Sperm IZUMO1-Dependent Gamete Fusion Influences Male Fertility in Mice


Abstract
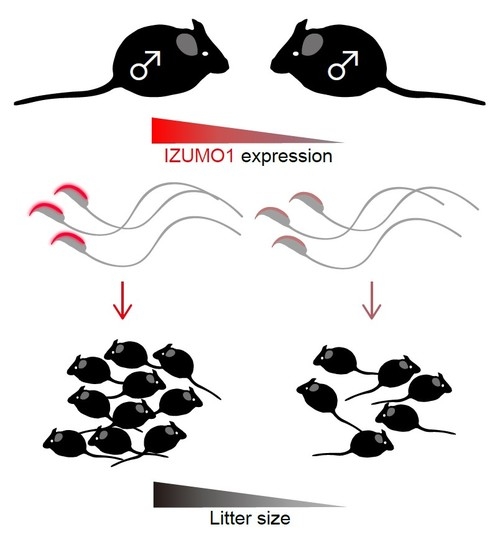
1. Introduction
2. Results
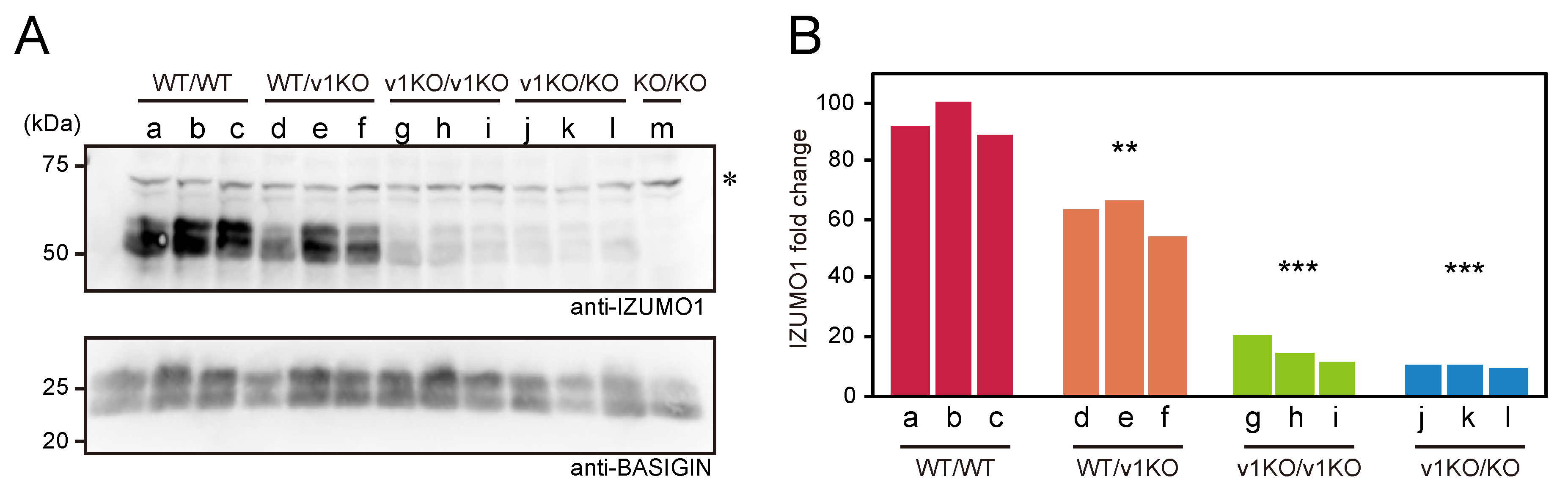
2.1. Comparison of IZUMO1 Protein Quantity Among Each Mouse Line
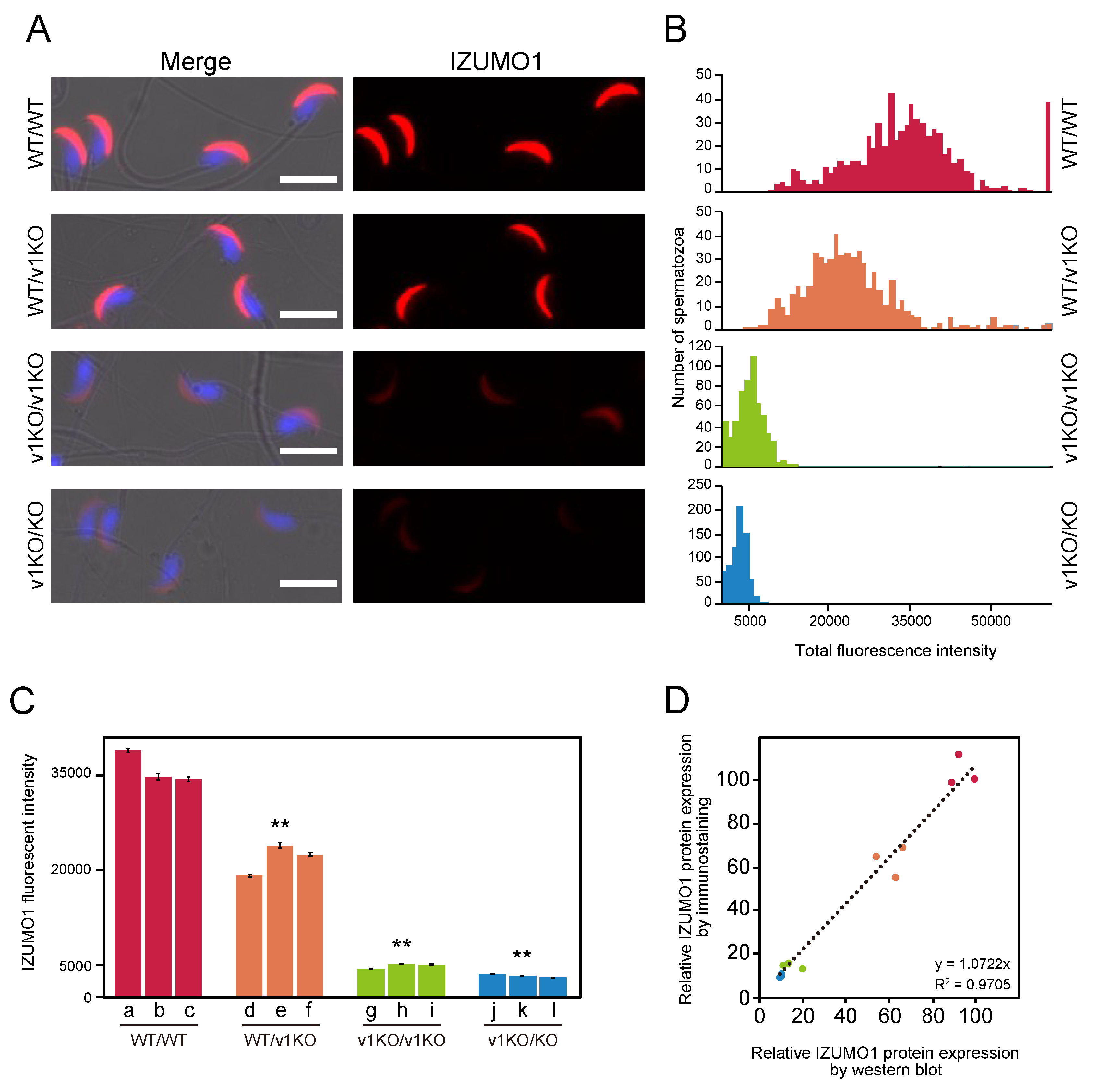
2.2. Immunostaining of Spermatozoa from Each Mouse Line
2.3. Measurement of IZUMO1 Protein by Immunostaining
2.4. Comparison of IZUMO1 Expression Levels Using Different Two Methods
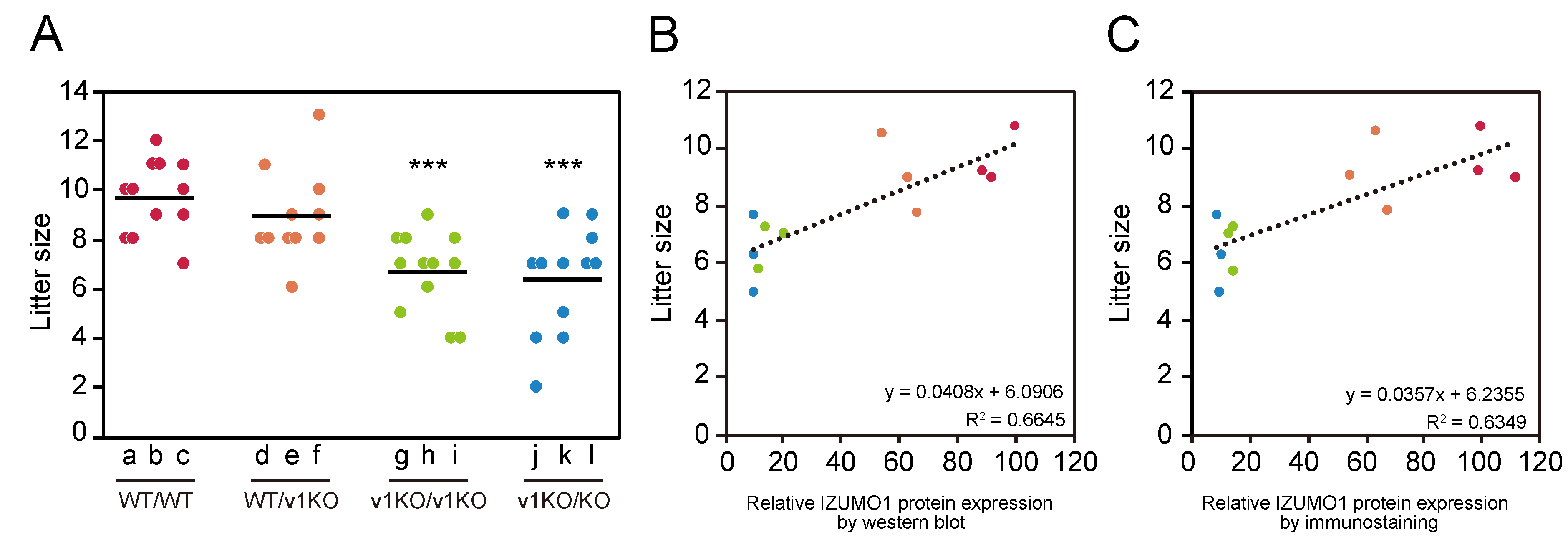
2.5. Average Litter Size from Each Mouse Line
2.6. Correlation of IZUMO1 Protein Amount with Litter Size
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Genotyping
4.3. Measurement of Litter Size
4.4. Western Blot Analysis of Spermatozoa
4.5. Fluorescence Imaging for Spermatozoa
4.6. Image and Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Yanagimachi, R. Mammalian Fertilization; Raven Press, LTD: New York, NY, USA, 1994. [Google Scholar]
- Tosti, E.; Menezo, Y. Gamete activation: Basic knowledge and clinical applications. Hum. Reprod Update 2016, 22, 420–439. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, Y.; Noda, T.; Kobayashi, K.; Oji, A.; Kobayashi, S.; Matsumura, T.; Larasati, T.; Oura, S.; Kojima-Kita, K.; Yu, Z.; et al. Identification of multiple male reproductive tract-specific proteins that regulate sperm migration through the oviduct in mice. Proc. Nat. Acad. Sci. USA 2019, 116, 18498–18506. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Wang, Z.; Shen, C. An update of the regulatory factors of sperm migration from the uterus into the oviduct by genetically manipulated mice. Mol. Reprod. Dev. 2019, 86, 935–955. [Google Scholar] [CrossRef] [PubMed]
- Kishida, K.; Harayama, H.; Kimura, F.; Murakami, T. Individual differences in the distribution of sperm acrosome-associated 1 proteins among male patients of infertile couples; their possible impact on outcomes of conventional in vitro fertilization. Zygote 2016, 24, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Harayama, H.; Minami, K.; Kishida, K.; Noda, T. Protein biomarkers for male artificial insemination subfertility in bovine spermatozoa. Reprod. Med. Biol. 2017, 16, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Ikawa, M.; Okabe, M. Putative sperm fusion protein IZUMO and the role of N-glycosylation. Biochem. Biophys. Res. Commun. 2008, 377, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Ellerman, D.A.; Pei, J.; Gupta, S.; Snell, W.J.; Myles, D.; Primakoff, P. Izumo is part of a multiprotein family whose members form large complexes on mammalian sperm. Mol. Reprod. Dev. 2009, 76, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Aydin, H.; Sultana, A.; Li, S.; Thavalingam, A.; Lee, J.E. Molecular architecture of the human sperm IZUMO1 and egg JUNO fertilization complex. Nature 2016, 534, 562–565. [Google Scholar] [CrossRef]
- Ohto, U.; Ishida, H.; Krayukhina, E.; Uchiyama, S.; Inoue, N.; Shimizu, T. Structure of IZUMO1-JUNO reveals sperm-oocyte recognition during mammalian fertilization. Nature 2016, 534, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Wada, I.; Inoue, N. Alternative splicing of the Izumo1 gene ensures triggering gamete fusion in mice. Sci. Rep. 2019, 9, 3151. [Google Scholar] [CrossRef] [PubMed]
- Satouh, Y.; Inoue, N.; Ikawa, M.; Okabe, M. Visualization of the moment of mouse sperm-egg fusion and dynamic localization of IZUMO1. J. Cell Sci. 2012, 125, 4985–4990. [Google Scholar] [CrossRef]
- Inoue, N.; Wada, I. Monitoring dimeric status of IZUMO1 during the acrosome reaction in living spermatozoon. Cell Cycle Georget. Tex. 2018, 17, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, A.; Mochida, K.; Inoue, H.; Noda, Y.; Endo, T.; Watanabe, G.; Ogura, A. High-Yield Superovulation in Adult Mice by Anti-Inhibin Serum Treatment Combined with Estrous Cycle Synchronization. Biol. Reprod. 2016, 94, 21. [Google Scholar] [CrossRef] [PubMed]
- Lorenzetti, D.; Poirier, C.; Zhao, M.; Overbeek, P.A.; Harrison, W.; Bishop, C.E. A transgenic insertion on mouse chromosome 17 inactivates a novel immunoglobulin superfamily gene potentially involved in sperm-egg fusion. Mamm. Genome 2014, 25, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, R.J.; Lunney, J.K.; Johnson, L.A.; Pursel, V.G.; Kraeling, R.R.; Rohrer, G.A.; Dobrinsky, J.R. Perspectives for artificial insemination and genomics to improve global swine populations. Theriogenology 2005, 63, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Sakase, M.; Fukushima, M.; Harayama, H. Changes of IZUMO1 in bull spermatozoa during the maturation, acrosome reaction, and cryopreservation. Theriogenology 2016, 86, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Grayson, P. Izumo1 and Juno: The evolutionary origins and coevolution of essential sperm-egg binding partners. R Soc. Open Sci. 2015, 2, 150296. [Google Scholar] [CrossRef]
- Inoue, N.; Hamada, D.; Kamikubo, H.; Hirata, K.; Kataoka, M.; Yamamoto, M.; Ikawa, M.; Okabe, M.; Hagihara, Y. Molecular dissection of IZUMO1, a sperm protein essential for sperm-egg fusion. Dev. Camb. Engl. 2013, 140, 3221–3229. [Google Scholar] [CrossRef]
- Chalbi, M.; Barraud-Lange, V.; Ravaux, B.; Howan, K.; Rodriguez, N.; Soule, P.; Ndzoudi, A.; Boucheix, C.; Rubinstein, E.; Wolf, J.P.; et al. Binding of sperm protein Izumo1 and its egg receptor Juno drives Cd9 accumulation in the intercellular contact area prior to fusion during mammalian fertilization. Dev. Camb. Engl. 2014, 141, 3732–3739. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Hagihara, Y.; Wright, D.; Suzuki, T.; Wada, I. Oocyte-triggered dimerization of sperm IZUMO1 promotes sperm-egg fusion in mice. Nat. Commun. 2015, 6, 8858. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Oda, S.; Shikano, T.; Ohnuki, T.; Uematsu, Y.; Sakagami, J.; Tada, N.; Miyazaki, S.; Kudo, A. The gamete fusion process is defective in eggs of Cd9-deficient mice. Nat. Genet. 2000, 24, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Le Naour, F.; Rubinstein, E.; Jasmin, C.; Prenant, M.; Boucheix, C. Severely reduced female fertility in CD9-deficient mice. Science 2000, 287, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Miyado, K.; Yamada, G.; Yamada, S.; Hasuwa, H.; Nakamura, Y.; Ryu, F.; Suzuki, K.; Kosai, K.; Inoue, K.; Ogura, A.; et al. Requirement of CD9 on the egg plasma membrane for fertilization. Science 2000, 287, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K. Vaccine for human contraception targeting sperm Izumo protein and YLP12 dodecamer peptide. Protein Sci. Publ. Protein Soc. 2014, 23, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Naz, R.K. Immunocontraceptive effect of Izumo and enhancement by combination vaccination. Mol. Reprod. Dev. 2008, 75, 336–344. [Google Scholar] [CrossRef]
- An, G.; Huang, T.H.; Wang, D.G.; Xie, Q.D.; Ma, L.; Chen, D.Y. In vitro and in vivo studies evaluating recombinant plasmid pCXN2-mIzumo as a potential immunocontraceptive antigen. Am. J. Reprod. Immunol. 2009, 61, 227–235. [Google Scholar] [CrossRef]
- Wang, M.; Lv, Z.; Shi, J.; Hu, Y.; Xu, C. Immunocontraceptive potential of the Ig-like domain of Izumo. Mol. Reprod. Dev. 2009, 76, 794–801. [Google Scholar] [CrossRef]
- Shrestha, A.; Wadhwa, N.; Gupta, S.K. Evaluation of recombinant fusion protein comprising dog zona pellucida glycoprotein-3 and Izumo and individual fragments as immunogens for contraception. Vaccine 2014, 32, 564–571. [Google Scholar] [CrossRef]
- Xue, F.; Wang, L.; Liu, Y.; Tang, H.; Xu, W.; Xu, C. Vaccination with an Epitope Peptide of IZUMO1 to Induce Contraception in Female Mice. Am. J. Reprod. Immunol. 2016, 75, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT/WT | WT/v1KO | v1KO/v1KO | v1KO/KO | |
|---|---|---|---|---|
| IZUMO1_v1 | +/+ | +/- | -/- | -/- |
| IZUMO1_v2 | +/+ | +/+ | +/+ | +/- |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saito, T.; Wada, I.; Inoue, N. Sperm IZUMO1-Dependent Gamete Fusion Influences Male Fertility in Mice. Int. J. Mol. Sci. 2019, 20, 4809. https://doi.org/10.3390/ijms20194809
Saito T, Wada I, Inoue N. Sperm IZUMO1-Dependent Gamete Fusion Influences Male Fertility in Mice. International Journal of Molecular Sciences. 2019; 20(19):4809. https://doi.org/10.3390/ijms20194809
Chicago/Turabian StyleSaito, Takako, Ikuo Wada, and Naokazu Inoue. 2019. "Sperm IZUMO1-Dependent Gamete Fusion Influences Male Fertility in Mice" International Journal of Molecular Sciences 20, no. 19: 4809. https://doi.org/10.3390/ijms20194809
APA StyleSaito, T., Wada, I., & Inoue, N. (2019). Sperm IZUMO1-Dependent Gamete Fusion Influences Male Fertility in Mice. International Journal of Molecular Sciences, 20(19), 4809. https://doi.org/10.3390/ijms20194809