Human Leukocyte Antigen and Red Blood Cells Impact Umbilical Cord Blood CD34+ Cell Viability after Thawing

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
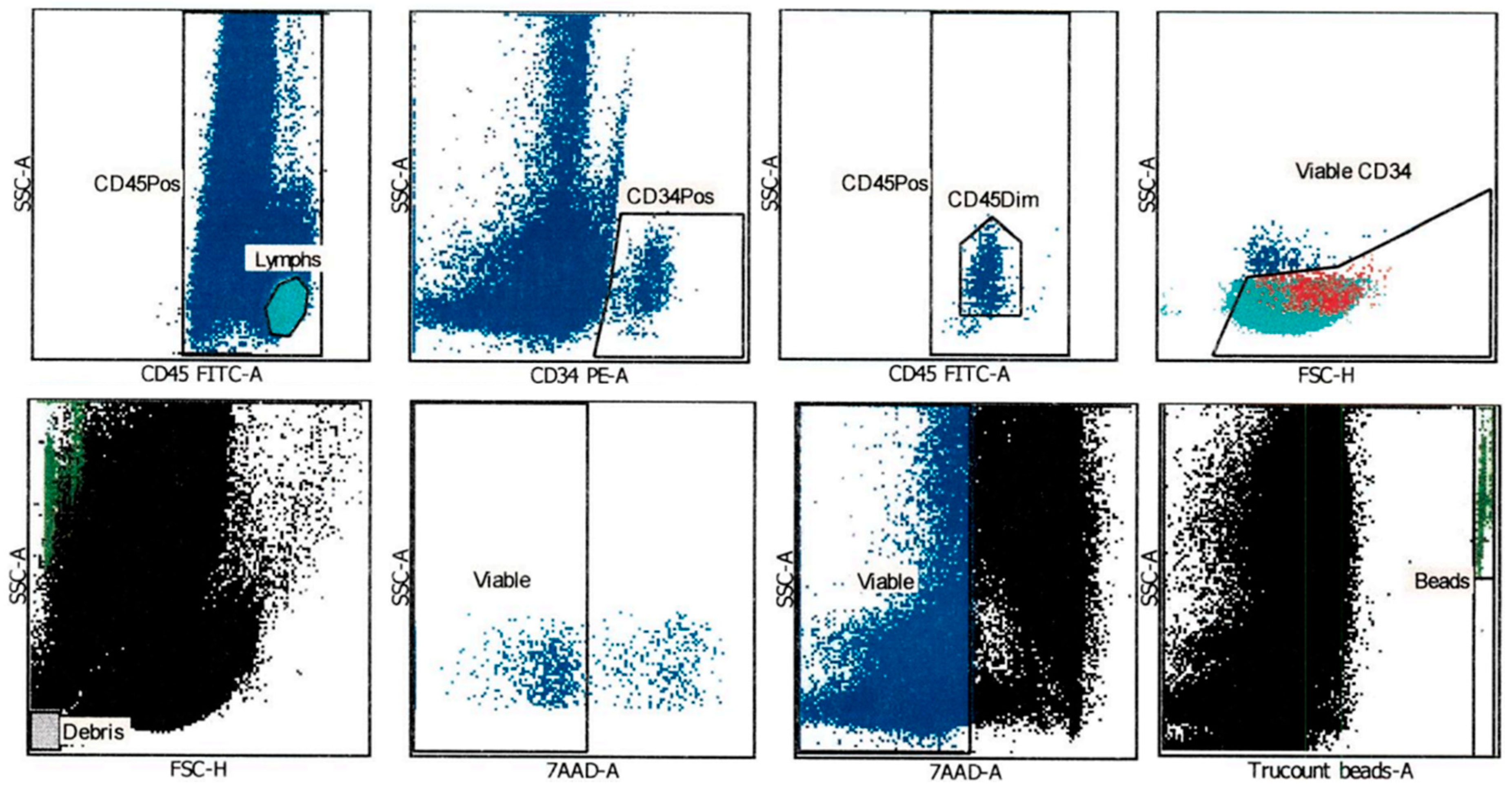
2.1. CD34+ Cell Viability after Thawing was Assessed Using an Automated Gating Method
2.2. Patient-Dependent CD34+ Cell Viability after Thawing
2.3. HLA Genotype per Locus Does Not Impact CD34+ Cell Viability after Thawing
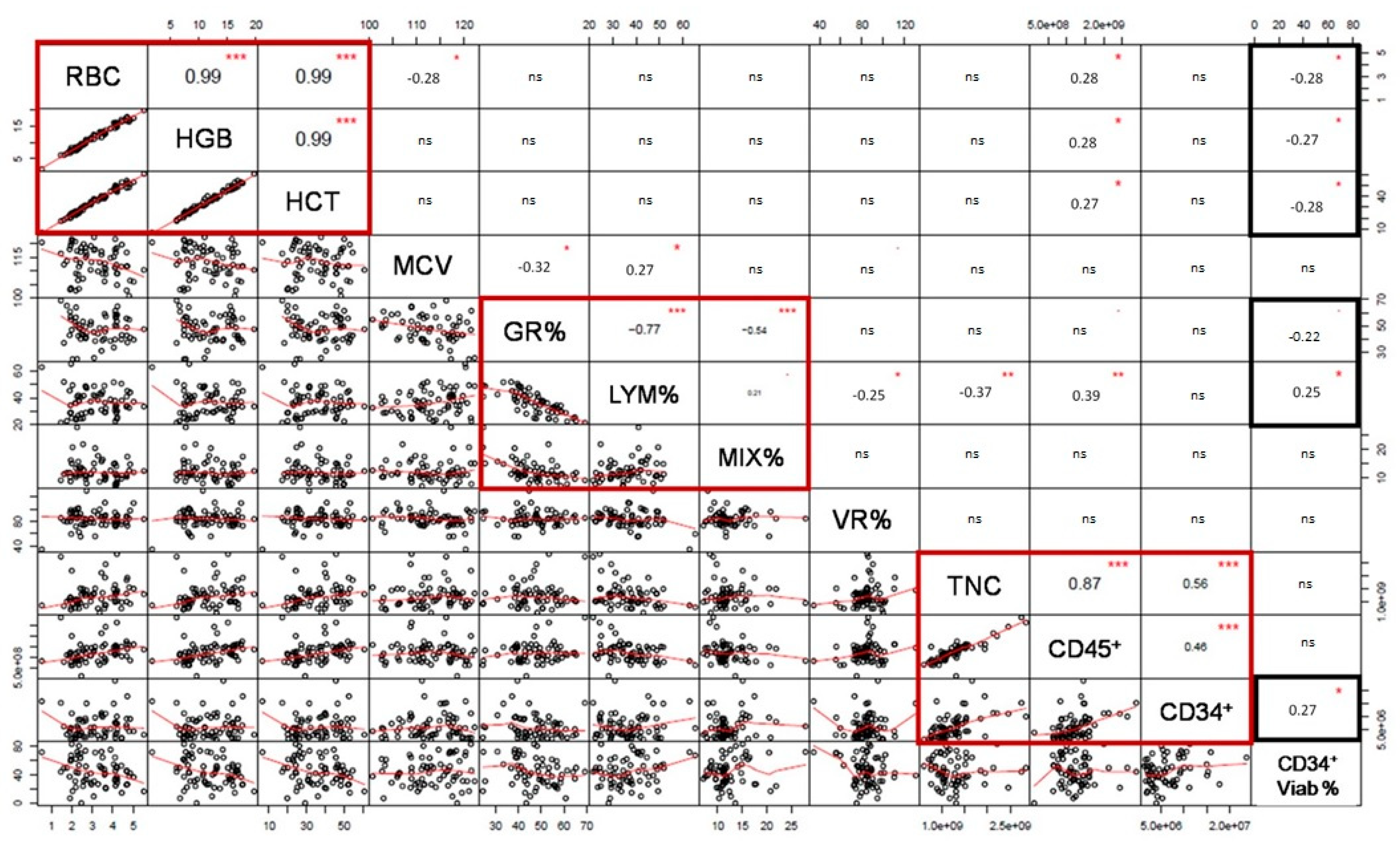
2.4. Pre-Cryopreservation RBC and HLA-B and -C Impact on CD34+ Cell Viability after Thawing
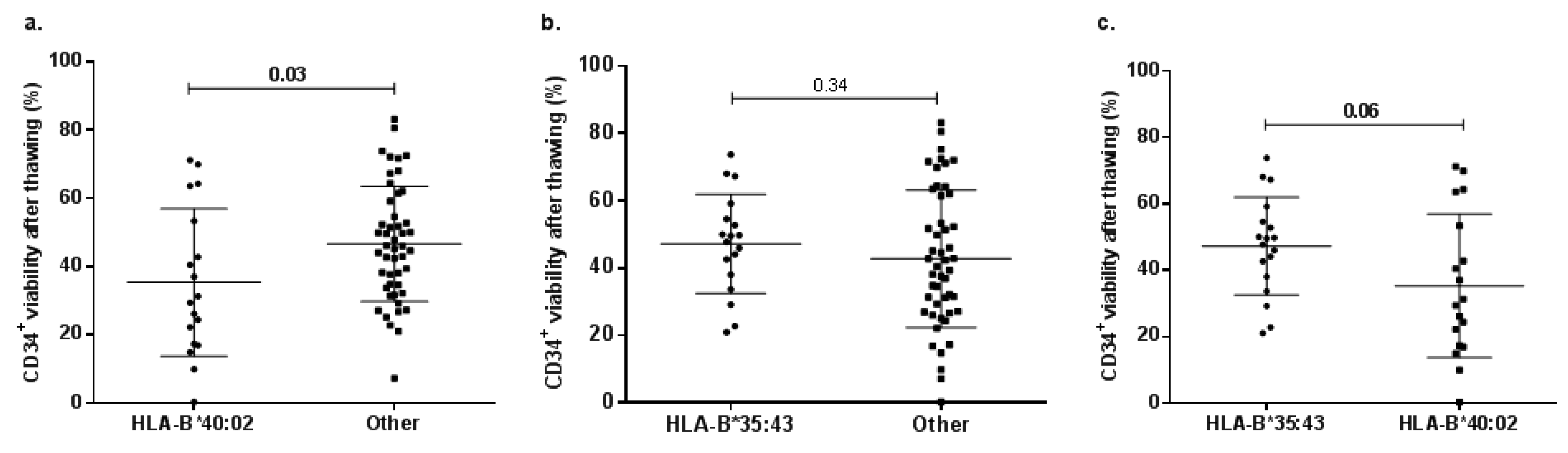
2.5. Allele-Related Differences in CD34+ Cell Viability after Thawing
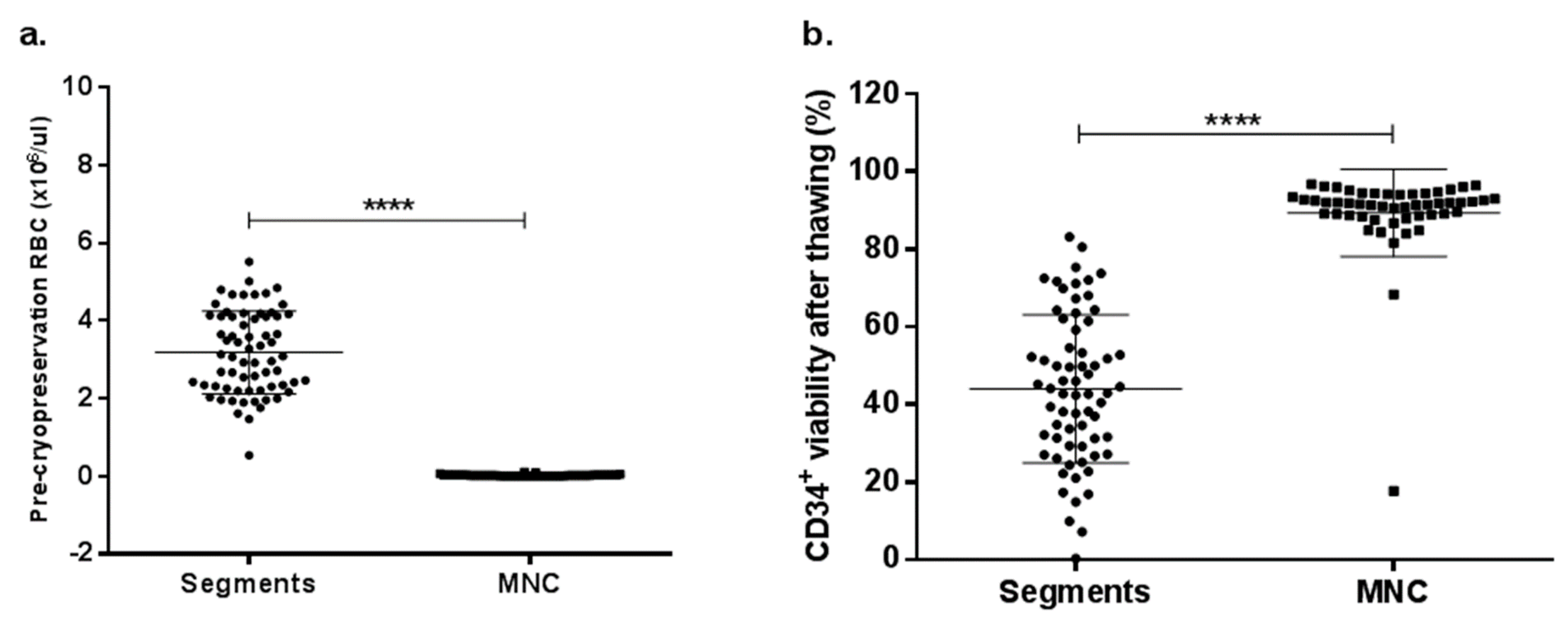
2.6. RBC Depletion Increases CD34+ Cell Viability after Thawing
3. Discussion
4. Materials and Methods
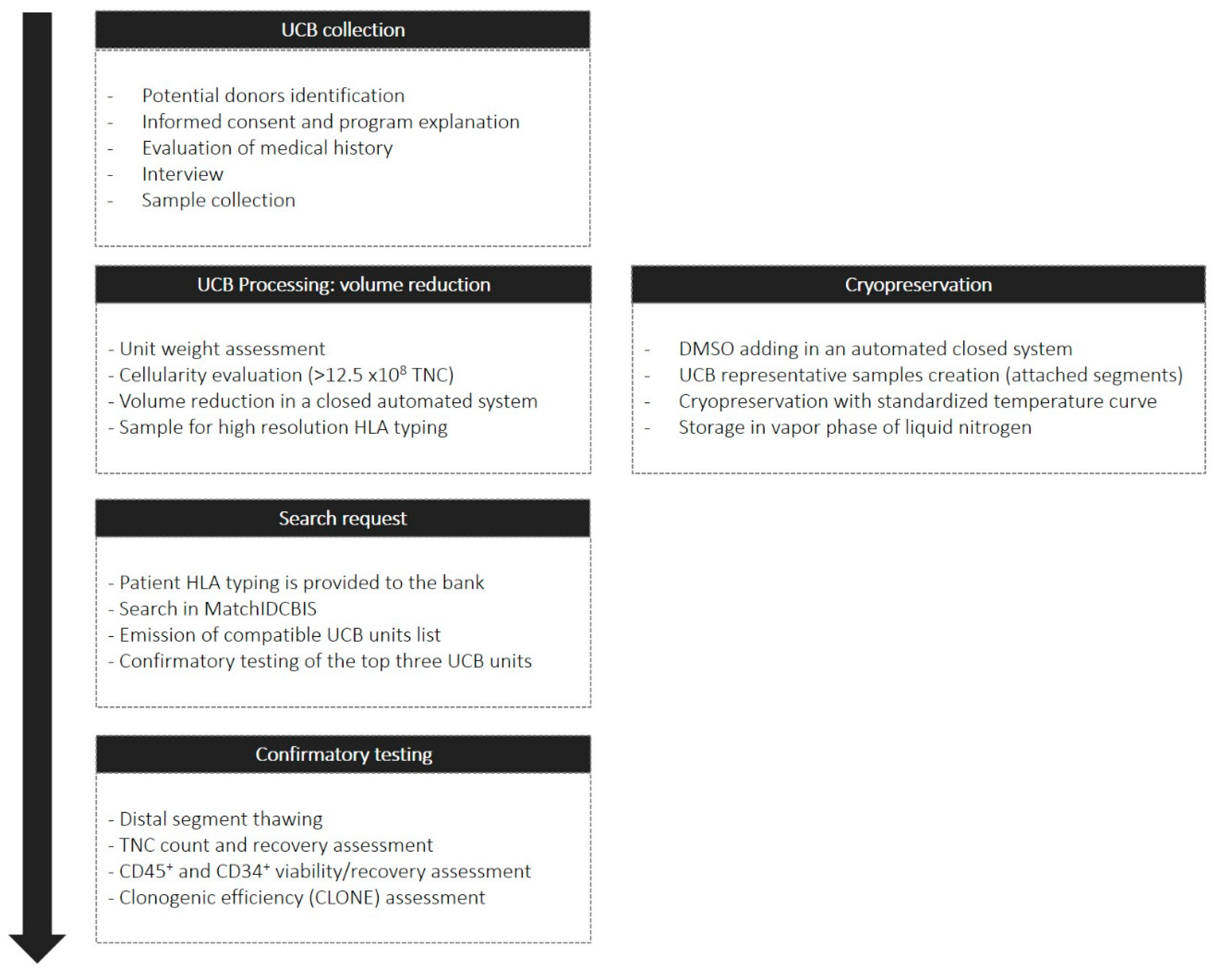
4.1. Umbilical Cord Blood Collection and Volume Reduction
4.2. Cord Blood Cryopreservation and Attached Distal Segment Thawing
4.3. Cord Blood HLA Typing
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HPC | Hematopoietic progenitor cell |
| UCB | Umbilical cord blood |
| HLA | Human leukocyte antigen |
| GVHD | Graft vs. host disease |
| TNC | Total nucleated cell |
| DMSO | Dimethyl sulfoxide |
| RBC | Red blood cells |
| CLONE | Clonogenic efficiency |
| HGB | Hemoglobin |
| HCT | Hematocrit |
| MCV | Mean corpuscular volume |
| GR% | Granulocytes percentage |
| LYM% | Lymphocytes percentage |
| MIX% | Mixed cells percentage |
| VR% | Volume reduction percentage |
Appendix A

References
- Hatzimichael, E.; Tuthill, M. Hematopoietic stem cell transplantation. Stem Cells Cloning Adv. Appl. 2010, 3, 105–117. [Google Scholar] [Green Version]
- Munoz, J.; Shah, N.; Rezvani, K.; Hosing, C.; Bollard, C.M.; Oran, B.; Olson, A.; Popat, U.; Molldrem, J.; Mcniece, I.K.; et al. Concise review: Umbilical cord blood transplantation: Past, present, and future. STEM CELLS Transl. Med. 2014, 3, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Ballen, K.K.; Gluckman, E.; Broxmeyer, H.E. Umbilical cord blood transplantation: The first 25 years and beyond. Blood 2013, 122, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Nakasone, H.; Tabuchi, K.; Uchida, N.; Ohno, Y.; Matsuhashi, Y.; Takahashi, S.; Onishi, Y.; Onizuka, M.; Kobayashi, H.; Fukuda, T.; et al. Which is more important for the selection of cord blood units for haematopoietic cell transplantation: The number of CD 34-positive cells or total nucleated cells? Br. J. Haematol. 2018, 185, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Eapen, M.; Rubinstein, P.; Zhang, M.-J.; Stevens, C.; Kurtzberg, J.; Scaradavou, A.; Loberiza, F.R.; Champlin, R.E.; Klein, J.P.; Horowitz, M.M.; et al. Outcomes of transplantation of unrelated donor umbilical cord blood and bone marrow in children with acute leukaemia: A comparison study. Lancet 2007, 369, 1947–1954. [Google Scholar] [CrossRef]
- Gluckman, E.; Rocha, V.; Ionescu, I.; Bierings, M.; Harris, R.E.; Wagner, J.; Kurtzberg, J.; Champagne, M.A.; Bonfim, C.; Bittencourt, M.; et al. Results of Unrelated Cord Blood Transplant in Fanconi Anemia Patients: Risk Factor Analysis for Engraftment and Survival. Boil. Blood Marrow Transplant. 2007, 13, 1073–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzberg, J.; Wingard, J.R.; Fasth, A.; Nigro, L.L. Umbilical Cord Blood Transplantation for Children with Thalassemia and Sickle Cell Disease. Biol. Blood Marrow Transplant. 2012, 17, 1375–1382. [Google Scholar]
- Barker, J.N.; Byam, C.; Scaradavou, A. How I treat: The selection and acquisition of unrelated cord blood grafts. Blood 2011, 117, 2332–2339. [Google Scholar] [CrossRef]
- Gluckman, E.; Rocha, V.; Arcese, W.; Michel, G.; Sanz, G.; Chan, K.-W.; Takahashi, T.A.; Ortega, J.; Filipovich, A.; Locatelli, F.; et al. Factors associated with outcomes of unrelated cord blood transplant: Guidelines for donor choice. Exp. Hematol. 2004, 32, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Spellman, S.R.; Eapen, M.; Logan, B.R.; Mueller, C.; Rubinstein, P.; Setterholm, M.I.; Woolfrey, A.E.; Horowitz, M.M.; Confer, D.L.; Hurley, C.K.; et al. A perspective on the selection of unrelated donors and cord blood units for transplantation. Blood 2012, 120, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Wagner, J.E.; Barker, J.N.; DeFor, T.E.; Baker, K.S.; Blazar, B.R.; Eide, C.; Goldman, A.; Kersey, J.; Krivit, W.; Macmillan, M.L.; et al. Transplantation of unrelated donor umbilical cord blood in 102 patients with malignant and nonmalignant diseases: Influence of CD34 cell dose and HLA disparity on treatment-related mortality and survival. Blood 2002, 100, 1611–1688. [Google Scholar] [PubMed]
- Sousa, T.; de Sousa, M.; Godinho, M.; Mendes, C.; Carvalhais, A.; Barbosa, I. Umbilical cord blood processing: Volume reduction and recovery of CD34+ cells. Bone Marrow Transplant. 1997, 19, 311–313. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, P.; Dobrila, L.; Rosenfield, R.E.; Adamson, J.W.; Migliaccio, G.; Migliaccio, A.R.; Taylor, P.E.; Stevens, C.E. Processing and cryopreservation of placental/umbilical cord blood for unrelated bone marrow reconstitution. Proc. Natl. Acad. Sci. USA 1995, 92, 10119–10122. [Google Scholar] [CrossRef] [PubMed]
- Hunt, C.J.; Armitage, S.E.; Pegg, D.E. Cryopreservation of umbilical cord blood: 2. Tolerance of CD34+ cells to multimolar dimethyl sulphoxide and the effect of cooling rate on recovery after freezing and thawing. Cryobiology 2003, 46, 76–87. [Google Scholar] [CrossRef]
- Woods, E.J.; Liu, J.; Derrow, C.W.; Smith, F.O.; Williams, D.A.; Critser, J.K. Cutting Edge Communication: Osmometric and Permeability Characteristics of Human Placental/Umbilical Cord Blood CD34T+ Cells and Their Application to Cryopreservation. J. Hematother. Stem Cell Res. 2000, 9, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Fry, L.J.; Giner, S.Q.; Gomez, S.G.; Green, M.; Anderson, S.; Horder, J.; McArdle, S.; Rees, R.; Madrigal, J.A. Avoiding room temperature storage and delayed cryopreservation provide better postthaw potency in hematopoietic progenitor cell grafts. Transfusion 2013, 53, 1834–1842. [Google Scholar] [CrossRef]
- Ruggeri, A.; Paviglianiti, A.; Gluckman, E.; Rocha, V. Impact of HLA in cord blood transplantation outcomes. HLA 2016, 87, 413–421. [Google Scholar] [CrossRef]
- Eapen, M.; Klein, J.P.; Sanz, G.F.; Spellman, S.; Ruggeri, A.; Anasetti, C.; Brown, M.; Champlin, R.E.; Garcia-Lopez, J.; Hattersely, G.; et al. Impact of donor-recipient HLA-matching at HLA-A,-B,-C and-DRB1 on outcomes after umbilical cord blood transplantation for leukemia and myelodysplastic syndrome: A retrospective analysis. Lancet Oncol. 2011, 12, 1214–1221. [Google Scholar] [CrossRef]
- Garfall, A.L.; Kim, H.T.; Cutler, C.; Ho, V.T.; Koreth, J.; Armand, P.; Chen, Y.B.A.; McAfee, S.L.; Dey, B.R.; Liney, D.; et al. Allele Matching At HLA-C or DRB1 Is Associated with Improved Survival After Reduced Intensity Double Umbilical Cord Blood Transplantation. Blood 2012, 120, 21. [Google Scholar]
- Brunstein, C.G.; Cutler, C.S.; DeFor, T.E.; Spitzer, T.R.; Bejanyan, N.; Garfall, A.L.; Verneris, M.R.; Chen, Y.B.; Warlick, E.; Kim, H.T.; et al. Antigen Level Matching at HLA-C Improves Long-Term Outcomes after Double Umbilical Cord Blood Transplantation. Blood 2015, 126, 23. [Google Scholar]
- Rocha, V.; Gluckman, E.; on behalf of the Eurocord-Netcord registry and European Blood and Marrow Transplant group. Improving outcomes of cord blood transplantation: HLA matching, cell dose and other graft- and transplantation-related factors. Br. J. Haematol. 2009, 147, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Pidala, J.; Lee, S.J.; Ahn, K.W.; Spellman, S.; Wang, H.L.; Aljurf, M.; Askar, M.; Dehn, J.; Viña, M.F.; Gratwohl, A.; et al. Nonpermissive HLA-DPB1 mismatch increases mortality after myeloablative unrelated allogeneic hematopoietic cell transplantation Key Points. Blood 2014, 124, 2596–2606. [Google Scholar] [CrossRef] [PubMed]
- Eapen, M.; Klein, J.P.; Ruggeri, A.; Spellman, S.; Lee, S.J.; Anasetti, C.; Arcese, W.; Barker, J.N.; Baxter-Lowe, L.A.; Brown, M.; et al. Impact of allele-level HLA matching on outcomes after myeloablative single unit umbilical cord blood transplantation for hematologic malignancy. Blood 2014, 123, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Klein, J.; Haagenson, M.; Baxter-Lowe, L.A.; Confer, D.L.; Eapen, M.; Fernandez-Vina, M.; Flomenberg, N.; Horowitz, M.; Hurley, C.K.; et al. High-resolution donor-recipient HLA matching contributes to the success of unrelated donor marrow transplantation. Blood 2007, 110, 4576–4583. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, D.R.; Anderson, L.; Keeney, M.; Nayar, R.; Chin-Yee, I. The ISHAGE Guidelines for CD34+ Cell Determination by Flow Cytometry. J. Hematotherapy 1996, 5, 213–226. [Google Scholar] [CrossRef]
- Brocklebank, A.M.; Sparrow, R.L. Enumeration of CD34+ cells in cord blood: A variation on a single-platform flow cytometric method based on the ISHAGE gating strategy. Cytometry 2001, 44, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Galindo, C.C.; Lozano, D.M.V.; Rodríguez, B.C.; Perdomo-Arciniegas, A.-M. Improved cord blood thawing procedure enhances the reproducibility and correlation between flow cytometry CD34+ cell viability and clonogenicity assays. Cytotherapy 2018, 20, 891–894. [Google Scholar] [CrossRef]
- Galindo, C.C.; Lozano, D.M.V.; Rodríguez, B.C. Cord blood attached-segments are not homogeneous in post-thaw CD34+ cell viability and clonogenicity. Cryobiology 2018, 82, 163–164. [Google Scholar] [CrossRef]
- Pope, B.; Hokin, B.; Grant, R. Effect of umbilical cord blood prefreeze variables on postthaw viability. Transfusion 2015, 55, 629–635. [Google Scholar] [CrossRef]
- Páez-Gutiérrez, I.A.; Hernández-Mejía, D.G.; Vanegas, D.; Camacho-Rodríguez, B.; Perdomo-Arciniegas, A.M. HLA-A, -B, -C, -DRB1 and -DQB1 allele and haplotype frequencies of 1463 umbilical cord blood units typed in high resolution from Bogotá, Colombia. Hum. Immunol. 2019, 80, 425–426. [Google Scholar] [CrossRef]
- Eapen, M.; Wang, T.; Veys, P.A.; Boelens, J.J.; St Martin, A.; Spellman, S.; Bonfim, C.S.; Brady, C.; Cant, A.J.; Dalle, J.H.; et al. Allele-level HLA matching for umbilical cord blood transplantation for non-malignant diseases in children: A retrospective analysis. Lancet Haematol. 2017, 4, e325–e333. [Google Scholar] [CrossRef]
- Broxmeyer, H.E.; Douglas, G.W.; Hangoc, G.; Cooper, S.; Bard, J.; English, D.; Arny, M.; Thomas, L.; Boyse, E.A. Human umbilical cord blood as a potential source of transplantable hematopoietic stem/progenitor cells. Proc. Natl. Acad. Sci. USA 1989, 86, 3828–3832. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, V.; Pellegrini, N.; Bardey, I.; Malugani, C.; Saas, P.; Garnache, F.; Racadot, E.; Maddens, S.; Schillinger, F. Cord blood volume reduction using an automated system (Sepax) vs. a semi-automated system (Optipress II) and a manual method (hydroxyethyl starch sedimentation) for routine cord blood banking: A comparative study. Cytotherapy 2007, 9, 165–169. [Google Scholar] [CrossRef]
- Assmus, B.; Tonn, T.; Seeger, F.H.; Yoon, C.-H.; Leistner, D.; Klotsche, J.; Schächinger, V.; Seifried, E.; Zeiher, A.M.; Dimmeler, S. Red Blood Cell Contamination of the Final Cell Product Impairs the Efficacy of Autologous Bone Marrow Mononuclear Cell Therapy. J. Am. Coll. Cardiol. 2010, 55, 1385–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaguma, Y.; Akatsuka, Y.; Hosokawa, K.; Maruyama, H.; Okamoto, A.; Katagiri, T.; Shiraishi, K.; Murayama, Y.; Tsuzuki-Iba, S.; Mizutani, Y.; et al. Induction of HLA-B*40:02-restricted T cells possessing cytotoxic and suppressive functions against haematopoietic progenitor cells from a patient with severe aplastic anaemia. Br. J. Haematol. 2016, 172, 131–134. [Google Scholar] [CrossRef]
- Chen, C.; Lu, S.; Luo, M.; Zhang, B.; Xiao, L. Correlations between HLA-A, HLA-B and HLA-DRB1 Allele Polymorphisms and Childhood Susceptibility to Acquired Aplastic Anemia. Acta Haematol. 2012, 128, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Jeong, T.D.; Mun, Y.C.; Chung, H.S.; Seo, D.; Im, J.; Huh, J. Novel deletion mutation of HLA-B*40:02 gene in acquired aplastic anemia. HLA 2017, 89, 47–51. [Google Scholar]
- Stanley, N.; Olson, T.S.; Babushok, D.V. Recent advances in understanding clonal haematopoiesis in aplastic anaemia. Br. J. Haematol. 2017, 177, 509–525. [Google Scholar] [CrossRef] [Green Version]
- Zaimoku, Y.; Takamatsu, H.; Hosomichi, K.; Ozawa, T.; Nakagawa, N.; Imi, T.; Maruyama, H.; Katagiri, T.; Kishi, H.; Tajima, A.; et al. Identification of an HLA class I allele closely involved in the autoantigen presentation in acquired aplastic anemia Key Points. Blood 2017, 129, 2908–2916. [Google Scholar] [CrossRef]
- Akel, S.; Regan, D.; Wall, D.; Petz, L.; McCullough, J. Current thawing and infusion practice of cryopreserved cord blood: The impact on graft quality, recipient safety, and transplantation outcomes. Transfusion 2014, 54, 2997–3009. [Google Scholar] [CrossRef]
- Vanegas, D.; Triviño, L.; Galindo, C.; Franco, L.; Salguero, G.; Camacho, B.; Perdomo-Arciniegas, A. A new strategy for umbilical cord blood collection developed at the first Colombian public cord blood bank increases total nucleated cell content. Transfusion 2017, 57, 2225–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cereb, N.; Kim, H.R.; Ryu, J.; Yang, S.Y. Advances in DNA sequencing technologies for high resolution HLA typing. Hum. Immunol. 2015, 76, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, C.; Schmidt, A.H.; Sauter, J. Hapl-o-Mat: Open-source software for HLA haplotype frequency estimation from ambiguous and heterogeneous data. BMC Bioinform. 2017, 18, 284. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 30 September 2019).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Variables after Thawing | |||||||
|---|---|---|---|---|---|---|---|
| Variable | n | Mean | Median | SD | SEM | Minimum | Maximum |
| TNC (×108) | 65 | 10.57 | 9.92 | 4.51 | 0.56 | 3.10 | 23.30 |
| TNC Recovery (%) | 65 | 85.10 | 88.10 | 26.33 | 3.27 | 23.70 | 141.50 |
| Viable CD45+ (×108) | 67 | 3.97 | 3.69 | 1.65 | 0.20 | 0.12 | 7.99 |
| CD45+ Recovery (%) | 67 | 33.82 | 34.50 | 14.34 | 1.75 | 8.90 | 71.90 |
| CD45+ Viability (%) | 60 | 49.98 | 49.35 | 13.38 | 1.73 | 5.10 | 81.0 |
| Viable CD34+ (×106) | 67 | 2.64 | 1.67 | 2.43 | 0.30 | 0.02 | 13.20 |
| CD34+ Recovery (%) | 67 | 40.57 | 40.20 | 20.37 | 2.49 | 0.60 | 83.30 |
| CD34+ Viability (%) | 67 | 44.02 | 42.90 | 19.06 | 2.33 | 0.31 | 83.20 |
| CLONE (%) | 64 | 36.40 | 36.35 | 18.10 | 2.26 | 0.0 | 82.30 |
| Multivariate Model (Dependent Variable: CD34+ Cell Viability) | ||||
|---|---|---|---|---|
| HLA | Explanatory Variables | R2 | p | Multicollinearity |
| A | RBC, GR%, CD34+ | 0.05 | 0.449 | No |
| RBC, GR%, TNC | 0.05 | 0.444 | No | |
| B | RBC, GR%, CD34+ | 0.95 | 0.014 | RBC, GR%, CD34+ |
| RBC, GR%, TNC | 0.79 | 0.108 | RBC, GR% | |
| C | RBC, GR%, CD34+ | 0.56 | 0.007 | No |
| RBC, GR%, TNC | 0.55 | 0.007 | No | |
| DRB1 | RBC, GR%, CD34+ | 0.26 | 0.316 | %GR, CD34+ |
| RBC, GR%, TNC | 0.28 | 0.298 | CNT | |
| DQB1 | RBC, GR%, CD34+ | 0.27 | 0.060 | No |
| RBC, GR%, TNC | 0.26 | 0.066 | No | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanegas, D.; Galindo, C.-C.; Páez-Gutiérrez, I.-A.; González-Acero, L.-X.; Medina-Valderrama, P.-T.; Lozano, J.-C.; Camacho-Rodríguez, B.; Perdomo-Arciniegas, A.-M. Human Leukocyte Antigen and Red Blood Cells Impact Umbilical Cord Blood CD34+ Cell Viability after Thawing. Int. J. Mol. Sci. 2019, 20, 4875. https://doi.org/10.3390/ijms20194875
Vanegas D, Galindo C-C, Páez-Gutiérrez I-A, González-Acero L-X, Medina-Valderrama P-T, Lozano J-C, Camacho-Rodríguez B, Perdomo-Arciniegas A-M. Human Leukocyte Antigen and Red Blood Cells Impact Umbilical Cord Blood CD34+ Cell Viability after Thawing. International Journal of Molecular Sciences. 2019; 20(19):4875. https://doi.org/10.3390/ijms20194875
Chicago/Turabian StyleVanegas, Diana, Cristian-Camilo Galindo, Iván-Aurelio Páez-Gutiérrez, Lorena-Xiomara González-Acero, Pavel-Tiberio Medina-Valderrama, Juan-Camilo Lozano, Bernardo Camacho-Rodríguez, and Ana-María Perdomo-Arciniegas. 2019. "Human Leukocyte Antigen and Red Blood Cells Impact Umbilical Cord Blood CD34+ Cell Viability after Thawing" International Journal of Molecular Sciences 20, no. 19: 4875. https://doi.org/10.3390/ijms20194875
APA StyleVanegas, D., Galindo, C.-C., Páez-Gutiérrez, I.-A., González-Acero, L.-X., Medina-Valderrama, P.-T., Lozano, J.-C., Camacho-Rodríguez, B., & Perdomo-Arciniegas, A.-M. (2019). Human Leukocyte Antigen and Red Blood Cells Impact Umbilical Cord Blood CD34+ Cell Viability after Thawing. International Journal of Molecular Sciences, 20(19), 4875. https://doi.org/10.3390/ijms20194875