G-Protein Coupled Estrogen Receptor in Breast Cancer


1
Ph.D. Program in Medical Biotechnology, College of Medical Science and Technology, Taipei Medical University, Taipei 110, Taiwan
2
Division of Pulmonary and Critical Care Medicine, Sun Yat-Sen Cancer Center, Taipei 112, Taiwan
3
Department of Medicine, National Yang-Ming University Medical School, Taipei 112, Taiwan
4
Department of Medical Oncology, Sun Yat-Sen Cancer Center, Taipei 112, Taiwan
5
School of Medical Laboratory Science and Biotechnology, College of Medical Science and Technology, Taipei Medical University, 250 Wu-Hsing Street, Taipei 110, Taiwan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(2), 306; https://doi.org/10.3390/ijms20020306
Submission received: 29 November 2018
/
Revised: 10 January 2019
/
Accepted: 12 January 2019
/
Published: 14 January 2019
(This article belongs to the Special Issue Sex Hormone Receptor Signals in Human Malignancies)
Abstract
:The G-protein coupled estrogen receptor (GPER), an alternate estrogen receptor (ER) with a structure distinct from the two canonical ERs, being ERα, and ERβ, is expressed in 50% to 60% of breast cancer tissues and has been presumed to be associated with the development of tamoxifen resistance in ERα positive breast cancer. On the other hand, triple-negative breast cancer (TNBC) constitutes 15% to 20% of breast cancers and frequently displays a more aggressive behavior. GPER is prevalent and involved in TNBC and can be a therapeutic target. However, contradictory results exist regarding the function of GPER in breast cancer, proliferative or pro-apoptotic. A better understanding of the GPER, its role in breast cancer, and the interactions with the ER and epidermal growth factor receptor will be beneficial for the disease management and prevention in the future.
1. Introduction
Breast cancer is the most common cancer among women worldwide [1,2]. There were over two million new cases in 2018. In 2015, 14,801 new cases were diagnosed in Taiwan [3]. It ranked fourth in mortality in Taiwan and led to the death of 18.2 persons per 100,000 of the population. Estrogen, predominantly 17β-estradiol (E2) and its receptor has long been known to enhance the development and progression of breast cancer. Drugs targeting the estrogen signaling pathway through the selective estrogen receptor modulator (SERM) (e.g., tamoxifen, raloxifene), the estrogen receptor (ER) antagonists (e.g., fulvestrant) and, the aromatase inhibitors, including the reversible nonsteroidal agents (e.g., letrozole, anastrozole), or the irreversible steroidal inactivator (e.g., exemestane), has been used for decades to treat ER positive breast cancer (Figure 1) [4]. Tamoxifen is the first SERM approved for the treatment of breast cancer and effectively demonstrated in the reduction of the recurrence and prevention of contralateral breast cancer. However, primary or acquired resistance frequently arises and becomes the major obstacle in hormone therapy, which indicates a more complex receptor and signaling pathways involved in the cancer progression. The G-protein coupled estrogen receptor (GPER), originally known as GPR30, a seven transmembrane domain protein, is an alternate estrogen receptor with a structure distinct from the two canonical estrogen receptors, ERα and ERβ mainly mediate a rapid non-genomic response [5,6,7,8,9,10]. This is expressed in about 50% to 60% of breast cancer tissues and has been reported as a modulator of neoplastic transformation (Figure 2) [11,12,13,14,15,16,17,18,19,20,21,22,23,24,25]. Paradoxically, the modulators or antagonists of the classical estrogen receptors such as tamoxifen, raloxifene, and fulvestrant, were found to be the GPER agonists [24]. The expression of GPER has been presumed to be associated with the development of tamoxifen resistance [26,27,28,29,30,31,32]. In breast cancer patients treated with tamoxifen, there is an increased risk of developing endometrial cancer and often has a poor clinical outcome. GPER was also supposed to mediate the contrary tissue-specific effect [33,34].
On the other hand, triple-negative breast cancer (TNBC), defined as a lack of ERα, progesterone receptor (PR), or the overexpression of human epidermal growth factor receptor 2 (HER2/neu), constitutes 15% to 20% of breast cancers. It is more prevalent in younger women and is frequently present at a more advanced stage with a more aggressive behavior. Lacking a well-defined receptor and signaling pathway, chemotherapy remains the treatment of choice but with a higher rate of recurrence. GPER is prevalent in TNBC and presumed to be involved in the growth of TNBC. It can be considered as a candidate of therapeutic target [35,36,37,38,39,40].
The endocrine disruptive chemicals, such as bisphenol (Figure 1) and thiodiphenol, at the environmentally relevant doses may exert effects through the GPER and estrogen-like signaling pathways, contribute to breast cancer progression, and drug resistance in both the ERα-positive and -negative breast cancer cells [41,42,43,44,45].
The epidermal growth factor receptor (EGFR) activation is a common and important event in the pathogenesis and progression of breast cancer. The EGFR transactivation by estrogen via the GPER has been proposed as an alternate signaling pathway with a potential significance for breast cancer [46,47,48,49,50,51,52,53,54].
However, contradictory results have existed regarding the response of GPER to estrogens/antiestrogens and the effect of GPER agonist/antagonist on the proliferation, migration and invasion of the breast cancer cells [55,56,57,58,59,60,61,62,63,64,65]. There were controversies on the subcellular localization of GPER and its function, proliferative or pro-apoptotic. Therefore, the role of GPER in ERα positive breast cancer and TNBC remains unclear.
In the following, updated evidence about the GPER in breast cancer were examined. Future epidemiology and laboratory studies, which may be helpful to elucidate the role of the GPER, were proposed.
2. G-Protein Coupled Estrogen Receptor Expression in Breast Cancer
The significance of GPER in human breast cancer was evaluated by comparing its relationship to ER, PR, and the cancer progression variables through immunohistochemical analysis [15]. A significant association between the GPER and ER was observed. GPER was positively correlated with the HER2/neu expression, tumor size, and metastasis. The distinct patterns of the GPER and ER in association with the cancer progression variable supported that the GPER and ER have independent influences on the estrogen responsiveness of breast cancer. The association between GPER expression and tamoxifen resistance was later confirmed [27]. The GPER was negatively correlated with relapse-free survival in patients only treated with tamoxifen. Multivariate analysis revealed that the GPER expression was an independent prognosticator for a poor outcome. In a study of postmenopausal lymph node negative breast cancer patients, the absence of the plasma membrane GPER predicted a 91% 20-year distant disease-free survival, as compared to 73% in the presence of GPER for the tamoxifen-treated ER-positive and PR-positive subgroup [20]. The GPER overexpression and plasma membrane localization are critical events in breast cancer progression. GPER was also prevalent in the TNBC, and the GPER expression was associated with a younger age and a more aggressive disease [37].
As compared with the corresponding primary tumors in the same patients, GPER expression in the recurrent tumors or metastases significantly increased under the tamoxifen treatment [27,28].
The tissue microarrays from the formalin-fixed, paraffin embedded samples of the primary invasive breast carcinomas suggested that the predominantly cytoplasmic or nuclear GPER expression were two distinct immunohistochemical patterns and may reflect different biological features [19]. Cytoplasmic GPER expression was associated with non-ductal histology, lower stage, more differentiation, and better overall survival, whereas the nuclear GPER expression was associated with poor differentiation and TNBC. In the breast cancer cell lines, confocal microscopy revealed the different GPER expression patterns. The T47D cells had a strong GPER expression, predominantly localized in the cytoplasm. The MCF-7 cells showed a less strong GPER expression and a mainly nuclear distribution. No distinct plasma membranous expression was observed.
Briefly, GPER was prevalent in the ERα positive breast cancer and TNBC. The GPER was a prognosticator for a poor outcome. There was a higher GPER expression in the re-biopsy specimen of tamoxifen resistant ERα positive breast cancer and chemotherapy refractory TNBC than the primary tumor. It should be noted that the subcellular location of the GPER may have a different prognostic implication in breast cancer.
3. G-Protein Coupled Estrogen Receptor Functions in Breast Cancer
The 17β-estradiol activated the extracellular regulated protein kinase 1/2 (ERK1/2), not only in the ERα-positive and the ERβ-positive MCF-7 cells, but also in the ERα-negative and the ERβ-negative SkBr3 cells [47]. Immunoblot analysis showed that this estrogen response was associated with the presence of the GPER protein in these cells. The ERα-negative, ERβ-positive MDA-MB-231 cells are GPER deficient and insensitive to ERK1/2 activation by E2. Transfection of the MDA-MB-231 cells with a GPER complementary DNA resulted in the overexpression of a GPER protein and conversion to an estrogen-responsive phenotype. In addition, the GPER-dependent ERK1/2 activation could be triggered by the ER antagonist, fulvestrant. The E2 signaling to the ERK1/2 occurred via a Gβγ-dependent, pertussis toxin-sensitive pathway. The β and γ subunits of the G protein activate the steroid receptor coactivator (SRC) tyrosine kinase, which binds to the integrin ανβ1 through the SHC adapter protein (Figure 3). The complex activates the matrix metalloproteinase, which then cleaves the pro-heparin-binding EGF-like growth factor and releases the heparin-binding EGF-like growth factor (HB-EGF) into the extracellular space. The free HB-EGF then transactivates the EGFR. The E2 signaling to the ERK1/2 could be blocked by down-modulating the HB-EGF from the cell surface with the diphtheria toxin mutant, CRM-197, neutralizing the HB-EGF with antibodies, or inhibiting the EGFR tyrosine kinase activity. ER-negative breast cancers that continue to express GPER may use estrogen to drive the EGFR-dependent cellular responses [47,50,51]. The crosstalk between the GPER and the EGFR was confirmed in the tamoxifen-resistant ERα positive cell, TNBC, and cancer-associated fibroblast (CAF), respectively (Table 1) [26,28,38,66,67].
3.1. Tamoxifen-Resistant ERα Positive Cells
The tamoxifen-resistant cells, TAM-R, exhibited an enhanced sensitivity to the E2 and the GPER-specific agonist, G1 (Figure 1), when compared to the parental MCF-7 cells [26,28]. Tamoxifen was able to stimulate the mitogen-activated protein kinase (MAPK) phosphorylation and cell growth in the TAM-R cells, and the effects were abolished by the GPER antagonist, G15 (Figure 1), the GPER anti-sense oligonucleotide, the selective SRC inhibitor PP2, and the EGFR inhibitor AG1478. The basal EGFR expression was only slightly elevated in the TAM-R cells, and the basal GPER expression, phosphorylation of the Ak strain transforming murine thymoma viral oncogene (AKT), and the MAPK remained unchanged when compared to the parental cells. Continuous treatment of the MCF-7 cells with G1 mimics the long-term treatment with tamoxifen and drastically increases its agonistic activity. Interestingly, the estrogen treatment significantly increased the GPER plasma membrane translocation, which was stronger in the TAM-R cells. The GPER plasma membrane translocation facilitated the crosstalk with the EGFR. The results have suggested the importance of the GPER and EGFR transactivation in the development of tamoxifen resistance. Combined therapy with the G15 and tamoxifen promoted apoptosis in a TAM-R xenograft and inhibited the drug-resistant tumor progression. The GPER activation led to the nuclear translocation of the forkhead box O3a, FOXO3a, and the down-regulation of caspase 3 and caspase 7 via the phosphoinositide 3-kinase (PI3K)/AKT pathway in the MCF-7 cells [52].
3.2. Triple-Negative Breast Cancer Cells
The GPER was strongly expressed in the TNBC cell lines, MDA-MB-468, and MDA-MB-436 [38]. Treatment with E2, tamoxifen, and the GPER-specific agonist, G1 led to the rapid activation of ERK1/2, but not AKT. The estrogen/GPER/ERK signaling pathway was involved in the increased cell growth, survival, migration, and invasion through upregulating the expression of cyclin A, cyclin D1, Bcl-2, and c-fos that were associated with the cell cycle, anti-apoptosis, and proliferation, respectively. Pretreatment with the GPER antagonist, G15, AG1478, the ERK1/2 inhibitor, U0126, or the transfection with the siRNA against the GPER could abolish the effects. Immunohistochemical analysis of the TNBC specimens showed a significantly stronger staining of the p-ERK1/2 in the GPER-positive tissues than the GPER-negative tissues [38]. The positivity of the GPER and p-ERK1/2 displayed a strong association with the large tumor size and advanced stage, indicating that the GPER/ERK signaling might also contribute to the tumor progression in the TNBC patients, which correlated with the in vitro experimental results. 17β-estradiol and 4-hydroxytamoxifen also increased the proliferation of another two TNBC cell lines, MDA-MB-435 and HCC1806, which was completely prevented by being transfected with siRNA against the GPER [36]. The increased activity of the SRC kinase, EGFR transactivation, and c-fos expression, was also abolished after the knock-down of the GPER expression.
3.3. Cancer-Associated Fibroblasts
The GPER was expressed in the stromal fibroblasts of the primary breast cancer tissues, and the CAFs isolated [66,67]. Tamoxifen, in addition to E2 and the G1 activated GPER, resulted in the transient increases in the cell index, intracellular calcium, ERK1/2 phosphorylation, and promoted the CAF cell cycle progression, proliferation, and migration. These effects were blocked by the G15, AG1478, and U0126. Importantly, tamoxifen, as well as G1, increased the E2 production in the breast CAFs via the GPER/EGFR/ERK signaling pathway when the substrate of E2, testosterone, was added to the medium. The GPER-mediated CAF-dependent estrogenic effects in the tumor-associated stroma are more likely to contribute to breast cancer progression, especially in the tamoxifen resistance, via a positive feedback loop involving the GPER/EGFR/ERK signaling pathway and E2 production.
17β-estradiol and G1 triggered the GPER/EGFR/ERK/c-fos signaling pathway that led to an increased vascular endothelial growth factor (VEGF) via the upregulation of the hypoxia-inducible factor-1α (HIF1α) in the ER-negative breast cancer cells and the CAFs [53]. The conditioned medium from the CAFs treated with E2 and G1 promoted the human endothelial tube formation in a GPER-dependent manner. In the mice breast cancer xenograft model, GPER activation enhanced the tumor growth and the expression of HIF1α, VEGF, and the endothelial marker, CD34. Fatty acid synthase catalyzes the de novo biogenesis of the fatty acids and acts as a metabolic oncogene. 17β-estradiol and G1 regulated the fatty acid synthase expression and activity through the GPER/EGFR/ERK/c-fos/activator protein 1 (AP-1) signaling pathway in the SkBr3 cells and CAFs [54].
In summary, E2, tamoxifen, and G1 upregulate estrogen production, increase GPER expression and plasma membrane translocation, and stimulate the proliferation, migration, invasion of breast cancer cell lines, and cancer-associated fibroblasts. Pretreatment with the GPER antagonist, G15 or transfected with siRNA against GPER attenuates the effects. The tumor promoting effects of the GPER operate through the EGFR transactivation and related signaling pathways.
3.4. Controversies about G-Protein Coupled Estrogen Receptor Function in Breast Cancer
Contrary to the majority of studies that have reported a tumor promoting effect of GPER activation in breast cancer, several studies have demonstrated that GPER functions as a tumor suppressor and induces apoptosis (Table 1) [55,56,57,58,59,60,61,62,63,64,65].
The GPER expression by immunohistochemistry had been reported as a prognosticator for the increased distant disease-free survival in patients with ER-positive breast cancer treated with tamoxifen [55]. A constitutive GPER-dependent pro-apoptotic signaling was proposed. The GPER expression at mRNA levels was significantly down-regulated in both the ERα-positive and ERα-negative breast cancer tissues in comparison with their matched normal tissues, and significantly lower in tumor tissues from the patients who had lymph node metastasis than those without [56]. The tumor samples from 118 Taiwanese patients with infiltrating ductal carcinoma of the breast had a lower GPER expression at the mRNA level than that in non-tumor mammary tissues [57]. The correlation of the GPER expression with clinical parameters and patient survival was not significant.
Filardo et al., suggested that via the GPER, estrogens as well as antiestrogens, were capable of stimulating the adenylyl cyclase activity and increasing the cAMP concentration, which in turn, led to the PKA-mediated suppression of the EGFR induced ERK1/2 activity (Figure 3) [58]. Thus, via the GPER, estrogen may balance ERK1/2 activity by stimulating two distinct G-protein signaling pathways that have opposing effects on the EGFR-to-MAPK axis. The other study concurred with the observation that the reduced cAMP generation attenuated the inhibition of EGFR signaling [28].
The GPER functions promoted the SkBr-3 but inhibited the MCF-7 cellular proliferation. An ER- and [Ca2+]-dependent negative feedback was proposed for the difference [59]. 17β-estradiol is known to downregulate the ERα expression in the MCF-7 cells as a negative feedback regulatory loop to prevent overexpression. Likewise, the GPER may also be negatively regulated by E2 via the ER to prevent an excessive GPER-dependent activity, such as being aberrantly high [Ca2+]. The maximum increases in [Ca2+] were much larger in the SkBr-3 cells than in the MCF-7 cells. It is possible that this was due to the lack of ERs in the SkBr-3 cells, which translated into a lack of negative feedback regulation.
Proliferative results were observed with two non-specific GPER agonists, estrogen and tamoxifen, but not with its specific agonist, G1. The G1-induced inhibitory effect was specific for the GPER expressing cells [60,61]. Radiation induced different changes in the GPER expression among the breast cancer cells; upregulated in the MDA-MB-231 and the MDA-MB-468 cells and down-regulated in the MCF-7 cells. They proposed that it was linked to the form of the p53 protein expressed in each cell. While the MDA-MB-231 and the MDA-MB-468 cells express a non-functional p53 gene, the MCF-7 cells have normal wild-type p53. A feedback mechanism exists between the expression of p53 and GPER. The GPER activation increases the p53 expression, which in turn down-regulates the expression of the GPER [61].
The activation of the GPER suppresses the epithelial mesenchymal transition, migration, and angiogenesis of the TNBC via the nuclear factor-kappa B (NF-κB) signals [62,63]. The GPER also inhibits the tumor necrosis factor alpha-induced expression of interleukin 6 through the repression of the NF-κB promoter activity in the SkBr3 cells [64].
Further studies are therefore necessary to define the role of the GPER, proliferative or pro-apoptotic in breast cancer.
4. G-protein Coupled Estrogen Receptor Knockout Mice
In contrast with the pharmacological methods, the GPER knockout more clearly understands the GPER function through targeted gene deletion or disruption [17]. Four GPER knockout mice have been reported [68,69,70,71]. Among them, three have the whole GPER coding region deleted, and the fourth with the C-terminal portion of the GPER remaining. Although the GPER was also distributed in normal breast tissues, histopathological analysis did not reveal any abnormalities in the GPER-knockout mice [68]. Mammary gland responses after estradiol with/without progesterone treatment were also unimpaired in the GPER knockout mice. The GPER knockout mice did not show overt phenotypes in viability or reproductive function, but some functions of estrogen have been absent in the GPER knockout mice that support the GPER as a physiologically relevant estrogen receptor [17,68,69,70,71,72]. In the only study of breast cancer, the GPER knockout in the polyoma middle T antigen-mouse mammary tumor virus transgenic mice revealed smaller tumors and reduced metastasis [18].
5. Interactions between Cancer Cells and Cancer-Associated Fibroblasts through G-Protein Coupled Estrogen Receptor
An increased aromatase expression and activity was found in the tamoxifen resistant breast cancer cells [30]. Knocking-down the GPER expression reversed the enhanced aromatase levels. In the ER-negative, GPER/aromatase-positive SkBr3 cells, tamoxifen acted as a GPER agonist. The tamoxifen treatment increased the aromatase expression through an enhanced recruitment of the c-fos/c-jun complex to the AP-1 responsive elements located within the promoter region. Tamoxifen also induced aromatase expression via the GPER in the CAFs. The increased estrogen production in the microenvironment may well lead to a more aggressive behavior of breast cancers.
On co-culturing the CAFs with the breast cancer cells, a significant GPER translocation from the nucleus to the cytoplasm was observed in the CAFs, similar to that observed in the stromal fibroblast in the breast cancer tissues, indicating that the cancer cells may affect the subcellular localization of the GPER in the CAFs. CRM1, a nuclear export protein, and activated PI3K/AKT signaling pathway are involved in the cytoplasmic GPER translocation in the CAFs, which in turn activates a novel estrogen/GPER/cyclic AMP (cAMP)/protein kinase A (PKA)/cAMP response element binding protein signaling pathway, and triggers aerobic glycolysis in the CAFs [73]. The glycolytic CAFs feed pyruvate and lactate to cancer cells to undergo oxidative phosphorylation and contribute to drug resistance. The stromal GPER-mediated drug resistance from the reprogramming of the tumor energy metabolism, i.e. the “reverse Warburg effect”, provided the rationale for the CAFs as a promising target for therapy [74,75]. The different subcellular location of the GPER in the breast CAFs may have biological implications [76]. Targeting the cytoplasmic GPER in the CAFs may restore the response to treatments in both the ER-positive and -negative breast cancers.
6. Future Perspectives in the Study of the G-Protein Coupled Estrogen Receptor in Breast Cancer
The inconsistent observations among these studies could be attributed to the usage of the different subtypes of breast cancer samples and the different subcellular localization of the GPER [60], the difference of the cell types and treatment conditions, and the specificities of the agonist [61]. The specificity of the GPER antibodies used in the immunohistochemistry and Western blot may affect the staining patterns of the GPER [77]. The epigenetic of GPER, tumor microenvironment, and hormone levels also affected the results [65].
Systemic approaches via epidemiology and laboratory studies are necessary to confirm the role of GPER in breast cancer in the future. ERα positive breast cancer and TNBC need to be studied separately. What is the true prevalence of GPER in the ERα positive breast cancer and the TNBC, respectively? The GPER expression of the archival tissue in both tumor and associated stroma should be measured by a unified scoring system including the staining intensity and the proportion of positive cancer cells, as per the Allred scoring for ER [78]. The subcellular location of the GPER, i.e. nucleus, cytoplasm, and plasma membrane, also need be notified to clarify the implication of the translocation and its role in the prognosis.
The GPER expression needs to be compared between the tumors and their matched normal tissues. The association of GPER with the ERα, PR, HER2/neu, and the correlations with the clinic-pathologic variables and survival also need to be investigated. In addition to stage, histology, and ER, PR, HER2/neu status, the smoking history, and menopausal status should be included in a multivariate survival analysis to determine as to whether the GPER is an independent prognosticator. Does high or low GPER expression before treatment predict the development of tamoxifen resistance or refractory TNBC? Pairwise comparison of the GPER expression by immunohistochemistry between the archival tissues of the treatment-naïve and re-biopsy specimen of tamoxifen resistant ERα positive breast cancer or the chemotherapy refractory TNBC, will help to better understand whether the GPER becomes predominant during treatment.
The baseline GPER expression in the ERα positive, tamoxifen-resistant ERα positive, TNBC cell lines, and the CAFs, their differences including the subcellular location, the estrogen production including the aromatase activity, the subsequent GPER expression, and the effects, i.e. proliferation, migration, invasion after treatment with E2, tamoxifen and G1, and whether the effects are abolished by pretreatment with G15 or siRNA against the GPER will help to confirm the role of GPER. The differences between the effects of E2, tamoxifen and G1 should also be observed for the receptor specificity. Immunofluorescent microscopy may be used to observe the cell surface translocation of the GPER, which facilitates the crosstalk with EGFR. In addition to the ERK signaling pathway, is the AKT pathway involved in the GPER-EGFR transactivation [38]? The reciprocal changes of the GPER expression, the biomarkers for the “reverse Warburg effect”, and the lactate shuttle such as the mono-carboxylate transporters 4, and the mitochondrial activities in the cancer cell lines and the CAFs, on treatment with E2, tamoxifen, or G1 also deserve to be studied, to explore the role of the metabolic coupling that occurs between the CAFs and the cancer cells.
For the reported anti-proliferative and pro-apoptotic effects of the GPER activation in the literature, the stimulation of the adenylyl cyclase to increase the intracellular cAMP or intracellular Ca2+ mobilization as a second messenger, and the pro-apoptotic signaling, measured by the increased cytochrome C release, caspase-3 cleavage, poly(ADP-ribose) polymerase, PARP cleavage, and the decreased cell viability after treatment with estrogen, tamoxifen, G1, or G15, could be correlated to understand the regulatory mechanism [58]. In contrast with the chronic exposure at a low nanomolar dosage, the higher bisphenol concentration in the micromolar exerts an anti-proliferative effect on the cancer cells in spite of the activation of the EGFR/ERK signaling pathway. An increased expression of p53 and its phosphorylation was observed. Therefore, we need to investigate whether the GPER effects are modified by the simultaneous p53 activation [79].
Several other signaling pathways, such as HIPPO, NOTCH, and target genes and proteins, such as SNAIL, β1-integrin, focal adhesion kinase, calpain, and connective tissue growth factor, have been reported to be involved in the GPER-mediated breast cancer progression [29,80,81,82,83]. The microRNAs target numerous genes and are involved in cancer progression. GPER is an important regulator of microRNAs. In the MCF-7 and MDA-MB-231 cells, the GPER activation down-regulated the miR-148a and caused an increase in the human leukocyte antigen-C expression that led to the cancer cells escaping from immune surveillance and allowed cancer progression [84]. In the SkBr-3 cells and CAFs, the GPER activation up-regulated the miR-144, which in turn inhibited the tumor suppressor runt-related transcription 1 factor, and increased the cell cycle propagation [85]. Systemic studies of microRNA expression are needed to better define the regulations by the GPER and their effects.
To include GPER in the functional screens for genes contributing to tamoxifen resistance in breast cancer cells and the usage of novel technology, e.g., DNA microarray, and proteomic analysis may help to investigate the association [86,87]. Insulin-like growth factor-I and HIF1α were shown to increase the GPER expression in the breast cancer cells and led to cell proliferation, migration, and tumor angiogenesis [88,89]. The epidermal growth factor may reciprocally up-regulate the GPER to facilitate a stimulatory role of estrogen, even in the TNBC [40]. How the GPER expression is regulated, e.g., through epigenetic by methylation or demethylation of the promoter, requires further studies to understand the EGFR and GPER crosstalk [61,90].
The classical ERs do not contain a hydrophobic part that may serve as a transmembrane domain. However, the presence of the ERs in the membrane of the somatic and cancer cells, and the rapid non-genomic responses that occur through the membrane-bound classical ERα and ERβ have been reported [49,91,92,93]. How the classical ERs translocate to the membrane [94], and the interactions between the classical ERs and the GPER is important to understand the membrane-associated non-genomic pathways of estrogen.
7. Conclusions
In this review, we have tried to explore the actions and to understand the molecular basis of the agonist/antagonist mechanisms of the GPER in breast cancer with tamoxifen resistance, and TNBC from the current epidemiology and laboratory studies. The combined non-genomic and genomic effects of estrogen are critical for its overall function, even in the absence of ligand, and the interactions between multiple receptors are complex [95]. Further studies will help to clarify the role of the GPER and support it as a novel target of therapeutic strategies. GPER expression may also be valuable as prognostic or predictive biomarkers.
Author Contributions
L.-H.H. and S.-H.K. conceived the paper; L.-H.H. and S.-H.K. wrote the paper; L.-H.H., N.-M.C., Y.-F.L., and S.-H.K. revised the paper.
Funding
This research received no external funding.
Acknowledgments
The authors would like to offer their sincere thanks to Michael Wise for help with English language editing; cytotechnologists Jen-Sheng Ko and Mei-Ling Wu for the figures preparation; Shiao-Chiu Huang for the assistance with the references preparation.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AKT | Ak strain transforming murine thymoma viral oncogene |
| AP-1 | activator protein 1 |
| CAF | cancer-associated fibroblast |
| cAMP | cyclic AMP |
| E2 | 17β-estradiol |
| EGFR | epidermal growth factor receptor |
| ER | estrogen receptor |
| ERα | estrogen receptor α |
| ERβ | estrogen receptor β |
| ERK | extracellular regulated protein kinase |
| GPER | G-protein coupled estrogen receptor |
| HB-EGF | heparin-binding EGF-like growth factor |
| HER2/neu | human epidermal growth factor receptor 2 |
| HIF1α | hypoxia-inducible transcription factor-1α |
| MAPK | mitogen-activated protein kinase |
| NF-κB | nuclear factor-kappa B |
| PI3K | Phosphoinositide 3-kinase |
| PKA | protein kinase A |
| PR | progesterone receptor |
| SERM | selective estrogen receptor modulator |
| SRC | steroid receptor coactivator |
| TNBC | triple-negative breast cancer |
| VEGF | vascular endothelial growth factor |
References
- Kamangar, F.; Dores, G.M.; Anderson, W.F. Patterns of cancer incidence, mortality, and prevalence across five continents: Defining priorities to reduce cancer disparities in different geographic regions of the world. J. Clin. Oncol. 2006, 24, 2137–2150. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Health Promotion Administration, Ministry of Health and Welfare, Taiwan Cancer Registry Annual Report, 2015, Taiwan. Available online: http://www.hpa.gov.tw (accessed on 21 November 2018).
- NCCN Clinical Practice Guidelines in Oncology: Breast Cancer Risk Reduction. Available online: http://www.nccn.org (accessed on 21 November 2018).
- Carmeci, C.; Thompson, D.A.; Ring, H.Z.; Francke, U.; Weigel, R.J. Identification of a gene (GPR30) with homology to the G-protein-coupled receptor superfamily associated with estrogen receptor expression in breast cancer. Genomics 1997, 45, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Pang, Y.; Filardo, E.J.; Dong, J. Identity of an estrogen membrane receptor coupled to a G protein in human breast cancer cells. Endocrinology 2005, 146, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.; Quinn, J.; Pang, Y.; Graeber, C.; Shaw, S.; Dong, J.; Thomas, P. Activation of the novel estrogen receptor G protein-coupled receptor 30 (GPR30) at the plasma membrane. Endocrinology 2007, 148, 3236–3245. [Google Scholar] [CrossRef]
- Dennis, M.K.; Burai, R.; Ramesh, C.; Petrie, W.K.; Alcon, S.N.; Nayak, T.K.; Bologa, C.G.; Leitao, A.; Brailoiu, E.; Deliu, E.; et al. In vivo effects of a GPR30 antagonist. Nat. Chem. Biol. 2009, 5, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Pupo, M.; Maggiolini, M.; Musti, A.M. GPER mediates non-genomic effects of estrogen. Methods Mol. Biol. 2016, 1366, 471–488. [Google Scholar]
- Rae, J.M.; Johnson, M.D. What does an orphan G-protein-coupled receptor have to do with estrogen? Breast Cancer Res. 2005, 7, 243–244. [Google Scholar] [CrossRef]
- Lappano, R.; Rosano, C.; Santolla, M.F.; Pupo, M.; De Francesco, E.M.; De Marco, P.; Ponassi, M.; Spallarossa, A.; Ranise, A.; Maggiolini, M. Two novel GPER agonists induce gene expression changes and growth effects in cancer cells. Curr. Cancer Drug Targets 2012, 12, 531–542. [Google Scholar] [CrossRef]
- Méndez-Luna, D.; Bello, M.; Correa-Basurto, J. Understanding the molecular basis of agonist/antagonist mechanism of GPER1/GPR30 through structural and energetic analyses. Steroid Biochem. Mol. Biol. 2016, 158, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Aiello, F.; Carullo, G.; Giordano, F.; Spina, E.; Nigro, A.; Garofalo, A.; Tassini, S.; Costantino, G.; Vincetti, P.; Bruno, A.; et al. Identification of breast cancer inhibitors specific for G protein-coupled estrogen receptor (GPER)-expressing cells. ChemMedChem 2017, 12, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Graeber, C.T.; Quinn, J.A.; Resnick, M.B.; Giri, D.; DeLellis, R.A.; Steinhoff, M.M.; Sabo, E. Distribution of GPR30, a seven membrane-spanning estrogen receptor, in primary breast cancer and its association with clinicopathologic determinants of tumor progression. Clin. Cancer Res. 2006, 12, 6359–6366. [Google Scholar] [CrossRef] [PubMed]
- Marot, D.; Bieche, I.; Aumas, C.; Esselin, S.; Bouquet, C.; Vacher, S.; Lazennec, G.; Perricaudet, M.; Kuttenn, F.; Lidereau, R.; et al. High tumoral levels of Kiss1 and G-protein-coupled receptor 54 expression are correlated with poor prognosis of estrogen receptor-positive breast tumors. Endocr. Relat. Cancer 2007, 14, 691–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prossnitz, E.R.; Hathaway, H.J. What have we learned about GPER function in physiology and disease from knockout mice? J. Steroid Biochem. Mol. Biol. 2015, 153, 114–126. [Google Scholar] [CrossRef] [Green Version]
- Marjon, N.A.; Hu, C.; Hathaway, H.J.; Prossnitz, E.R. G protein-coupled estrogen receptor regulates mammary tumorigenesis and metastasis. Mol. Cancer Res. 2014, 12, 1644–1654. [Google Scholar] [CrossRef]
- Samartzis, E.P.; Noske, A.; Meisel, A.; Varga, Z.; Fink, D.; Imesch, P. The G protein-coupled estrogen receptor (GPER) is expressed in two different subcellular localizations reflecting distinct tumor properties in breast cancer. PLoS ONE 2014, 9, e83296. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, M.; Hartman, L.; Grabau, D.; Fornander, T.; Malmström, P.; Nordenskjöld, B.; Sgroi, D.C.; Skoog, L.; Stål, O.; Leeb-Lundberg, L.M.; et al. Lack of G protein-coupled estrogen receptor (GPER) in the plasma membrane is associated with excellent long-term prognosis in breast cancer. Breast Cancer Res. Treat. 2014, 145, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J. A role for G-protein coupled estrogen receptor (GPER) in estrogen-induced carcinogenesis: Dysregulated glandular homeostasis, survival and metastasis. J. Steroid Biochem. Mol. Biol. 2018, 176, 38–48. [Google Scholar] [CrossRef]
- Wang, D.; Hu, L.; Zhang, G.; Zhang, L.; Chen, C. G protein-coupled receptor 30 in tumor development. Endocrine 2010, 38, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Lappano, R.; Pisano, A.; Maggiolini, M. GPER function in breast cancer: An overview. Front. Endocrinol. 2014, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Jacenik, D.; Cygankiewicz, A.I.; Krajewska, W.M. The G protein-coupled estrogen receptor as a modulator of neoplastic transformation. Mol. Cell Endocrinol. 2016, 429, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Molina, L.; Figueroa, C.D.; Bhoola, K.D.; Ehrenfeld, P. GPER-1/GPR30 a novel estrogen receptor sited in the cell membrane: Therapeutic coupling to breast cancer. Expert Opin. Ther. Targets 2017, 21, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, A.; Ignatov, T.; Roessner, A.; Costa, S.D.; Kalinski, T. Role of GPR30 in the mechanisms of tamoxifen resistance in breast cancer MCF-7 cells. Breast Cancer Res. Treat. 2010, 123, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, A.; Ignatov, T.; Weissenborn, C.; Eggemann, H.; Bischoff, J.; Semczuk, A.; Roessner, A.; Costa, S.D.; Kalinski, T. G-protein-coupled estrogen receptor GPR30 and tamoxifen resistance in breast cancer. Breast Cancer Res. Treat. 2011, 128, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Liu, M.; Yang, F.; Luo, H.; Li, Z.; Tu, G.; Yang, G. GPR30 as an initiator of tamoxifen resistance in hormone-dependent breast cancer. Breast Cancer Res. 2013, 15, R114. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Z.; He, Y.; Shang, D.; Pan, J.; Wang, H.; Chen, H.; Zhu, Z.; Wan, L.; Wang, X. Estrogen and pure antiestrogen fulvestrant (ICI 182 780) augment cell-matrigel adhesion of MCF-7 breast cancer cells through a novel G protein coupled estrogen receptor (GPR30)-to-calpain signaling axis. Toxicol. Appl. Pharmacol. 2014, 275, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Catalano, S.; Giordano, C.; Panza, S.; Chemi, F.; Bonofiglio, D.; Lanzino, M.; Rizza, P.; Romeo, F.; Fuqua, S.A.; Maggiolini, M.; et al. Tamoxifen through GPER upregulates aromatase expression: A novel mechanism sustaining tamoxifen-resistant breast cancer cell growth. Breast Cancer Res. Treat. 2014, 146, 273–285. [Google Scholar] [CrossRef]
- Nass, N.; Kalinski, T. Tamoxifen resistance: From cell culture experiments towards novel biomarkers. Pathol. Res. Pract. 2015, 211, 189–197. [Google Scholar] [CrossRef]
- Rondón-Lagos, M.; Villegas, V.E.; Rangel, N.; Sánchez, M.C.; Zaphiropoulos, P.G. Tamoxifen resistance: Emerging molecular targets. Int. J. Mol. Sci. 2016, 17, 1357. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, T.; Eggemann, H.; Semczuk, A.; Smith, B.; Bischoff, J.; Roessner, A.; Costa, S.D.; Kalinski, T.; Ignatov, A. Role of GPR30 in endometrial pathology after tamoxifen for breast cancer. Am. J. Obstet. Gynecol. 2010, 203, 595. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Hilakivi-Clarke, L.; Clarke, R. Molecular mechanisms of tamoxifen-associated endometrial cancer. Oncol. Lett. 2015, 9, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Lappano, R.; Rosano, C.; De Marco, P.; De Francesco, E.M.; Pezzi, V.; Maggiolini, M. Estriol acts as a GPR30 antagonist in estrogen receptor-negative breast cancer cells. Mol. Cell Endocrinol. 2010, 320, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Girgert, R.; Emons, G.; Gründker, C. Inactivation of GPR30 reduces growth of triple-negative breast cancer cells: Possible application in targeted therapy. Breast Cancer Res. Treat. 2012, 134, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Steiman, J.; Peralta, E.A.; Louis, S.; Kamel, O. Biology of the estrogen receptor, GPR30, in triple negative breast cancer. Am. J. Surg. 2013, 206, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Liu, M.; Luo, H.; Wu, C.; Tang, X.; Tang, S.; Hu, P.; Yan, Y.; Wang, Z.; Tu, G. GPER mediates enhanced cell viability and motility via non-genomic signaling induced by 17β-estradiol in triple-negative breast cancer (TNBC) cells. J. Steroid Biochem. Mol. Biol. 2014, 143, 392–403. [Google Scholar] [CrossRef]
- Zhou, K.; Sun, P.; Zhang, Y.; You, X.; Li, P.; Wang, T. Estrogen stimulated migration and invasion of estrogen receptor-negative breast cancer cells involves an ezrin-dependent crosstalk between G protein-coupled receptor 30 and estrogen receptor beta signaling. Steroids 2016, 111, 113–120. [Google Scholar] [CrossRef]
- Albanito, L.; Sisci, D.; Aquila, S.; Brunelli, E.; Vivacqua, A.; Madeo, A.; Lappano, R.; Pandey, D.P.; Picard, D.; Mauro, L.; et al. Epidermal growth factor induces G protein-coupled receptor 30 expression in estrogen receptor-negative breast cancer cells. Endocrinology 2008, 149, 3799–3808. [Google Scholar] [CrossRef]
- Lapensee, E.W.; Tuttle, T.R.; Fox, S.R.; Ben-Jonathan, N. Bisphenol A at low nanomolar doses confers chemoresistance in estrogen receptor-alpha-positive and -negative breast cancer cells. Environ. Health Perspect. 2009, 117, 175–180. [Google Scholar] [CrossRef]
- Dong, S.; Terasaka, S.; Kiyama, R. Bisphenol A induces a rapid activation of Erk1/2 through GPR30 in human breast cancer cells. Environ. Pollut. 2011, 159, 212–218. [Google Scholar] [CrossRef]
- Yu, X.; Filardo, E.J.; Shaikh, Z.A. The membrane estrogen receptor GPR30 mediates cadmium-induced proliferation of breast cancer cells. Toxicol. Appl. Pharmacol. 2010, 245, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, T.; Yang, P.; Li, M.; Yang, Y.; Wang, Y.; Du, J.; Pan, K.; Zhang, K. Low doses of bisphenol A stimulate the proliferation of breast cancer cells via ERK1/2/ERRγ signals. Toxicol. Vitro 2015, 30, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Peng, W.; Xu, G.; Wu, M.; Wen, Y.; Xu, J.; Yu, Z.; Wang, Y. Activation of G protein-coupled receptor 30 by thiodiphenol promotes proliferation of estrogen receptor α-positive breast cancer cells. Chemosphere 2017, 169, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Biscardi, J.S.; Ishizawar, R.C.; Silva, C.M.; Parsons, S.J. Tyrosine kinase signalling in breast cancer: Epidermal growth factor receptor and c-Src interactions in breast cancer. Breast Cancer Res. 2000, 2, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R., Jr. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J. Epidermal growth factor receptor (EGFR) transactivation by estrogen via the G-protein-coupled receptor, GPR30: A novel signaling pathway with potential significance for breast cancer. J. Steroid Biochem. Mol. Biol. 2002, 80, 231–238. [Google Scholar] [CrossRef]
- Levin, E.R. Bidirectional signaling between the estrogen receptor and the epidermal growth factor receptor. Mol. Endocrinol. 2003, 17, 309–317. [Google Scholar] [CrossRef]
- Filardo, E.J.; Thomas, P. GPR30: A seven-transmembrane-spanning estrogen receptor that triggers EGF release. Trends Endocrinol. Metab. 2005, 16, 362–367. [Google Scholar] [CrossRef]
- Quinn, J.A.; Graeber, C.T.; Frackelton, A.R., Jr.; Kim, M.; Schwarzbauer, J.E.; Filardo, E.J. Coordinate regulation of estrogen-mediated fibronectin matrix assembly and epidermal growth factor receptor transactivation by the G protein-coupled receptor, GPR30. Mol. Endocrinol. 2009, 23, 1052–1064. [Google Scholar] [CrossRef]
- Zekas, E.; Prossnitz, E.R. Estrogen-mediated inactivation of FOXO3a by the G protein-coupled estrogen receptor GPER. BMC Cancer 2015, 15, 702. [Google Scholar] [CrossRef]
- De Francesco, E.M.; Pellegrino, M.; Santolla, M.F.; Lappano, R.; Ricchio, E.; Abonante, S.; Maggiolini, M. GPER mediates activation of HIF1α/VEGF signaling by estrogens. Cancer Res. 2014, 74, 4053–4064. [Google Scholar] [CrossRef] [PubMed]
- Santolla, M.F.; Lappano, R.; De Marco, P.; Pupo, M.; Vivacqua, A.; Sisci, D.; Abonante, S.; Iacopetta, D.; Cappello, A.R.; Dolce, V.; et al. G protein-coupled estrogen receptor mediates the up-regulation of fatty acid synthase induced by 17β-estradiol in cancer cells and cancer-associated fibroblasts. J. Biol.Chem. 2012, 287, 43234–43245. [Google Scholar] [CrossRef] [PubMed]
- Broselid, S.; Cheng, B.; Sjöström, M.; Lövgren, K.; Klug-De Santiago, H.L.; Belting, M.; Jirström, K.; Malmström, P.; Olde, B.; Bendahl, P.O.; et al. G protein-coupled estrogen receptor is apoptotic and correlates with increased distant disease-free survival of estrogen receptor-positive breast cancer patients. Clin. Cancer Res. 2013, 19, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Poola, I.; Abraham, J.; Liu, A.; Marshalleck, J.J.; Dewitty, R.L. The cell surface estrogen receptor, G protein-coupled receptor 30 (GPR30), is markedly down regulated during breast tumorigenesis. Breast Cancer 2008, 1, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.H.; Chang, L.Y.; Liu, D.L.; Hwa, H.L.; Lin, J.J.; Lee, P.H.; Chen, C.N.; Lien, H.C.; Yuan, R.H.; Shun, C.T.; et al. The interactions between GPR30 and the major biomarkers in infiltrating ductal carcinoma of the breast in an Asian population. Taiwan J. Obstet. Gynecol. 2007, 46, 135–145. [Google Scholar] [CrossRef]
- Filardo, E.J.; Quinn, J.A.; Frackelton, A.R., Jr.; Bland, K.I. Estrogen action via the G protein-coupled receptor, GPR30: Stimulation of adenylyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptor-to-MAPK signaling axis. Mol. Endocrinol. 2002, 16, 70–84. [Google Scholar] [CrossRef]
- Ariazi, E.A.; Brailoiu, E.; Yerrum, S.; Shupp, H.A.; Slifker, M.J.; Cunliffe, H.E.; Black, M.A.; Donato, A.L.; Arterburn, J.B.; Oprea, T.I.; et al. The G protein-coupled receptor GPR30 inhibits proliferation of estrogen receptor-positive breast cancer cells. Cancer Res. 2010, 70, 1184–1194. [Google Scholar] [CrossRef]
- Weißenborn, C.; Ignatov, T.; Poehlmann, A.; Wege, A.K.; Costa, S.D.; Zenclussen, A.C.; Ignatov, A. GPER functions as a tumor suppressor in MCF-7 and SK-BR-3 breast cancer cells. J. Cancer Res. Clin. Oncol. 2014, 140, 663–671. [Google Scholar] [CrossRef]
- Weißenborn, C.; Ignatov, T.; Ochel, H.J.; Costa, S.D.; Zenclussen, A.C.; Ignatova, Z.; Ignatov, A. GPER functions as a tumor suppressor in triple-negative breast cancer cells. J. Cancer Res. Clin. Oncol. 2014, 140, 713–723. [Google Scholar] [CrossRef]
- Chen, Z.J.; Wei, W.; Jiang, G.M.; Liu, H.; Wei, W.D.; Yang, X.; Wu, Y.M.; Liu, H.; Wong, C.K.; Du, J.; et al. Activation of GPER suppresses epithelial mesenchymal transition of triple negative breast cancer cells via NF-κB signals. Mol. Oncol. 2016, 10, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Chen, Z.; Jiang, G.; Zhou, Y.; Liu, Q.; Su, Q.; Wei, W.; Du, J.; Wang, H. Activation of GPER suppresses migration and angiogenesis of triple negative breast cancer via inhibition of NF-κB/IL-6 signals. Cancer Lett. 2017, 386, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Mizukami, Y. GPER negatively regulates TNFα-induced IL-6 production in human breast cancer cells via NF-κB pathway. Endocr. J. 2016, 63, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Xuan, J.; Liu, Y.; Shi, G. Function of G-protein-coupled estrogen receptor-1 in reproductive system tumors. J. Immunol. Res. 2016, 2016, 7128702. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Yang, G.; Yu, T.; Luo, S.; Wu, C.; Sun, Y.; Liu, M.; Tu, G. GPER-mediated proliferation and estradiol production in breast cancer-associated fibroblasts. Endocr. Relat. Cancer 2014, 21, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Liu, M.; Luo, S.; Yu, T.; Wu, C.; Yang, G.; Tu, G. Dynamic monitoring of GPER-mediated estrogenic effects in breast cancer associated fibroblasts: An alternative role of estrogen in mammary carcinoma development. Steroids 2016, 112, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Fuchs, I.; Kauselmann, G.; Kern, H.; Zevnik, B.; Andreasen, P.; Schwarz, G.; Altmann, H.; Klewer, M.; Schoor, M.; et al. GPR30 does not mediate estrogenic responses in reproductive organs in mice. Biol. Reprod. 2009, 80, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Martensson, U.E.; Salehi, S.A.; Windahl, S.; Gomez, M.F.; Sward, K.; Daszkiewicz-Nilsson, J.; Wendt, A.; Andersson, N.; Hellstrand, P.; Grande, P.O.; et al. Deletion of the G protein-coupled receptor 30 impairs glucose tolerance, reduces bone growth, increases blood pressure, and eliminates estradiol-stimulated insulin release in female mice. Endocrinology 2009, 150, 687–698. [Google Scholar] [CrossRef]
- Isensee, J.; Meoli, L.; Zazzu, V.; Nabzdyk, C.; Witt, H.; Soewarto, D.; Effertz, K.; Fuchs, H.; Gailus-Durner, V.; Busch, D.; et al. Expression pattern of G protein-coupled receptor 30 in LacZ reporter mice. Endocrinology 2009, 150, 1722–1730. [Google Scholar] [CrossRef]
- Wang, C.; Dehghani, B.; Magrisso, I.J.; Rick, E.A.; Bonhomme, E.; Cody, D.B.; Elenich, L.A.; Subramanian, S.; Murphy, S.J.; Kelly, M.J.; et al. GPR30 contributes to estrogen-induced thymic atrophy. Mol. Endocrinol. 2008, 22, 636–648. [Google Scholar] [CrossRef]
- Prossnitz, E.R.; Barton, M. The G-protein-coupled estrogen receptor GPER in health and disease. Nat. Rev. Endocrinol. 2011, 7, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Yang, G.; Hou, Y.; Tang, X.; Wu, C.; Wu, X.A.; Guo, L.; Zhu, Q.; Luo, H.; Du, Y.E.; et al. Cytoplasmic GPER translocation in cancer-associated fibroblasts mediates cAMP/PKA/CREB/glycolytic axis to confer tumor cells with multidrug resistance. Oncogene 2017, 36, 2131–2145. [Google Scholar] [CrossRef] [PubMed]
- Bonuccelli, G.; Whitaker-Menezes, D.; Castello-Cros, R.; Pavlides, S.; Pestell, R.G.; Fatatis, A.; Witkiewicz, A.K.; Vander Heiden, M.G.; Migneco, G.; Chiavarina, B.; et al. The reverse Warburg effect: Glycolysis inhibitors prevent the tumor promoting effects of caveolin-1 deficient cancer associated fibroblasts. Cell Cycle 2010, 9, 1960–1971. [Google Scholar] [CrossRef] [PubMed]
- Whitaker-Menezes, D.; Martinez-Outschoorn, U.E.; Lin, Z.; Ertel, A.; Flomenberg, N.; Witkiewicz, A.K.; Birbe, R.C.; Howell, A.; Pavlides, S.; Gandara, R.; et al. Evidence for a stromal-epithelial “lactate shuttle” in human tumors: MCT4 is a marker of oxidative stress in cancer-associated fibroblasts. Cell Cycle 2011, 10, 1772–1783. [Google Scholar] [CrossRef] [PubMed]
- Lappano, R.; Maggiolini, M. GPER is involved in the functional liaison between breast tumor cells and cancer-associated fibroblasts (CAFs). J. Steroid Biochem. Mol. Biol. 2018, 176, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, Y. In vivo functions of GPR30/GPER-1, a membrane receptor for estrogen: From discovery to functions in vivo. Endocr. J. 2010, 57, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.C.; Harvey, J.M.; Berardo, M.; Clark, G.M. Prognostic and predictive factors in breast cancer by immunohistochemical analysis. Mod. Pathol. 1998, 11, 155–168. [Google Scholar]
- Bilancio, A.; Bontempo, P.; Di Donato, M.; Conte, M.; Giovannelli, P.; Altucci, L.; Migliaccio, A.; Castoria, G. Bisphenol A induces cell cycle arrest in primary and prostate cancer cells through EGFR/ERK/p53 signaling pathway activation. Oncotarget 2017, 8, 115620–115631. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, S.; Wang, Z.; Feng, X.; Liu, P.; Lv, X.B.; Li, F.; Yu, F.X.; Sun, Y.; Yuan, H.; et al. Estrogen regulates Hippo signaling via GPER in breast cancer. J. Clin. Investig. 2015, 125, 2123–2135. [Google Scholar] [CrossRef] [Green Version]
- Pupo, M.; Pisano, A.; Abonante, S.; Maggiolini, M.; Musti, A.M. GPER activates Notch signaling in breast cancer cells and cancer-associated fibroblasts (CAFs). Int. J. Biochem. Cell Biol. 2014, 46, 56–67. [Google Scholar] [CrossRef]
- Yuan, J.; Liu, M.; Yang, L.; Tu, G.; Zhu, Q.; Chen, M.; Cheng, H.; Luo, H.; Fu, W.; Li, Z.; et al. Acquisition of epithelial-mesenchymal transition phenotype in the tamoxifen-resistant breast cancer cell: A new role for G protein-coupled estrogen receptor in mediating tamoxifen resistance through cancer-associated fibroblast-derived fibronectin and β1-integrin signaling pathway in tumor cells. Breast Cancer Res. 2015, 17, 69. [Google Scholar]
- Pandey, D.P.; Lappano, R.; Albanito, L.; Madeo, A.; Maggiolini, M.; Picard, D. Estrogenic GPR30 signalling induces proliferation and migration of breast cancer cells through CTGF. EMBO J. 2009, 28, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; He, H.; Chen, Q.; Yue, W. GPER mediated estradiol reduces miR-148a to promote HLA-G expression in breast cancer. Biochem. Biophys. Res. Commun. 2014, 451, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Vivacqua, A.; De Marco, P.; Santolla, M.F.; Cirillo, F.; Pellegrino, M.; Panno, M.L.; Abonante, S.; Maggiolini, M. Estrogenic GPER signaling regulates miR144 expression in cancer cells and cancer-associated fibroblasts (CAFs). Oncotarget 2015, 6, 16573–16587. [Google Scholar] [CrossRef] [PubMed]
- Meijer, D.; van Agthoven, T.; Bosma, P.T.; Nooter, K.; Dorssers, L.C. Functional screen for genes responsible for tamoxifen resistance in human breast cancer cells. Mol. Cancer Res. 2006, 4, 379–386. [Google Scholar] [CrossRef] [PubMed]
- van Agthoven, T.; Sieuwerts, A.M.; Meijer-van Gelder, M.E.; Look, M.P.; Smid, M.; Veldscholte, J.; Sleijfer, S.; Foekens, J.A.; Dorssers, L.C. Relevance of breast cancer antiestrogen resistance genes in human breast cancer progression and tamoxifen resistance. J. Clin. Oncol. 2009, 27, 542–549. [Google Scholar] [CrossRef] [PubMed]
- De Marco, P.; Bartella, V.; Vivacqua, A.; Lappano, R.; Santolla, M.F.; Morcavallo, A.; Pezzi, V.; Belfiore, A.; Maggiolini, M. Insulin-like growth factor-I regulates GPER expression and function in cancer cells. Oncogene 2013, 32, 678–688. [Google Scholar] [CrossRef]
- Recchia, A.G.; De Francesco, E.M.; Vivacqua, A.; Sisci, D.; Panno, M.L.; Andò, S.; Maggiolini, M. The G protein-coupled receptor 30 is up-regulated by hypoxia-inducible factor-1alpha (HIF-1alpha) in breast cancer cells and cardiomyocytes. J. Biol. Chem. 2011, 286, 10773–10782. [Google Scholar] [CrossRef]
- Manjegowda, M.C.; Gupta, P.S.; Limaye, A.M. Hyper-methylation of the upstream CpG island shore is a likely mechanism of GPER1 silencing in breast cancer cells. Gene 2017, 614, 65–73. [Google Scholar] [CrossRef]
- Giovannelli, P.; Di Donato, M.; Giraldi, T.; Migliaccio, A.; Castoria, G.; Auricchio, F. Targeting rapid action of sex steroid receptors in breast and prostate cancers. Front. Biosci. 2011, 16, 2224–2232. [Google Scholar] [CrossRef]
- Cato, A.C.; Nestl, A.; Mink, S. Rapid actions of steroid receptors in cellular signaling pathways. Sci. STKE 2002, 138, re9. [Google Scholar] [CrossRef]
- Prossnitz, E.R. New developments in the rapid actions of steroids and their receptors. J. Steroid Biochem. Mol. Biol. 2018, 176, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lucas, T.F.; Siu, E.R.; Esteves, C.A.; Monteiro, H.P.; Oliveira, C.A.; Porto, C.S.; Lazari, M.F. 17 beta-estradiol induces the translocation of the estrogen receptors ESR1 and ESR2 to the cell membrane, MAPK3/1 phosphorylation and proliferation of cultured immature rat Sertoli cells. Biol. Reprod. 2008, 78, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.; Filardo, E.J.; Lolait, S.J.; Thomas, P.; Maggiolini, M.; Prossnitz, E.R. Twenty years of the G protein-coupled estrogen receptor GPER: Historical and personal perspectives. J. Steroid Biochem. Mol. Biol. 2018, 176, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
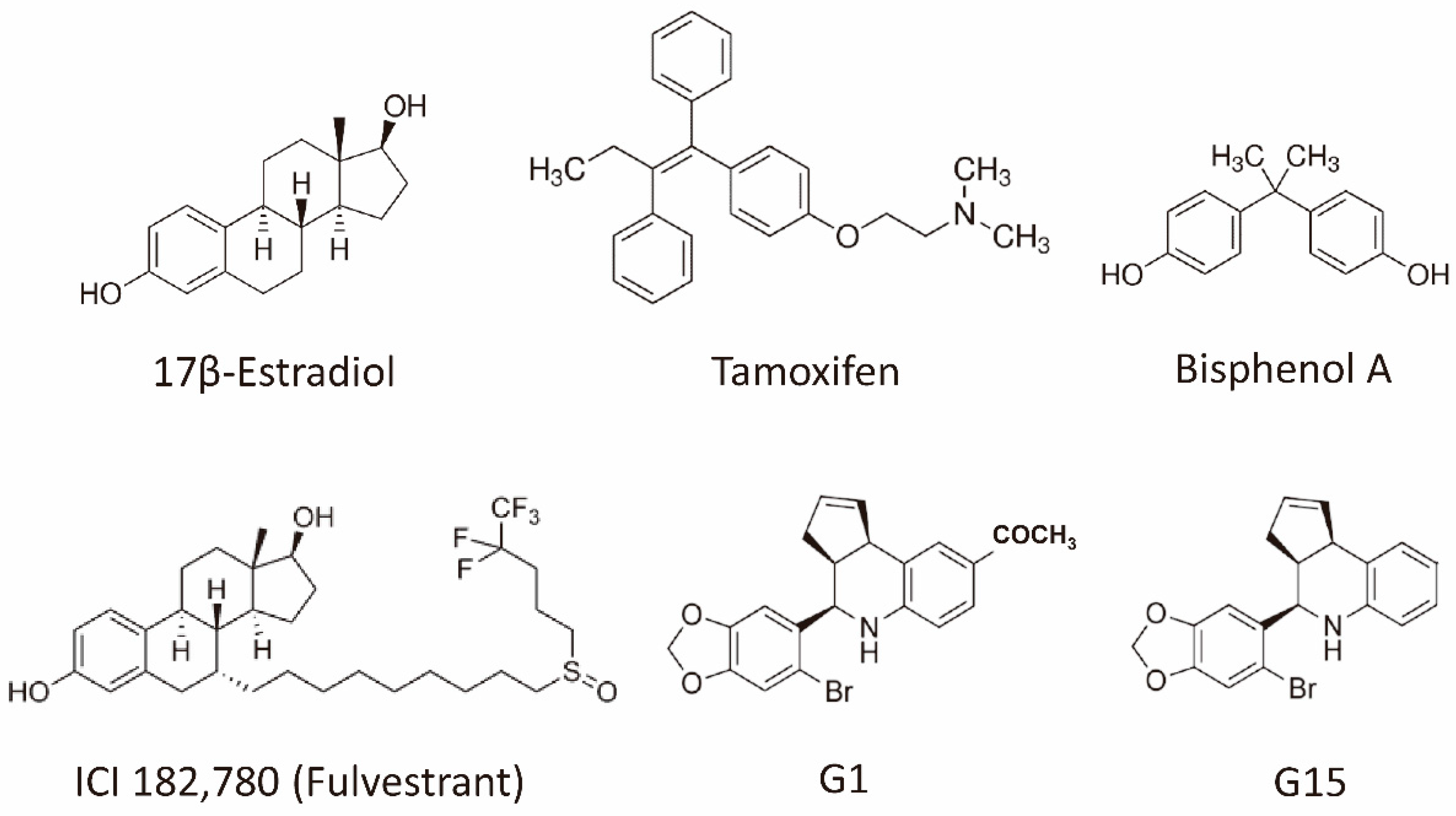
Figure 1.
Structures of the representative GPER agonists and antagonists. 17β-estradiol is one of the major physiological forms of estrogen. Tamoxifen is both a selective estrogen receptor modulator and an agonist for the GPER. Bisphenol A is a xenoestrogen. Fulvestrant is a selective estrogen receptor downregulator (ER antagonist) and an agonist for the GPER. G-1 is a selective GPER agonist, whereas G15 is a selective GPER antagonist. Abbreviation: ER, estrogen receptor; GPER, G-protein coupled estrogen receptor.
Figure 1.
Structures of the representative GPER agonists and antagonists. 17β-estradiol is one of the major physiological forms of estrogen. Tamoxifen is both a selective estrogen receptor modulator and an agonist for the GPER. Bisphenol A is a xenoestrogen. Fulvestrant is a selective estrogen receptor downregulator (ER antagonist) and an agonist for the GPER. G-1 is a selective GPER agonist, whereas G15 is a selective GPER antagonist. Abbreviation: ER, estrogen receptor; GPER, G-protein coupled estrogen receptor.

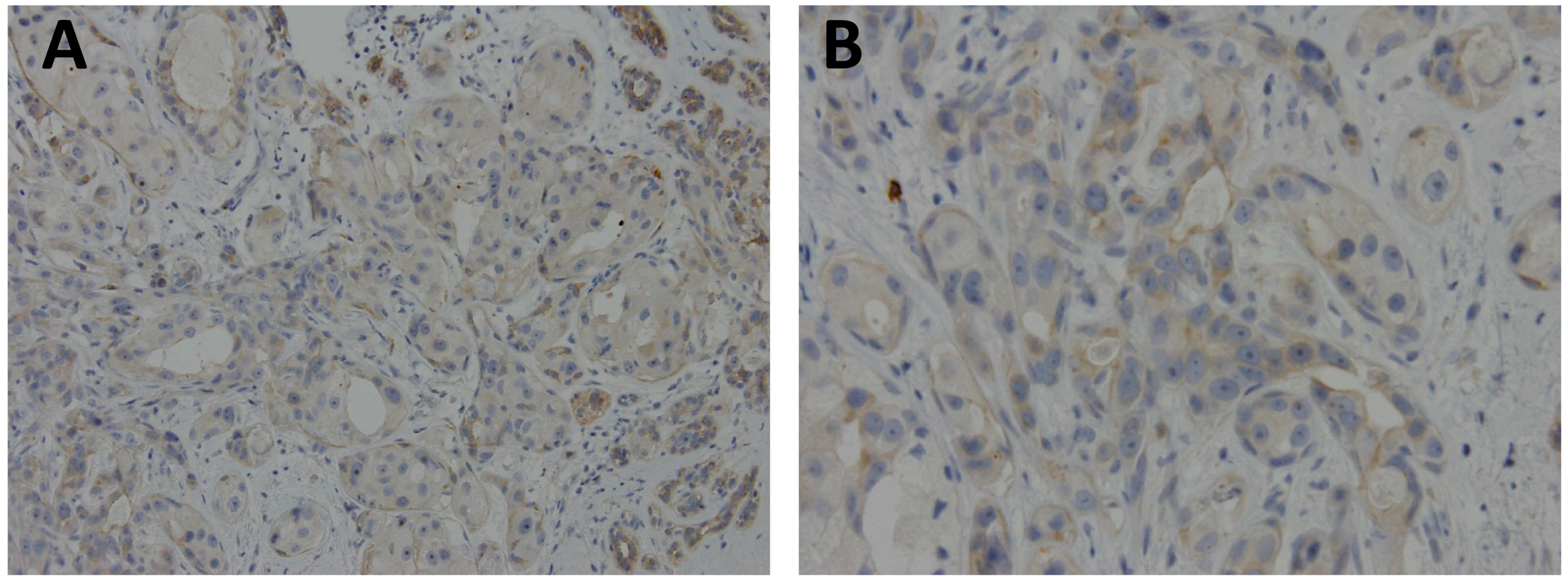
Figure 2.
Representative case of archival, paraffin-embedded breast ductal carcinoma stained with polyclonal GPER1 antibody (Sigma-Aldrich, 1:50 dilution) showed focal, weak membranous and cytoplasmic expression. (A) original × 200; and (B) original × 400.
Figure 2.
Representative case of archival, paraffin-embedded breast ductal carcinoma stained with polyclonal GPER1 antibody (Sigma-Aldrich, 1:50 dilution) showed focal, weak membranous and cytoplasmic expression. (A) original × 200; and (B) original × 400.

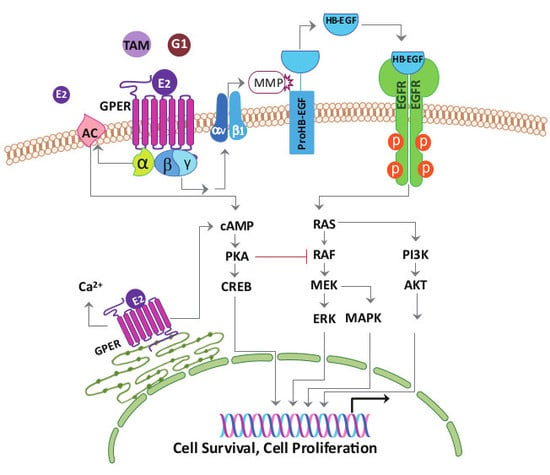
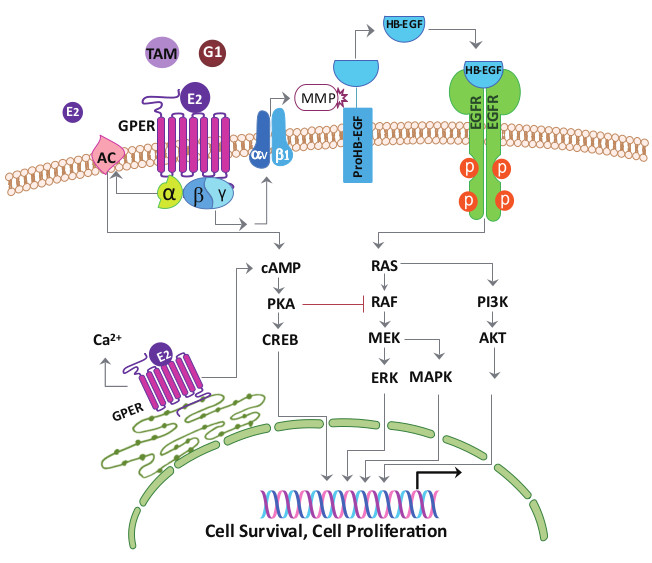
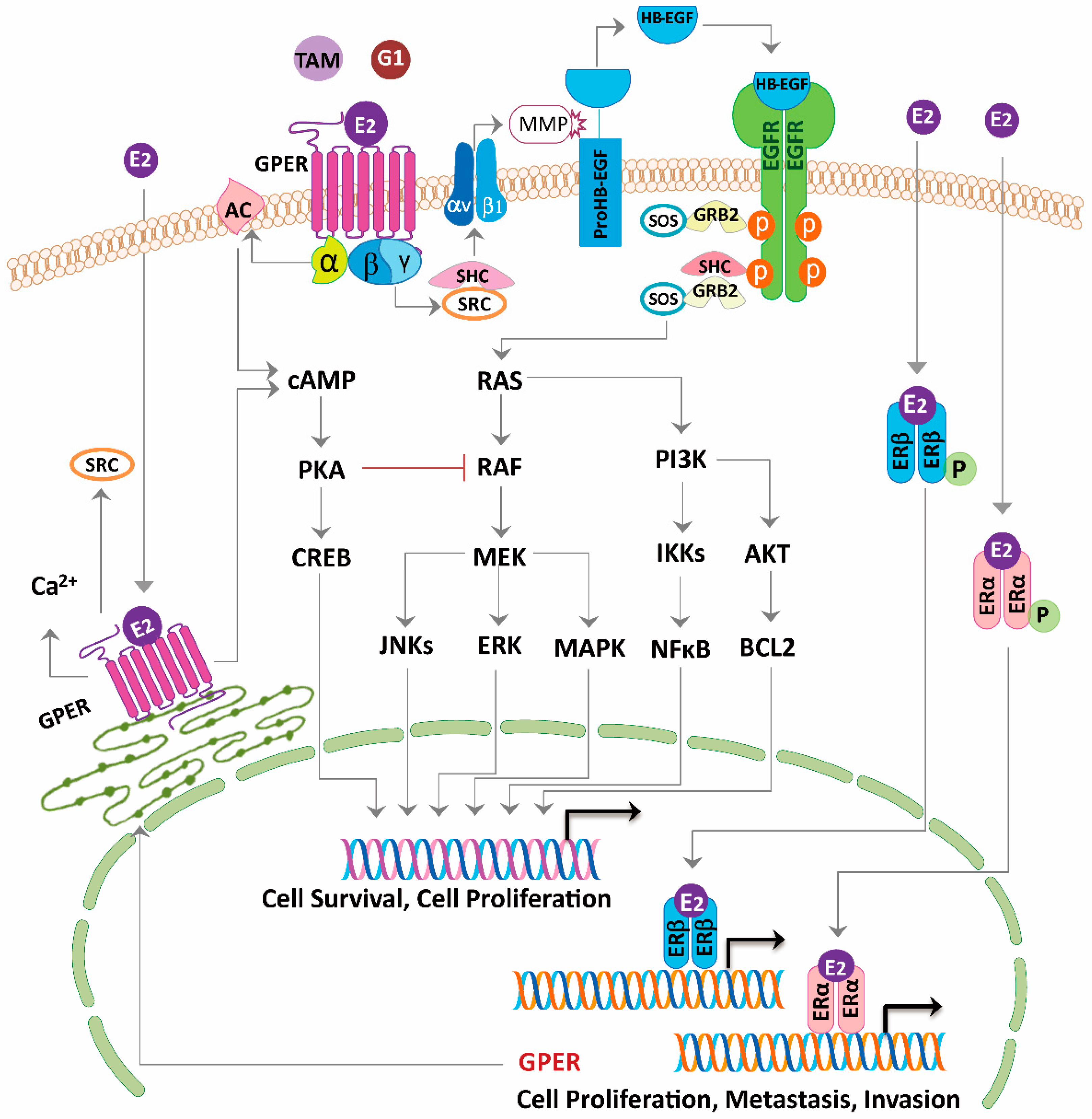
Figure 3.
Model of estrogen signaling pathways in cancer. 17β-estradiol (E2) activates ERα or ERβ to induce the receptor dimerization, and subsequently acts as a transcription factor or interacts with other transcription factors binding to the promoter region of the target genes. E2, tamoxifen (TAM) or G1 activate the G-protein Coupled Estrogen Receptor (GPER) distributed in the nucleus, cytoplasm, and plasma membrane. Activation of GPER located in the plasma membrane stimulates steroid receptor coactivator (SRC) through a Gβγ-subunit protein pathway. The β and γ subunits of the G protein activate the SRC tyrosine kinase, which binds to the integrin ανβ1 through the SHC adapter protein. The complex activates the matrix metalloproteinase (MMP), which then cleaves the pro-heparin-binding EGF-like growth factor (proHB-EGF) and releases the heparin-binding EGF-like growth factor (HB-EGF) into the extracellular space. The free HB-EGF then transactivates the epidermal growth factor receptor (EGFR). Phosphorylation of EGFR in turn activates the downstream pathways, which can induce rapid non-genomic effects, or genomic effects regulating different genes transcription and leads to cell survival and proliferation. On the other hand, through GPER, E2, tamoxifen or G1 is able to stimulate the adenylyl cyclase activity through a Gα-subunit protein pathway, which then leads to the protein kinase A (PKA)-mediated suppression of the EGFR-induced ERK activity. Thus, via the GPER, E2, tamoxifen or G1 may balance the ERK activity by stimulating two distinct G-protein signaling pathways that have opposing effects on the EGFR-to-MAPK axis. Long-term tamoxifen treatment could sensitize the cancer cells through E2-stimulated upregulation of GPER and translocation from the endoplasmic reticulum to the plasma membrane.
Figure 3.
Model of estrogen signaling pathways in cancer. 17β-estradiol (E2) activates ERα or ERβ to induce the receptor dimerization, and subsequently acts as a transcription factor or interacts with other transcription factors binding to the promoter region of the target genes. E2, tamoxifen (TAM) or G1 activate the G-protein Coupled Estrogen Receptor (GPER) distributed in the nucleus, cytoplasm, and plasma membrane. Activation of GPER located in the plasma membrane stimulates steroid receptor coactivator (SRC) through a Gβγ-subunit protein pathway. The β and γ subunits of the G protein activate the SRC tyrosine kinase, which binds to the integrin ανβ1 through the SHC adapter protein. The complex activates the matrix metalloproteinase (MMP), which then cleaves the pro-heparin-binding EGF-like growth factor (proHB-EGF) and releases the heparin-binding EGF-like growth factor (HB-EGF) into the extracellular space. The free HB-EGF then transactivates the epidermal growth factor receptor (EGFR). Phosphorylation of EGFR in turn activates the downstream pathways, which can induce rapid non-genomic effects, or genomic effects regulating different genes transcription and leads to cell survival and proliferation. On the other hand, through GPER, E2, tamoxifen or G1 is able to stimulate the adenylyl cyclase activity through a Gα-subunit protein pathway, which then leads to the protein kinase A (PKA)-mediated suppression of the EGFR-induced ERK activity. Thus, via the GPER, E2, tamoxifen or G1 may balance the ERK activity by stimulating two distinct G-protein signaling pathways that have opposing effects on the EGFR-to-MAPK axis. Long-term tamoxifen treatment could sensitize the cancer cells through E2-stimulated upregulation of GPER and translocation from the endoplasmic reticulum to the plasma membrane.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
G-protein coupled estrogen receptor (GPER) as a prognosticator in breast cancer cell lines and tissues.
Table 1.
G-protein coupled estrogen receptor (GPER) as a prognosticator in breast cancer cell lines and tissues.
| References | Materials | Methods | Subcellular Localizations | Effects on Tumor |
|---|---|---|---|---|
| Tamoxifen-resistant ERα positive cells | ||||
| Ignatov 2010 [26] | MCF-7, TAM-R MCF-7 | Western blot | membrane/endoplasmic reticulum | promoting |
| Ignatov 2011 [27] | TAM-R cancer tissue | immunohistochemistry | nucleus/cytoplasm | promoting |
| Mo 2013 [28] | MCF-7, TAM-R MCF-7 | immunohistochemistry | membrane/cytoplasm | promoting |
| TAM-R cancer tissue | immunofluorescence | |||
| TAM-R mouse xenograft | RT-PCR, Western blot | |||
| Chen 2014 [29] | MCF-7, SkBr3 cells | qRT-PCR, Western blot | non-specified | promoting |
| Catalano 2014 [30] | MCF-7, TAM-R MCF-7, SkBr3, CAF | RT-PCR, Western blot for aromatase activity | non-specified | promoting |
| Triple-negative breast cancer cells | ||||
| Lappano 2010 [35] | MCF-7, SkBr3 | RT-PCR, Western blot | non-specified | promoting |
| Girgert 2012 [36] | MDA-MB-435, HCC1806 | RT-PCR, Western blot | non-specified | promoting |
| Steiman 2013 [37] | TNBC cancer tissue | immunohistochemistry | non-specified | promoting |
| Yu 2014 [38] | MDA-MB-468, MDA-MB-436 | immunohistochemistry | nucleus/cytoplasm | promoting |
| TNBC cancer tissue | immunofluorescence | |||
| RT-PCR, Western blot | ||||
| Zhou 2016 [39] | SkBr3, MDA-MB-231 | Western blot | nucleus/cytoplasm | promoting |
| immunofluorescence | ||||
| Albanito 2008 [40] | SkBr3, BT20 | RT-PCR, Western blot | nucleus/cytoplasm | promoting |
| immunofluorescence | ||||
| Cancer-associated fibroblast | ||||
| Luo 2014, 2016 [66,67] | CAFs isolated from surgical | RT-PCR, Western blot | nucleus/cytoplasm | promoting |
| specimens | immunofluorescence | |||
| Conflicting results | ||||
| Broselid 2013 [55] | ER-(+) cancer tissue | RT-PCR, Western blot | non-specified | suppressive |
| MCF-7 ± GPER knockdown | immunofluorescence | |||
| T47D, HEK ± GPER | ||||
| Poola 2008 [56] | ER-(+)&(−) cancer tissue | qRT-PCR | non-specified | suppressive |
| Kuo 2007 [57] | ER-(+)&(−) cancer tissue | qPCR | non-specified | suppressive |
| Filardo 2002 [58] | MCF-7, SkBr3, MDA-MB-231 | Western blot | non-specified | suppressive |
| Ariazi 2010 [59] | ER-(+)/(−) cancer microarray | RT-PCR, Western blot | non-specified | suppressive |
| MCF-7, SkBr3 | Ca2+ imaging | |||
| Weißenborn 2014 [60] | MCF-7, SkBr3 | RT-PCR, Western blot | non-specified | suppressive |
| methylation PCR | ||||
| bioinformatic | ||||
| Weißenborn 2014 [61] | MDA-MB-231, MDA-MB-468 | RT-PCR, Western blot | non-specified | suppressive |
| methylation PCR | ||||
| bioinformatic | ||||
| Chen 2016 [62] | MDA-MB-231 | qRT-PCR, Western blot | nucleus/cytoplasm | suppressive |
| TNBC cancer tissue | immunofluorescence | |||
| MDA-MB-231 mice xenograft | ||||
| Liang 2017 [63] | MDA-MB-231 | qRT-PCR, Western blot | nucleus/cytoplasm | suppressive |
| TNBC tissue microarray | immunofluorescence | |||
| MDA-MB-231 mice xenograft | ||||
| Okamoto 2016 [64] | SkBr3 cells | qRT-PCR, Western blot | non-specified | suppressive |
ER, estrogen receptor; TAM-R, tamoxifen-resistant; TNBC, triple-negative breast cancer; CAF, cancer-associated fibroblast; qRT-PCR, quantitative RT-PCR.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hsu, L.-H.; Chu, N.-M.; Lin, Y.-F.; Kao, S.-H. G-Protein Coupled Estrogen Receptor in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 306. https://doi.org/10.3390/ijms20020306
AMA Style
Hsu L-H, Chu N-M, Lin Y-F, Kao S-H. G-Protein Coupled Estrogen Receptor in Breast Cancer. International Journal of Molecular Sciences. 2019; 20(2):306. https://doi.org/10.3390/ijms20020306
Chicago/Turabian StyleHsu, Li-Han, Nei-Min Chu, Yung-Feng Lin, and Shu-Huei Kao. 2019. "G-Protein Coupled Estrogen Receptor in Breast Cancer" International Journal of Molecular Sciences 20, no. 2: 306. https://doi.org/10.3390/ijms20020306
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.