cell death |
| LAL | Limulus Amoebocyte Lysate |
| LPS | Lipopolysaccharide |
| MFI | Mean fluorescence intensity |
| MHC | Major histocompatibility complex |
| Mr | Relative molecular mass |
| PAMP | Pathogen-associated molecular pattern |
| PBST | PBS with 0.1% tween |
| PBS | Phosphate-buffered saline |
| PS | Phosphotidyl serine |
| PMB | Polymyxin B |
| pI | Isoelectric point |
| rCRT | Recombinant CRT |
| SDS | Sodium dodecyl sulfate |
| TBST | Tris-buffered saline containing 0.1% Tween |
| TLR | Toll-like receptor |
| TMB | Tetramethylbenzidine |
References
- Michalak, M.; Robert Parker, J.M.; Opas, M. Ca2+ signaling and calcium binding chaperones of the endoplasmic reticulum. Cell Calcium 2002, 32, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Williams, D.; Groenendyk, J.; Michalak, M.; Eggleton, P. Unfolding the complexities of ER chaperones in health and disease: Report on the 11th international calreticulin workshop. Cell Stress Chaperones 2015, 20, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Groenendyk, J.; Szabo, E.; Gold, L.I.; Opas, M. Calreticulin, a multi-process calcium-buffering chaperone of the endoplasmic reticulum. Biochem. J. 2009, 417, 651–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, L.I.; Eggleton, P.; Sweetwyne, M.T.; Van Duyn, L.B.; Greives, M.R.; Naylor, S.M.; Michalak, M.; Murphy-Ullrich, J.E. Calreticulin: Non-endoplasmic reticulum functions in physiology and disease. FASEB J. 2010, 24, 665–683. [Google Scholar] [CrossRef] [PubMed]
- Groenendyk, J.; Lee, D.; Jung, J.; Dyck, J.R.; Lopaschuk, G.D.; Agellon, L.B.; Michalak, M. Inhibition of the Unfolded Protein Response Mechanism Prevents Cardiac Fibrosis. PLoS ONE 2016, 11, e0159682. [Google Scholar] [CrossRef] [PubMed]
- Charonis, A.S.; Michalak, M.; Groenendyk, J.; Agellon, L.B. Endoplasmic reticulum in health and disease: The 12th International Calreticulin Workshop, Delphi, Greece. J. Cell. Mol. Med. 2017, 21, 3141–3149. [Google Scholar] [CrossRef] [PubMed]
- Chandrahas, V.K.; Han, J.; Kaufman, R.J. Coordinating Organismal Metabolism during Protein Misfolding in the ER Through the Unfolded Protein Response. Curr. Top. Microbiol. Immunol. 2018, 414, 103–130. [Google Scholar]
- Del Cid, N.; Jeffery, E.; Rizvi, S.M.; Stamper, E.; Peters, L.R.; Brown, W.C.; Provoda, C.; Raghavan, M. Modes of calreticulin recruitment to the major histocompatibility complex class I assembly pathway. J. Biol. Chem. 2010, 285, 4520–4535. [Google Scholar] [CrossRef]
- Raghavan, M.; Wijeyesakere, S.J.; Peters, L.R.; Del Cid, N. Calreticulin in the immune system: Ins and outs. Trends Immunol. 2013, 34, 13–21. [Google Scholar] [CrossRef]
- Peters, L.R.; Raghavan, M. Endoplasmic reticulum calcium depletion impacts chaperone secretion, innate immunity, and phagocytic uptake of cells. J. Immunol. 2011, 187, 919–931. [Google Scholar] [CrossRef]
- Obeid, M.; Tesniere, A.; Ghiringhelli, F.; Fimia, G.M.; Apetoh, L.; Perfettini, J.L.; Castedo, M.; Mignot, G.; Panaretakis, T.; Casares, N.; et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat. Med. 2007, 13, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Tesniere, A.; Apetoh, L.; Ghiringhelli, F.; Joza, N.; Panaretakis, T.; Kepp, O.; Schlemmer, F.; Zitvogel, L.; Kroemer, G. Immunogenic cancer cell death: A key-lock paradigm. Curr. Opin. Immunol. 2008, 20, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Apetoh, L.; Ghiringhelli, F.; Andre, F.; Tesniere, A.; Kroemer, G. The anticancer immune response: Indispensable for therapeutic success? J. Clin. Investig. 2008, 118, 1991–2001. [Google Scholar] [CrossRef] [PubMed]
- Kepp, O.; Gdoura, A.; Martins, I.; Panaretakis, T.; Schlemmer, F.; Tesniere, A.; Fimia, G.M.; Ciccosanti, F.; Burgevin, A.; Piacentini, M.; et al. Lysyl tRNA synthetase is required for the translocation of calreticulin to the cell surface in immunogenic death. Cell Cycle 2010, 9, 3072–3077. [Google Scholar] [CrossRef] [PubMed]
- Vatner, R.E.; Cooper, B.T.; Vanpouille-Box, C.; Demaria, S.; Formenti, S.C. Combinations of immunotherapy and radiation in cancer therapy. Front. Oncol. 2014, 4, 325. [Google Scholar] [CrossRef] [PubMed]
- Golden, E.B.; Frances, D.; Pellicciotta, I.; Demaria, S.; Helen Barcellos-Hoff, M.; Formenti, S.C. Radiation fosters dose-dependent and chemotherapy-induced immunogenic cell death. Oncoimmunology 2014, 3, e28518. [Google Scholar] [CrossRef] [PubMed]
- Bajor, A.; Tischer, S.; Figueiredo, C.; Wittmann, M.; Immenschuh, S.; Blasczyk, R.; Eiz-Vesper, B. Modulatory role of calreticulin as chaperokine for dendritic cell-based immunotherapy. Clin. Exp. Immunol. 2011, 165, 220–234. [Google Scholar] [CrossRef]
- Tarr, J.; Eggleton, P. Immune function of C1q and its modulators CD91 and CD93. Crit. Rev. Immunol. 2005, 25, 305–330. [Google Scholar] [CrossRef]
- Eggleton, P.; Lieu, T.S.; Zappi, E.G.; Sastry, K.; Coburn, J.; Zaner, K.S.; Sontheimer, R.D.; Capra, J.D.; Ghebrehiwet, B.; Tauber, A.I. Calreticulin is released from activated neutrophils and binds to C1q and mannan-binding protein. Clin. Immunol. Immunopathol. 1994, 72, 405–409. [Google Scholar] [CrossRef]
- Paidassi, H.; Tacnet-Delorme, P.; Verneret, M.; Gaboriaud, C.; Houen, G.; Duus, K.; Ling, W.L.; Arlaud, G.J.; Frachet, P. Investigations on the C1q-calreticulin-phosphatidylserine interactions yield new insights into apoptotic cell recognition. J. Mol. Biol. 2011, 408, 277–290. [Google Scholar] [CrossRef]
- Verneret, M.; Tacnet-Delorme, P.; Osman, R.; Awad, R.; Grichine, A.; Kleman, J.P.; Frachet, P. Relative contribution of c1q and apoptotic cell-surface calreticulin to macrophage phagocytosis. J. Innate Immun. 2014, 6, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Kishore, U.; Sontheimer, R.D.; Sastry, K.N.; Zappi, E.G.; Hughes, G.R.; Khamashta, M.A.; Reid, K.B.; Eggleton, P. The systemic lupus erythematosus (SLE) disease autoantigen-calreticulin can inhibit C1q association with immune complexes. Clin. Exp. Immunol. 1997, 108, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Land, W.G.; Agostinis, P.; Gasser, S.; Garg, A.D.; Linkermann, A. DAMP-Induced Allograft and Tumor Rejection: The Circle Is Closing. Am. J. Transplant. 2016, 16, 3322–3337. [Google Scholar] [CrossRef] [PubMed]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Tang, Y.; Li, W.; Wang, X.; Zhang, R.; Zhang, X.; Zhao, X.; Liu, J.; Tang, C.; Liu, Z.; et al. The Endotoxin Delivery Protein HMGB1 Mediates Caspase-11-Dependent Lethality in Sepsis. Immunity 2018, 49, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Bak, S.P.; Amiel, E.; Walters, J.J.; Berwin, B. Calreticulin requires an ancillary adjuvant for the induction of efficient cytotoxic T cell responses. Mol. Immunol. 2008, 45, 1414–1423. [Google Scholar] [CrossRef] [Green Version]
- Reed, R.C.; Berwin, B.; Baker, J.P.; Nicchitta, C.V. GRP94/gp96 elicits ERK activation in murine macrophages. A role for endotoxin contamination in NF-kappa B activation and nitric oxide production. J. Biol. Chem. 2003, 278, 31853–31860. [Google Scholar] [CrossRef]
- Kelley, S.L.; Lukk, T.; Nair, S.K.; Tapping, R.I. The crystal structure of human soluble CD14 reveals a bent solenoid with a hydrophobic amino-terminal pocket. J. Immunol. 2013, 190, 1304–1311. [Google Scholar] [CrossRef]
- Kenneth Todar, P. Todar’s Online Textbook of Bacteriology. Available online: http://textbookofbacteriology.net/endotoxin.html (accessed on 16 November 2018).
- Qureshi, N.; Takayama, K.; Mascagni, P.; Honovich, J.; Wong, R.; Cotter, R.J. Complete structural determination of lipopolysaccharide obtained from deep rough mutant of Escherichia coli. Purification by high performance liquid chromatography and direct analysis by plasma desorption mass spectrometry. J. Biol. Chem. 1988, 263, 11971–11976. [Google Scholar]
- Vagenende, V.; Ching, T.J.; Chua, R.J.; Gagnon, P. Allantoin as a solid phase adsorbent for removing endotoxins. J. Chromatogr. A 2013, 1310, 15–20. [Google Scholar] [CrossRef]
- Gusarov, D.; Sokolova, I.; Vorobjeva, T.; Brykova, N. Positively charged proteins: Separation and depyrogenation by means of HPLC (by the example of recombinant histone H1.3 variant). Biopharm. J. 2011, 3, 16–23. [Google Scholar]
- Kozlov, G.; Pocanschi, C.L.; Rosenauer, A.; Bastos-Aristizabal, S.; Gorelik, A.; Williams, D.B.; Gehring, K. Structural basis of carbohydrate recognition by calreticulin. J. Biol. Chem. 2010, 285, 38612–38620. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Michalak, M.; Opas, M.; Eggleton, P. The ins and outs of calreticulin: From the ER lumen to the extracellular space. Trends Cell Biol. 2001, 11, 122–129. [Google Scholar] [CrossRef]
- Chouquet, A.; Paidassi, H.; Ling, W.L.; Frachet, P.; Houen, G.; Arlaud, G.J.; Gaboriaud, C. X-ray structure of the human calreticulin globular domain reveals a peptide-binding area and suggests a multi-molecular mechanism. PLoS ONE 2011, 6, e17886. [Google Scholar] [CrossRef] [PubMed]
- Michalak, M.; Corbett, E.F.; Mesaeli, N.; Nakamura, K.; Opas, M. Calreticulin: One protein, one gene, many functions. Biochem. J. 1999, 344 Pt 2, 281–292. [Google Scholar] [CrossRef]
- Wijeyesakere, S.J.; Bedi, S.K.; Huynh, D.; Raghavan, M. The C-Terminal Acidic Region of Calreticulin Mediates Phosphatidylserine Binding and Apoptotic Cell Phagocytosis. J. Immunol. 2016, 196, 3896–3909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boelt, S.G.; Norn, C.; Rasmussen, M.I.; Andre, I.; Ciplys, E.; Slibinskas, R.; Houen, G.; Hojrup, P. Mapping the Ca(2+) induced structural change in calreticulin. J. Prot. 2016, 142, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Pockley, A.G.; Muthana, M.; Calderwood, S.K. The dual immunoregulatory roles of stress proteins. Trends Biochem. Sci. 2008, 33, 71–79. [Google Scholar] [CrossRef]
- Henderson, B.; Calderwood, S.K.; Coates, A.R.; Cohen, I.; van Eden, W.; Lehner, T.; Pockley, A.G. Caught with their PAMPs down? The extracellular signalling actions of molecular chaperones are not due to microbial contaminants. Cell Stress Chaperones 2010, 15, 123–141. [Google Scholar] [CrossRef]
- Gold, L.I.; Pandya, U.M.; Panchal, R.G. Calreticulin: Immunoregulatory and Anitmicrobial Activities. Unpublished work. 2019. [Google Scholar]
- Tarr, J.M.; Winyard, P.G.; Ryan, B.; Harries, L.W.; Haigh, R.; Viner, N.; Eggleton, P. Extracellular calreticulin is present in the joints of patients with rheumatoid arthritis and inhibits FasL (CD95L)-mediated apoptosis of T cells. Arthritis Rheum 2011, 62, 2919–2929. [Google Scholar] [CrossRef]
- Quirke, A.M.; Perry, E.; Cartwright, A.; Kelly, C.; De Soyza, A.; Eggleton, P.; Hutchinson, D.; Venables, P.J. Bronchiectasis is a Model for Chronic Bacterial Infection Inducing Autoimmunity in Rheumatoid Arthritis. Arthritis Rheumatol. (Hoboken, NJ) 2015, 67, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Andrin, C.; Corbett, E.F.; Johnson, S.; Dabrowska, M.; Campbell, I.D.; Eggleton, P.; Opas, M.; Michalak, M. Expression and purification of mammalian calreticulin in Pichia pastoris. Protein Expr. Purif. 2000, 20, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Baksh, S.; Burns, K.; Busaan, J.; Michalak, M. Expression and purification of recombinant and native calreticulin. Protein Expr. Purif. 1992, 3, 322–331. [Google Scholar] [CrossRef]
- Pier, G.B.; Sidberry, H.F.; Zolyomi, S.; Sadoff, J.C. Isolation and characterization of a high-molecular-weight polysaccharide from the slime of Pseudomonas aeruginosa. Infect Immun. 1978, 22, 908–918. [Google Scholar] [PubMed]
- Domingues, M.M.; Inacio, R.G.; Raimundo, J.M.; Martins, M.; Castanho, M.A.; Santos, N.C. Biophysical characterization of polymyxin B interaction with LPS aggregates and membrane model systems. Biopolymers 2012, 98, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Mares, J.; Kumaran, S.; Gobbo, M.; Zerbe, O. Interactions of lipopolysaccharide and polymyxin studied by NMR spectroscopy. J. Biol. Chem. 2009, 284, 11498–11506. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xu, N.; Zhang, S. Calreticulin is a microbial-binding molecule with phagocytosis-enhancing capacity. Fish Shellfish Immunol. 2013, 35, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Jiang, Z.; Zhang, M.; Yang, N.; Zhu, D. Identification of a new calreticulin homolog from Yesso scallop (Patinopecten yessoensis) and its role in innate immunity. Fish Shellfish Immunol. 2016, 58, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Jiang, Z.; Yang, N.; Zhu, D.; Zhang, M. Identification and characterization of a novel calreticulin involved in the immune response of the Zhikong scallop, Chlamys farreri. Fish Shellfish Immunol. 2017, 64, 251–259. [Google Scholar] [CrossRef]
- Huang, Y.; Hui, K.; Jin, M.; Yin, S.; Wang, W.; Ren, Q. Two endoplasmic reticulum proteins (calnexin and calreticulin) are involved in innate immunity in Chinese mitten crab (Eriocheir sinensis). Sci. Rep. 2016, 6, 27578. [Google Scholar] [CrossRef]
- Brito, L.A.; Singh, M. Acceptable levels of endotoxin in vaccine formulations during preclinical research. J. Pharm. Sci. 2011, 100, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Malyala, P.; Singh, M. Endotoxin limits in formulations for preclinical research. J. Pharm. Sci. 2008, 97, 2041–2044. [Google Scholar] [CrossRef] [PubMed]
- Nanney, L.B.; Woodrell, C.D.; Greives, M.R.; Cardwell, N.L.; Pollins, A.C.; Bancroft, T.A.; Chesser, A.; Michalak, M.; Rahman, M.; Siebert, J.W.; et al. Calreticulin enhances porcine wound repair by diverse biological effects. Am. J. Pathol. 2008, 173, 610–630. [Google Scholar] [CrossRef]
- Greives, M.R.; Samra, F.; Pavlides, S.C.; Blechman, K.M.; Naylor, S.M.; Woodrell, C.D.; Cadacio, C.; Levine, J.P.; Bancroft, T.A.; Michalak, M.; et al. Exogenous calreticulin improves diabetic wound healing. Wound Repair Regen. 2012, 20, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Pandya, U.M.; Gold, L.I. The Novel Biotherapeutic Calreticulin (CRT) Corrects Multiple Defects of Non-Healing Diabetic Wounds. J. Dermatol. Clin. Res. 2016, 4, 1083–1093. [Google Scholar]
- Cleveland, M.G.; Gorham, J.D.; Murphy, T.L.; Tuomanen, E.; Murphy, K.M. Lipoteichoic acid preparations of gram-positive bacteria induce interleukin-12 through a CD14-dependent pathway. Infect Immun. 1996, 64, 1906–1912. [Google Scholar]
- Hattar, K.; Grandel, U.; Moeller, A.; Fink, L.; Iglhaut, J.; Hartung, T.; Morath, S.; Seeger, W.; Grimminger, F.; Sibelius, U. Lipoteichoic acid (LTA) from Staphylococcus aureus stimulates human neutrophil cytokine release by a CD14-dependent, Toll-like-receptor-independent mechanism: Autocrine role of tumor necrosis factor-[alpha] in mediating LTA-induced interleukin-8 generation. Crit. Care Med. 2006, 34, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Pockley, A.G.; Henderson, B. Extracellular cell stress (heat shock) proteins-immune responses and disease: An overview. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373, 1738. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Triantafilou, K. Cellular Trafficking of Cell Stress Proteins in Health and Disease; Springer: Dordrecht, The Netherlands, 2012; Volume 6. [Google Scholar]
- Habich, C.; Kempe, K.; van der Zee, R.; Rumenapf, R.; Akiyama, H.; Kolb, H.; Burkart, V. Heat shock protein 60: Specific binding of lipopolysaccharide. J. Immunol. 2005, 174, 1298–1305. [Google Scholar] [CrossRef]
- Basu, S.; Binder, R.J.; Suto, R.; Anderson, K.M.; Srivastava, P.K. Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-kappa B pathway. Int. Immunol. 2000, 12, 1539–1546. [Google Scholar] [CrossRef]
- Osman, R.; Tacnet-Delorme, P.; Kleman, J.P.; Millet, A.; Frachet, P. Calreticulin Release at an Early Stage of Death Modulates the Clearance by Macrophages of Apoptotic Cells. Front. Immunol. 2017, 8, 1034. [Google Scholar] [CrossRef]
- Duo, C.C.; Gong, F.Y.; He, X.Y.; Li, Y.M.; Wang, J.; Zhang, J.P.; Gao, X.M. Soluble calreticulin induces tumor necrosis factor-alpha (TNF-alpha) and interleukin (IL)-6 production by macrophages through mitogen-activated protein kinase (MAPK) and NFkappaB signaling pathways. Int. J. Mol. Sci. 2014, 15, 2916–2928. [Google Scholar] [CrossRef]
- He, M.C.; Wang, J.; Wu, J.; Gong, F.Y.; Hong, C.; Xia, Y.; Zhang, L.J.; Bao, W.R.; Gao, X.M. Immunological activity difference between native calreticulin monomers and oligomers. PLoS ONE 2014, 9, e105502. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Qiu, X.; Li, Y.; Huang, Q.; Zhong, Z.; Zhang, Y.; Liu, X.; Sun, L.; Lv, P.; Gao, X.M. Functional analysis of recombinant calreticulin fragment 39-272: Implications for immunobiological activities of calreticulin in health and disease. J. Immunol. 2010, 185, 4561–4569. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Zhao, L.X.; Hong, C.; Duo, C.C.; Guo, B.N.; Zhang, L.J.; Gong, Z.; Xiong, S.D.; Gong, F.Y.; Gao, X.M. Self-oligomerization is essential for enhanced immunological activities of soluble recombinant calreticulin. PLoS ONE 2013, 8, e64951. [Google Scholar] [CrossRef] [PubMed]
- Pawaria, S.; Binder, R.J. CD91-dependent programming of T-helper cell responses following heat shock protein immunization. Nat. Commun. 2011, 2, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawaria, S.; Kropp, L.E.; Binder, R.J. Immunotherapy of tumors with alpha2-macroglobulin-antigen complexes pre-formed in vivo. PLoS ONE 2012, 7, e50365. [Google Scholar] [CrossRef] [PubMed]
- Osterloh, A.; Breloer, M. Heat shock proteins: Linking danger and pathogen recognition. Med. Microbial. Immunol. 2008, 197, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Zhang, T.; Gao, X.M. Recombinant murine calreticulin fragment 39-272 expands CD1d(hi)CD5+ IL-10-secreting B cells that modulate experimental autoimmune encephalomyelitis in C57BL/6 mice. Mol. Immunol. 2013, 55, 237–246. [Google Scholar] [CrossRef]
- Young, P.; Sezestakowska, D.; Morse, R.; Winyard, P.G.; Whatmore, J.; Reid, K.B. Purification, isolation and characterization of native and recombinant calreticulin. Calcium Bind. Proteins 2006, 1, 160–169. [Google Scholar]
- Corbett, E.F.; Michalak, K.M.; Oikawa, K.; Johnson, S.; Campbell, I.D.; Eggleton, P.; Kay, C.; Michalak, M. The conformation of calreticulin is influenced by the endoplasmic reticulum luminal environment. J. Biol. Chem. 2000, 275, 27177–27185. [Google Scholar] [PubMed]
- Petsch, D.; Deckwer, W.D.; Anspach, F.B. Proteinase K digestion of proteins improves detection of bacterial endotoxins by the Limulus amebocyte lysate assay: Application for endotoxin removal from cationic proteins. Anal. Biochem. 1998, 259, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Paramonov, N.A.; Aduse-Opoku, J.; Hashim, A.; Rangarajan, M.; Curtis, M.A. Structural analysis of the core region of O-lipopolysaccharide of Porphyromonas gingivalis from mutants defective in O-antigen ligase and O-antigen polymerase. J. Bacterial. 2009, 191, 5272–5282. [Google Scholar] [CrossRef] [PubMed]
- Chaput, N.; De Botton, S.; Obeid, M.; Apetoh, L.; Ghiringhelli, F.; Panaretakis, T.; Flament, C.; Zitvogel, L.; Kroemer, G. Molecular determinants of immunogenic cell death: Surface exposure of calreticulin makes the difference. J. Mol. Med. 2007, 85, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Ciplys, E.; Zitkus, E.; Gold, L.I.; Daubriac, J.; Pavlides, S.C.; Hojrup, P.; Houen, G.; Wang, W.A.; Michalak, M.; Slibinskas, R. High-level secretion of native recombinant human calreticulin in yeast. Microb. Cell Fact. 2015, 14, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojrup, P.; Roepstorff, P.; Houen, G. Human placental calreticulin characterization of domain structure and post-translational modifications. Eur. J. Biochem. 2001, 268, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Honore, C.; Hummelshoj, T.; Hansen, B.E.; Madsen, H.O.; Eggleton, P.; Garred, P. The innate immune component ficolin 3 (Hakata antigen) mediates the clearance of late apoptotic cells. Arthritis Rheum 2007, 56, 1598–1607. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRT Source | Endotoxin Level (EU/µg) |
|---|---|
| Recombinant CRT from Escherichia coli | 0.5 ± 0.4 (n = 6) |
| Recombinant CRT from Pichia pastoris | 0.1 ± 0.02 (n = 3) |
| Recombinant CRT from Saccharomyces Cerevisiae | 1.3 (n = 1) |
| Native CRT from human placenta | 2.0 (n = 1) |
| Recombinant CRT NP-domain | 1.7 (n = 1) |
| Recombinant CRT N1C1 domain | 1.3 ± 0.6 (n = 2) |
| Recombinant CRT P-domain | 0.4 ± 0.5 (n = 3) |
| Recombinant CRT PC domain | 1.3 ± 0.2 (n = 2) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandya, U.M.; Egbuta, C.; Abdullah Norman, T.M.; Chiang, C.-Y.; Wiersma, V.R.; Panchal, R.G.; Bremer, E.; Eggleton, P.; Gold, L.I. The Biophysical Interaction of the Danger-Associated Molecular Pattern (DAMP) Calreticulin with the Pattern-Associated Molecular Pattern (PAMP) Lipopolysaccharide. Int. J. Mol. Sci. 2019, 20, 408. https://doi.org/10.3390/ijms20020408
Pandya UM, Egbuta C, Abdullah Norman TM, Chiang C-Y, Wiersma VR, Panchal RG, Bremer E, Eggleton P, Gold LI. The Biophysical Interaction of the Danger-Associated Molecular Pattern (DAMP) Calreticulin with the Pattern-Associated Molecular Pattern (PAMP) Lipopolysaccharide. International Journal of Molecular Sciences. 2019; 20(2):408. https://doi.org/10.3390/ijms20020408
Chicago/Turabian StylePandya, Unnati M., Chinaza Egbuta, Trefa M. Abdullah Norman, Chih-Yuan (Edward) Chiang, Valerie R. Wiersma, Rekha G. Panchal, Edwin Bremer, Paul Eggleton, and Leslie I Gold. 2019. "The Biophysical Interaction of the Danger-Associated Molecular Pattern (DAMP) Calreticulin with the Pattern-Associated Molecular Pattern (PAMP) Lipopolysaccharide" International Journal of Molecular Sciences 20, no. 2: 408. https://doi.org/10.3390/ijms20020408