Role of Intracellular Na+ in the Regulation of [Ca2+]i in the Rat Suprachiasmatic Nucleus Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
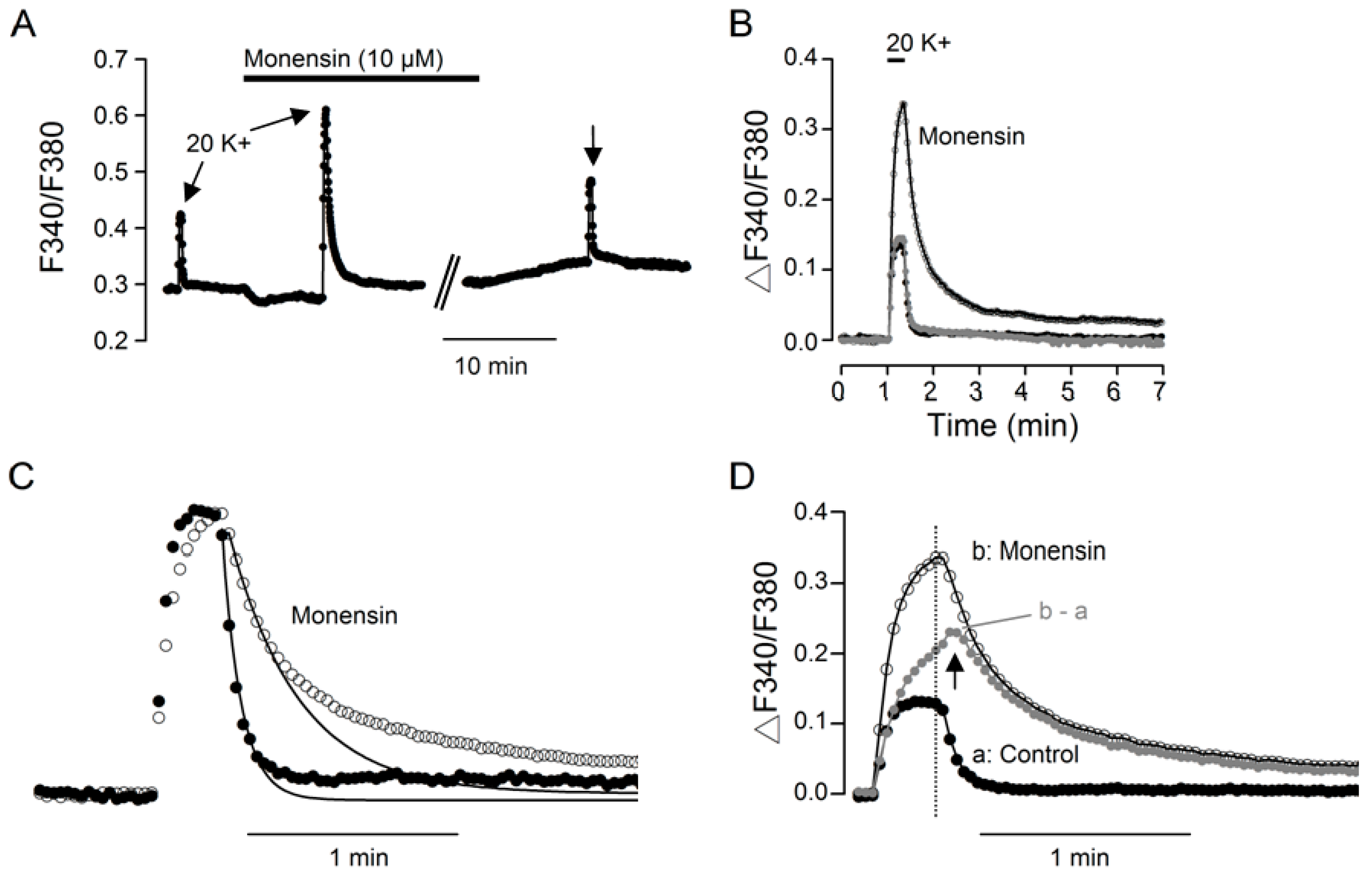
2.1. Monensin Effects
2.2. Ouabain Effects
2.3. Ouabain Preferentially Enhances Nimodipine-Insensitive Ca2+ Rise
3. Discussion
3.1. Monensin Effects
3.2. Ouabain Effects
3.3. Functional Implicationss
4. Materials and Methods
4.1. Hypothalamic Brain Slices and Reduced SCN Preparations
4.2. Electrical Recordings
4.3. Ca2+ and Na+ Imaging
4.4. Drugs
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FCCP | Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone |
| NCX | Na+/Ca2+ exchanger |
| NKA | Na+/K+-ATPase |
| SCN | Suprachiasmatic nucleus |
References
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [PubMed]
- Green, D.J.; Gillette, R. Circadian rhythm of firing rate recorded from single cells in the rat suprachiasmatic brain slice. Brain Res. 1982, 245, 198–200. [Google Scholar] [CrossRef]
- Groos, G.A.; Hendriks, J. Circadian rhythms in electrical discharge of rat suprachiasmatic neurones recorded in vitro. Neurosci. Lett. 1982, 34, 283–288. [Google Scholar] [CrossRef]
- Inouye, S.I.T.; Kawamura, H. Persistence of circadian rhythmicity in a mammalian hypothalamic ‘island’ containing the suprachiasmatic nucleus. Proc. Natl. Acad. Sci. USA 1979, 76, 5962–5966. [Google Scholar] [CrossRef] [PubMed]
- Shibata, S.; Oomura, Y.; Kita, H.; Hattori, K. Circadian rhythmic changes of neuronal activity in the suprachiasmatic nucleus of the rat hypothalamic slice. Brain Res. 1982, 247, 154–158. [Google Scholar] [CrossRef]
- Colwell, C.S. Circadian modulation of calcium levels in cells in the suprachiasmatic nucleus. Eur. J. Neurosci. 2000, 12, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Sugiyama, T.; Wallace, C.S.; Gompf, H.S.; Yoshioka, T.; Miyawaki, A.; Allen, C.N. Circadian dynamics of cytosolic and nuclear Ca2+ in single suprachiasmatic nucleus neurons. Neuron 2003, 38, 253–263. [Google Scholar] [CrossRef]
- Enoki, R.; Huroda, S.; Ono, D.; Hasan, M.T.; Ueda, T.; Honma, S.; Honma, K.I. Topological specificity and hierarchical network of the circadian calcium rhythm in the suprachiasmatic nucleus. Proc. Nat. Acad. Sci. USA 2012, 109, 21498–21503. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Huang, R.C. Diurnal modulation of the Na+/K+-ATPase and spontaneous firing in the rat retinorecipient clock neurons. J. Neurophysiol. 2004, 92, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chen, Y.S.; Cheng, R.C.; Huang, R.C. Role of Na+/Ca2+ exchanger in Ca2+ homeostasis in the rat suprachiasmatic nucleus neurons. J. Neurophysiol. 2015, 113, 2114–2126. [Google Scholar] [CrossRef][Green Version]
- López, L.; Lorente, L.; Arias, J.; González-Pardo, H.; Cimadevilla, J.; Arias, J.L. Changes of cytochrome oxidase activity in rat suprachiasmatic nucleus. Brain Res. 1997, 769, 367–371. [Google Scholar] [CrossRef]
- Newman, G.C.; Hospod, F.E.; Patlak, C.S.; Moore, R.Y. Analysis of in vitro glucose utilization in a circadian pacemaker model. J. Neurosci. 1992, 12, 2015–2021. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, W.J.; Gainer, H. Suprachiasmatic nucleus: use of 14C-labeled deoxyglucose uptake as a functional marker. Science 1977, 197, 1089–1091. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.C.; Wang, Y.C.; Chen, Y.S.; Cheng, R.C.; Yang, J.J.; Huang, R.C. Differential regulation of nimodipine-sensitive and -insensitive Ca2+ influx by the Na+/Ca2+ exchanger and mitochondria in the rat suprachiasmatic nucleus neurons. J. Biomed. Sci. 2018, 25, 44. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, M.P.; Lederer, W.J. Sodium/calcium exchange: its physiological implications. Physiol. Rev. 1999, 79, 763–854. [Google Scholar] [CrossRef]
- De Stefani, D.; Rizzuto, R.; Pozzan, T. Enjoy the trip: Calcium in mitochondria back and forth. Ann. Rev. Biochem. 2016, 85, 161–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Yang, J.J.; Huang, R.C. Intracellular Na+ and metabolic modulation of Na/K pump and excitability in the rat suprachiasmatic nucleus neurons. J. Neurophysiol. 2012, 108, 2024–2032. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cheng, R.C.; Cheng, P.C.; Huang, R.C. Department of Physiology and Pharmacology, College of Medicine, Chang Gung University, Tao-Yuan, Taiwan. Unpublished observation. 2019. [Google Scholar]
- Wang, Y.C.; Huang, R.C. Effects of sodium pump activity on spontaneous firing in neurons of the rat suprachiasmatic nucleus. J. Neurophysiol. 2006, 96, 109–118. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thayer, S.A.; Miller, R.J. Regulation of the intracellular free calcium concentration in single rat dorsal root ganglion neurones in vitro. J. Physiol. 1990, 425, 85–115. [Google Scholar] [CrossRef]
- Friel, D.D.; Tsien, R.W. An FCCP-sensitive Ca2+ store in bullfrog sympathetic neurons and its participation in stimulus-evoked changes in [Ca2+]i. J. Neurosci. 1994, 14, 4007–4024. [Google Scholar] [CrossRef] [PubMed]
- Baron, K.T.; Thayer, S.A. CGP37157 modulates mitochondrial Ca2+ homeostasis in cultured rat dorsal root ganglion neurons. Eur. J. Pharmacol. 1997, 340, 295–300. [Google Scholar] [CrossRef]
- Colegrove, S.L.; Albrecht, M.A.; Friel, D.D. Dissection of mitochondrial Ca2+ uptake and release fluxes in situ after depolarization-evoked [Ca2+]i elevations in sympathetic neurons. J. Gen. Physiol. 2000, 115, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Sedova, M.; Blatter, L.A. Intracellular sodium modulates mitochondrial calcium signaling in vascular endothelial cells. J. Biol. Chem. 2000, 275, 35402–35407. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Matsuoka, S. Cytoplasmic Na+-dependent modulation of mitochondrial Ca2+ via electrogenic mitochondrial Na+-Ca2+ exchange. J. Physiol. 2008, 586, 1683–1697. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.C.; Lin, H.Y.; Chen, Y.S.; Cheng, R.C.; Su, H.C.; Huang, R.C. The Na+/H+-exchanger NHE1 regulates extra- and intracellular pH and nimodipine-sensitive [Ca2+]i in the suprachiasmatic nucleus. Sci. Rep. 2019, 9, 6430. [Google Scholar] [CrossRef]
- Shattock, M.J.; Ottolia, M.; Bers, D.M.; Blaustein, M.P.; Boguslavskyi, A.; Bossuyt, J.; Bridge, J.H.; Chen-Izu, Y.; Clancy, C.E.; Edwards, A.; et al. Na+/Ca2+ exchange and Na+/K+-ATPase in the heart. J. Physiol. 2015, 593, 1361–1382. [Google Scholar] [CrossRef] [PubMed]
- Rizzuto, R.; De Stefani, D.; Raffaello, A.; Mammucari, C. Mitochondria as sensors and regulators of calcium signalling. Nature Rev. Mol. Cell Biol. 2012, 13, 566–578. [Google Scholar] [CrossRef]
- Green, C.B.; Takahashi, J.S.; Bass, J. The meter of metabolism. Cell 2008, 134, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Challet, E. Interactions between light, mealtime and calorie restriction to control daily timing in mammals. J. Comp. Physiol. B 2010, 180, 631–644. [Google Scholar] [CrossRef]
- Yang, J.J.; Cheng, R.C.; Cheng, P.C.; Wang, Y.C.; Huang, R.C. KATP channels mediate differential metabolic responses to glucose shortage of the dorsomedial and ventrolateral oscillators in the central clock. Sci. Rep. 2017, 7, 640. [Google Scholar] [CrossRef]
- Chen, C.H.; Hsu, Y.T.; Chen, C.C.; Huang, R.C. Acid-sensing ion channels in neurons of the rat suprachiasmatic nucleus. J. Physiol. 2009, 587, 1727–1737. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [PubMed]
- Harootunian, A.T.; Kao, J.P.Y.; Eckert, B.K.; Tsien, R.Y. Fluorescence ratio imaging of cytosolic free Na+ in individual fibroblasts and lymphocytes. J. Biol. Chem. 1989, 264, 19458–19467. [Google Scholar] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, R.-C.; Cheng, P.-C.; Wang, Y.-C.; Huang, R.-C. Role of Intracellular Na+ in the Regulation of [Ca2+]i in the Rat Suprachiasmatic Nucleus Neurons. Int. J. Mol. Sci. 2019, 20, 4868. https://doi.org/10.3390/ijms20194868
Cheng R-C, Cheng P-C, Wang Y-C, Huang R-C. Role of Intracellular Na+ in the Regulation of [Ca2+]i in the Rat Suprachiasmatic Nucleus Neurons. International Journal of Molecular Sciences. 2019; 20(19):4868. https://doi.org/10.3390/ijms20194868
Chicago/Turabian StyleCheng, Ruo-Ciao, Pi-Cheng Cheng, Yi-Chi Wang, and Rong-Chi Huang. 2019. "Role of Intracellular Na+ in the Regulation of [Ca2+]i in the Rat Suprachiasmatic Nucleus Neurons" International Journal of Molecular Sciences 20, no. 19: 4868. https://doi.org/10.3390/ijms20194868
APA StyleCheng, R.-C., Cheng, P.-C., Wang, Y.-C., & Huang, R.-C. (2019). Role of Intracellular Na+ in the Regulation of [Ca2+]i in the Rat Suprachiasmatic Nucleus Neurons. International Journal of Molecular Sciences, 20(19), 4868. https://doi.org/10.3390/ijms20194868