Relation of the pdxB-usg-truA-dedA Operon and the truA Gene to the Intracellular Survival of Salmonella enterica Serovar Typhimurium

 ,
,
Abstract
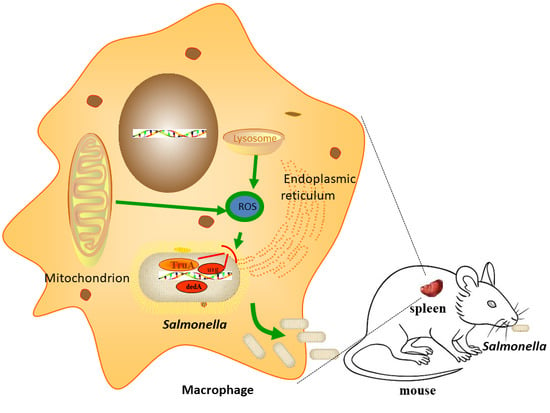
:
1. Introduction
2. Results
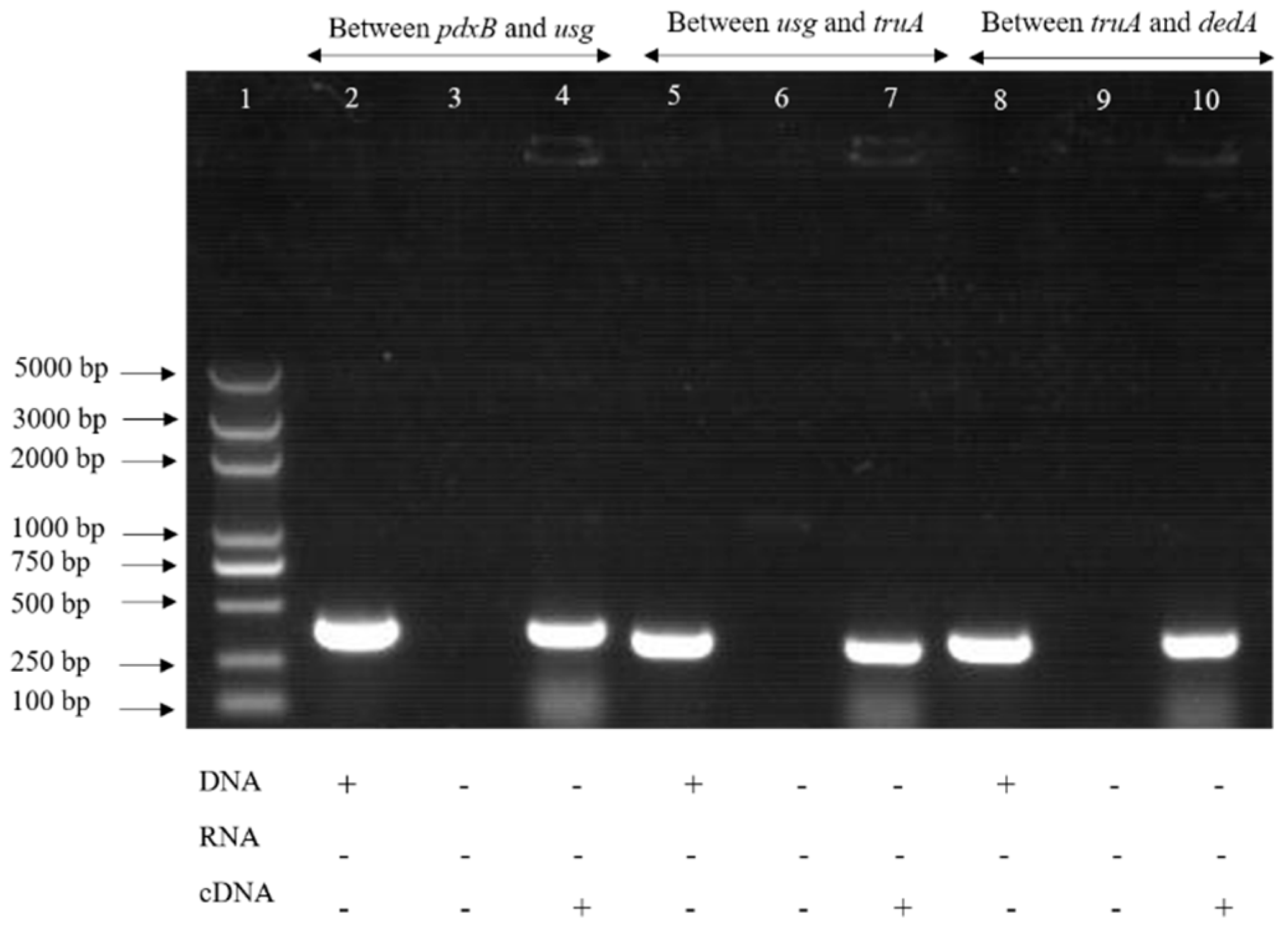
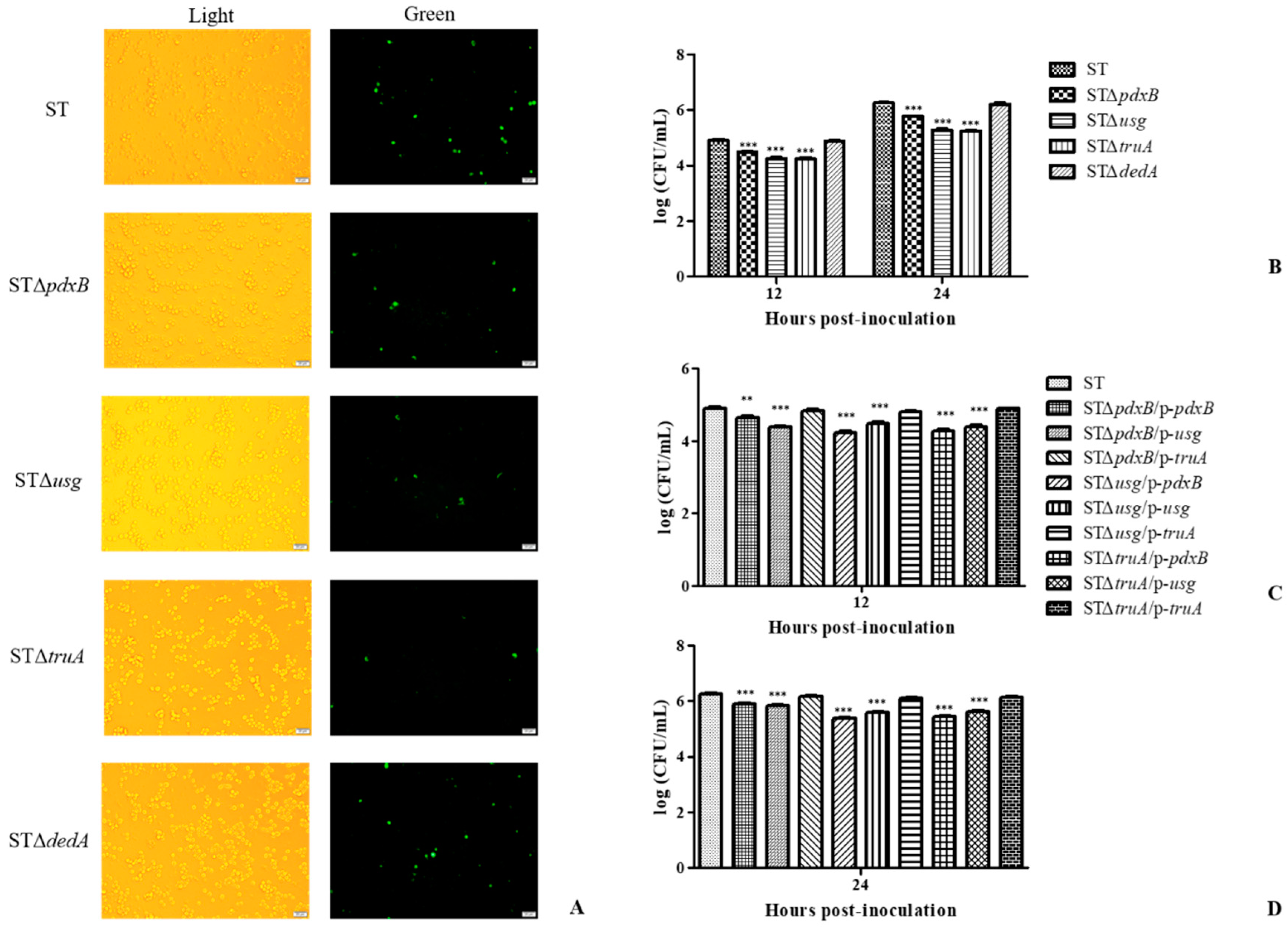
2.1. The pdxB-usg-truA-dedA Operon Is Required for Intracellular Survival of S. enterica
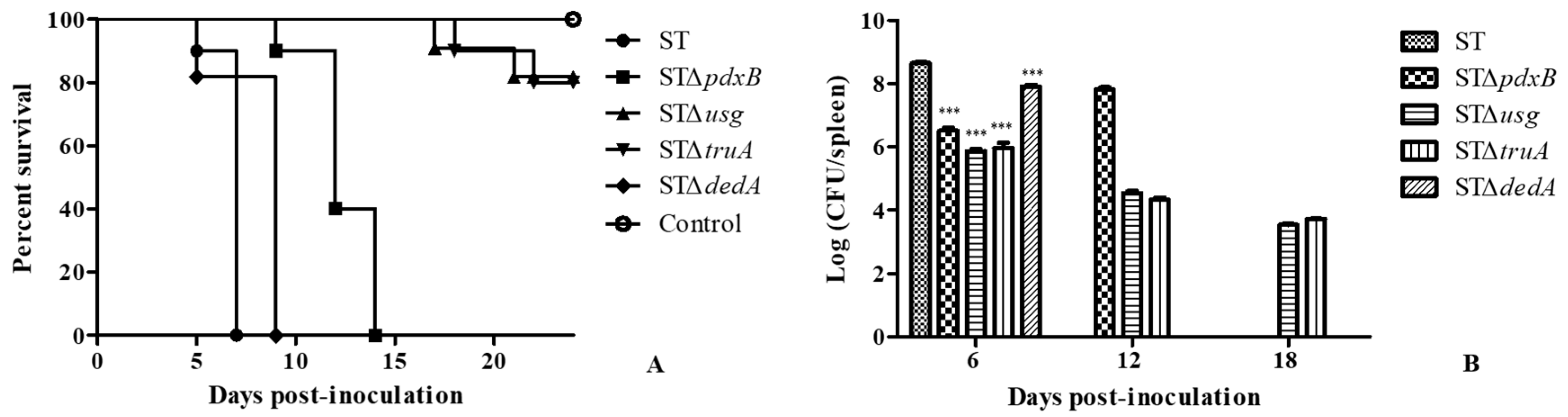
2.2. The pdxB-usg-truA-dedA Operon Contributes to Virulence in Mice
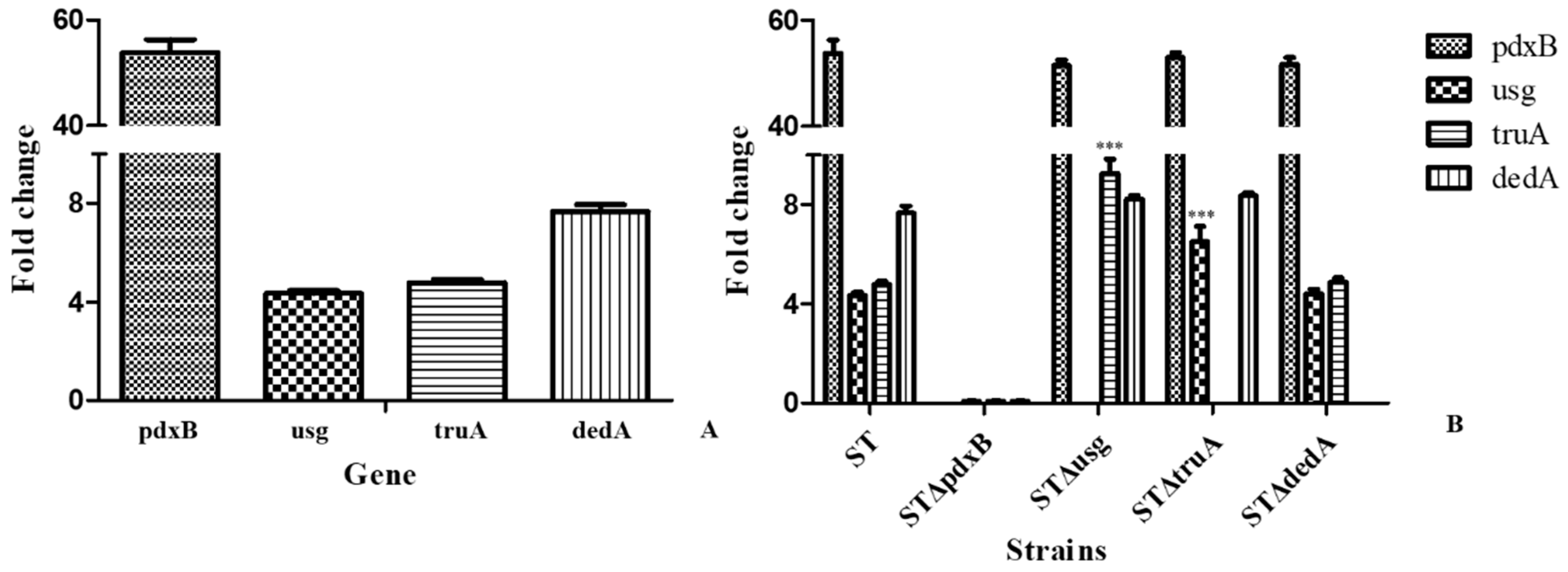
2.3. The usg and truA Expression Levels Were Higher in the Oxidative Environment
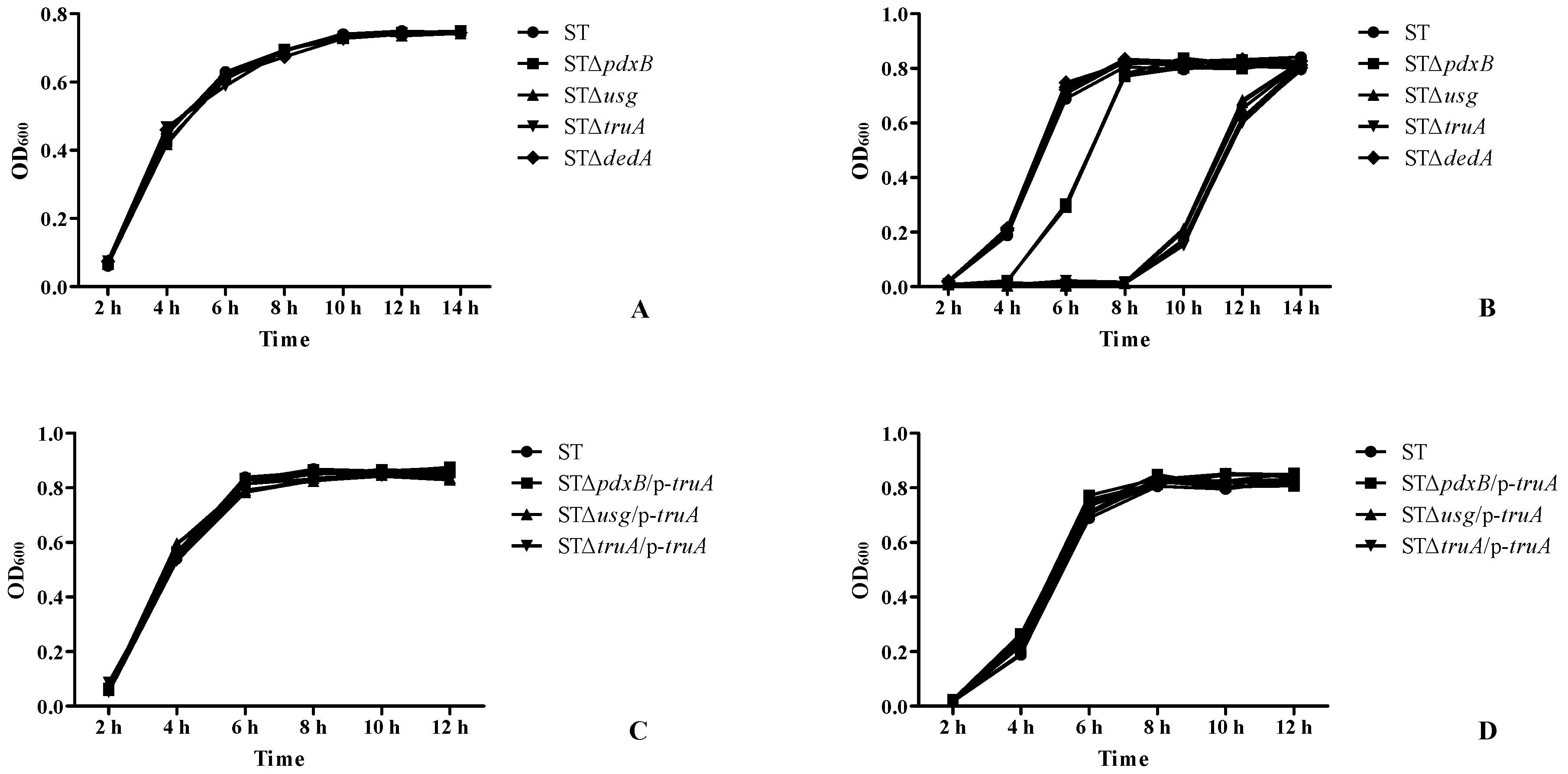
2.4. The pdxB-usg-truA-dedA Operon Contributed to Resistance to Oxidative Conditions
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Bacterial Strains and Media
4.3. Mice
4.4. Construction of Gene Deletion and Complemented Gene Deletion Mutant Strains
4.5. Cell Infection Assay
4.6. Growth Characteristics and Oxidative Resistance Assay
4.7. Gene Expression Levels in an Oxidative Environment
4.8. Virulence in BALB/c Mice
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Baker, S.; Dougan, G. The genome of Salmonella enterica serovar Typhi. Clin. Infect. Dis. 2007, 45 (Suppl. 1), S29–S33. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Grimont, P.A.; Garrity, G.M.; Euzeby, J.P. Nomenclature and taxonomy of the genus Salmonella. Int. J. Syst.Evol. Microbiol. 2005, 55, 521–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vazquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Nyachuba, D.G. Foodborne illness: Is it on the rise? Nutr. Rev. 2010, 68, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, J.; Bing, G.; Bie, P.; De, Y.; Lyu, Y.; Wu, Q. Ortholog-based screening and identification of genes related to intracellular survival. Gene 2018, 651, 134–142. [Google Scholar] [CrossRef]
- Taboada, B.; Ciria, R.; Martinez-Guerrero, C.E.; Merino, E. ProOpDB: Prokaryotic Operon DataBase. Nucleic Acids Res. 2012, 40, D627–D631. [Google Scholar] [CrossRef]
- Ha, J.Y.; Lee, J.H.; Kim, K.H.; Kim, D.J.; Lee, H.H.; Kim, H.K.; Yoon, H.J.; Suh, S.W. Crystal structure of D-erythronate-4-phosphate dehydrogenase complexed with NAD. J. Mol. Biol. 2007, 366, 1294–1304. [Google Scholar] [CrossRef]
- Behnsen, J.; Perez-Lopez, A.; Nuccio, S.P.; Raffatellu, M. Exploiting host immunity: The Salmonella paradigm. Trends Immunol. 2015, 36, 112–120. [Google Scholar] [CrossRef]
- Zhao, G.; Pease, A.J.; Bharani, N.; Winkler, M.E. Biochemical characterization of gapB-encoded erythrose 4-phosphate dehydrogenase of Escherichia coli K-12 and its possible role in pyridoxal 5′-phosphate biosynthesis. J. Bacteriol. 1995, 177, 2804–2812. [Google Scholar] [CrossRef]
- Rudolph, J.; Kim, J.; Copley, S.D. Multiple turnovers of the nicotino-enzyme PdxB require alpha-keto acids as cosubstrates. Biochemistry 2010, 49, 9249–9255. [Google Scholar] [CrossRef]
- Chen, F.; Mackey, A.J.; Vermunt, J.K.; Roos, D.S. Assessing performance of orthology detection strategies applied to eukaryotic genomes. PLoS ONE 2007, 2, e383. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Breis, J.T.; Chiba, H.; Legaz-Garcia Mdel, C.; Uchiyama, I. The Orthology Ontology: Development and applications. J. Biomed. Semant. 2016, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, J.; Kwon, M.; Lee, K.; Min, H.; Kim, S.H.; Kim, D.; Lee, N.; Kim, J.; Kim, D.; et al. Screening for Escherichia coli K-12 genes conferring glyoxal resistance or sensitivity by transposon insertions. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Hao, H.; Dai, M.; Wang, Y.; Huang, L.; Peng, D.; Wang, X.; Wang, H.; Yao, M.; Sun, Y.; et al. Development of quinoxaline 1, 4-dioxides resistance in Escherichia coli and molecular change under resistance selection. PLoS ONE 2012, 7, e43322. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yi, L.; Zhang, J.; Sun, L.; Wen, W.; Zhang, C.; Wang, S. Functional analysis of superoxide dismutase of Salmonella typhimurium in serum resistance and biofilm formation. J. Appl. Microbiol. 2018, 125, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Hebrard, M.; Viala, J.P.; Meresse, S.; Barras, F.; Aussel, L. Redundant hydrogen peroxide scavengers contribute to Salmonella virulence and oxidative stress resistance. J. Bacteriol. 2009, 191, 4605–4614. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Qi, L.; Hu, M.; Liu, Y.; Yu, K.; Liu, Q.; Liu, X. Salmonella proteomics under oxidative stress reveals coordinated regulation of antioxidant defense with iron metabolism and bacterial virulence. J. Proteom. 2017, 157, 52–58. [Google Scholar] [CrossRef]
- Buckner, M.M.; Finlay, B.B. Host-microbe interaction: Innate immunity cues virulence. Nature 2011, 472, 179–180. [Google Scholar] [CrossRef]
- Blumenthal, T.; Davis, P.; Garrido-Lecca, A. Operon and non-operon gene clusters in the C. elegans genome. WormBook 2015, 1–20. [Google Scholar] [CrossRef]
- Eswarappa, S.M.; Panguluri, K.K.; Hensel, M.; Chakravortty, D. The yejABEF operon of Salmonella confers resistance to antimicrobial peptides and contributes to its virulence. Microbiology 2008, 154, 666–678. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, Y.; Zhang, X.; Zhan, L.; Zhao, X.; Xu, S.; Sheng, X.; Huang, X. The novel cis-encoded antisense RNA AsrC positively regulates the expression of rpoE-rseABC operon and thus enhances the motility of Salmonella enterica serovar typhi. Front Microbiol. 2015, 6, 990. [Google Scholar] [CrossRef] [PubMed]
- Brandis, G.; Bergman, J.M.; Hughes, D. Autoregulation of the tufB operon in Salmonella. Mol. Microbiol. 2016, 100, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Ken-Ichiro, I.; Ren, H.Y.; Zhou, H.J.; Yoshida, S. Legionella dumoffii Tex-KL Mutated in an Operon Homologous to traC-traD is Defective in Epithelial Cell Invasion. Biomed. Environ. Sci. 2016, 29, 424–434. [Google Scholar] [PubMed]
- Godfrey, R.E.; Lee, D.J.; Busby, S.J.W.; Browning, D.F. Regulation of nrf operon expression in pathogenic enteric bacteria: Sequence divergence reveals new regulatory complexity. Mol. Microbiol. 2017, 104, 580–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Wang, J.; Qi, X.; Su, J. Transcription Elongation Factor GreA Plays a Key Role in Cellular Invasion and Virulence of Francisella tularensis subsp. novicida. Sci. Rep. 2018, 8, 6895. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Qi, L.; Xiao, Y.; Wang, M.; Qin, C.; Zhang, H.; Sheng, Y.; Du, H. SufC may promote the survival of Salmonella enterica serovar Typhi in macrophages. Microb. Pathog. 2015, 85, 40–43. [Google Scholar] [CrossRef]
- Gong, H.; Vu, G.P.; Bai, Y.; Chan, E.; Wu, R.; Yang, E.; Liu, F.; Lu, S. A Salmonella small non-coding RNA facilitates bacterial invasion and intracellular replication by modulating the expression of virulence factors. PLoS Pathog. 2011, 7, e1002120. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Description/Purpose | Source or Reference |
|---|---|---|
| Strains | ||
| S. typhimurium ATCC14028 (ST) | Wild type (WT) | Guangdong Culture Collection Center |
| STΔpdxB | ΔpdxB mutant of ST by the λ-Red recombination system | This study |
| STΔusg | Δusg mutant of ST by the λ-Red recombination system | Our lab |
| STΔtruA | ΔtruA mutant of ST by the λ-Red recombination system | This study |
| STΔdedA | ΔdedA mutant of ST by the λ-Red recombination system | This study |
| STΔpdxB/p-pdxB | STΔpdxB harboring the pBR322-pdxB plasmid, complement strains | This study |
| STΔpdxB/p-usg | STΔpdxB harboring the pBR322-usg plasmid, complement strains | This study |
| STΔpdxB/p-truA | STΔpdxB harboring the pBR322-truA plasmid, complement strains | This study |
| STΔusg/p-pdxB | STΔusg harboring the pBR322-pdxB plasmid, complement strains | This study |
| STΔusg/p-usg | STΔusg harboring the pBR322-usg plasmid, complement strains | Our lab |
| STΔusg/p-truA | STΔusg harboring the pBR322-truA plasmid, complement strains | This study |
| STΔtruA/p-pdxB | STΔtruA harboring the pBR322-pdxB plasmid, complement strains | This study |
| STΔtruA/p-usg | STΔtruA harboring the pBR322-usg plasmid, complement strains | This study |
| STΔtruA/p-truA | STΔtruA harboring the pBR322-truA plasmid, complement strains | This study |
| DH5α | For cloning | Takara |
| Plasmids | ||
| pKD3, pKD46 and pCP20 | λ-Red recombination system | Datsenko and Wanner, 2000 [26] |
| pBR322 | For constructed complement strains | Virulent laboratory |
| p-pdxB | pdxB of ST product cloned into pBR322 for complementation assay | This study |
| p-usg | usg of ST product cloned into pBR322 for complementation assay | Our lab |
| p-truA | truA of ST product cloned into pBR322 for complementation assay | This study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Wang, J.; Feng, Z.; Zhang, X.; Wang, X.; Wu, Q. Relation of the pdxB-usg-truA-dedA Operon and the truA Gene to the Intracellular Survival of Salmonella enterica Serovar Typhimurium. Int. J. Mol. Sci. 2019, 20, 380. https://doi.org/10.3390/ijms20020380
Yang X, Wang J, Feng Z, Zhang X, Wang X, Wu Q. Relation of the pdxB-usg-truA-dedA Operon and the truA Gene to the Intracellular Survival of Salmonella enterica Serovar Typhimurium. International Journal of Molecular Sciences. 2019; 20(2):380. https://doi.org/10.3390/ijms20020380
Chicago/Turabian StyleYang, Xiaowen, Jiawei Wang, Ziyan Feng, Xiangjian Zhang, Xiangguo Wang, and Qingmin Wu. 2019. "Relation of the pdxB-usg-truA-dedA Operon and the truA Gene to the Intracellular Survival of Salmonella enterica Serovar Typhimurium" International Journal of Molecular Sciences 20, no. 2: 380. https://doi.org/10.3390/ijms20020380