In Vitro Evaluation of Novel Hybrid Cooperative Complexes in a Wound Healing Model: A Step Toward Improved Bioreparation

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
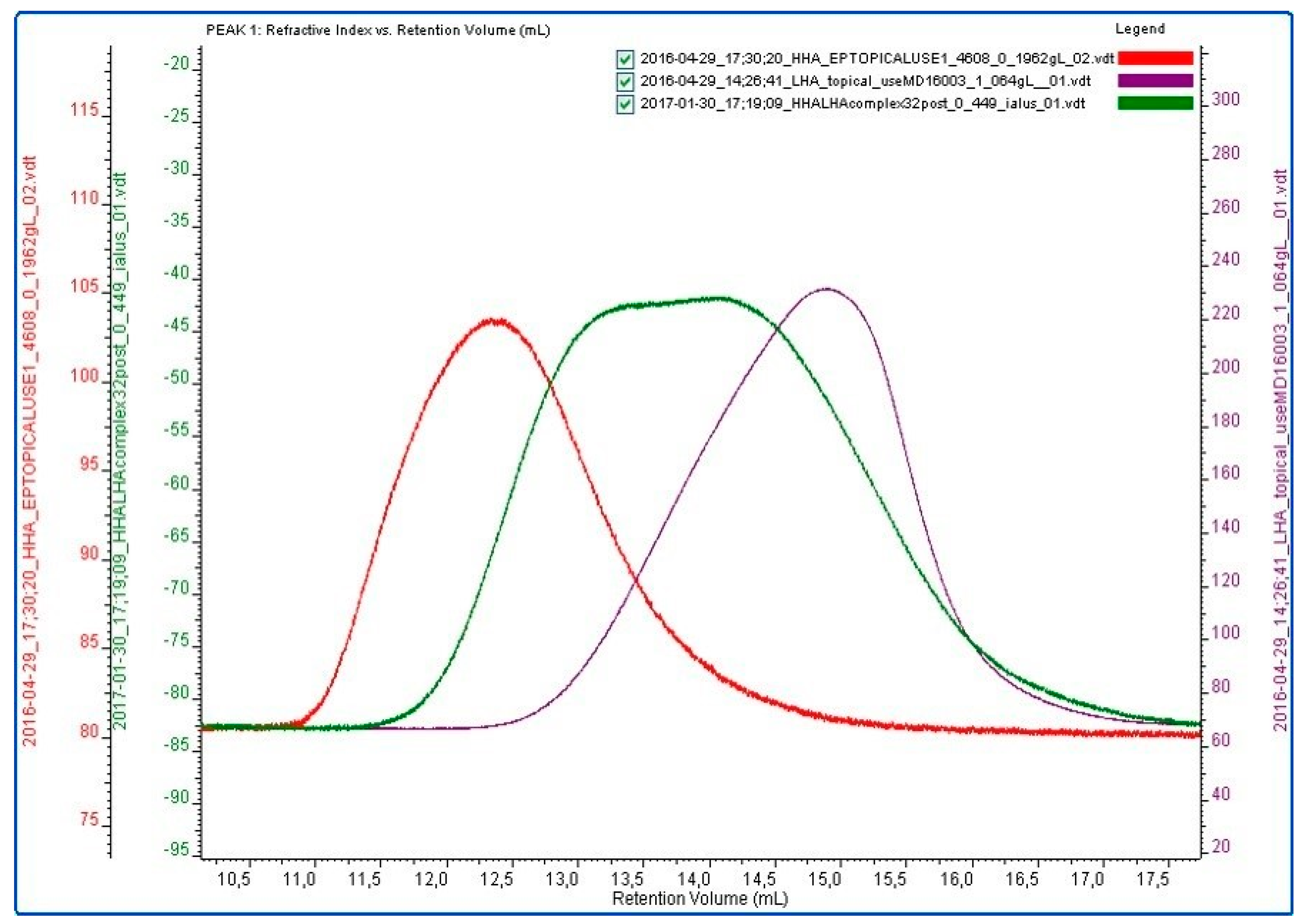
2.1. Hydrodynamic and Rheological Characterization
2.2. Biological Response of HaCaT/HDF to HA-Based Treatments
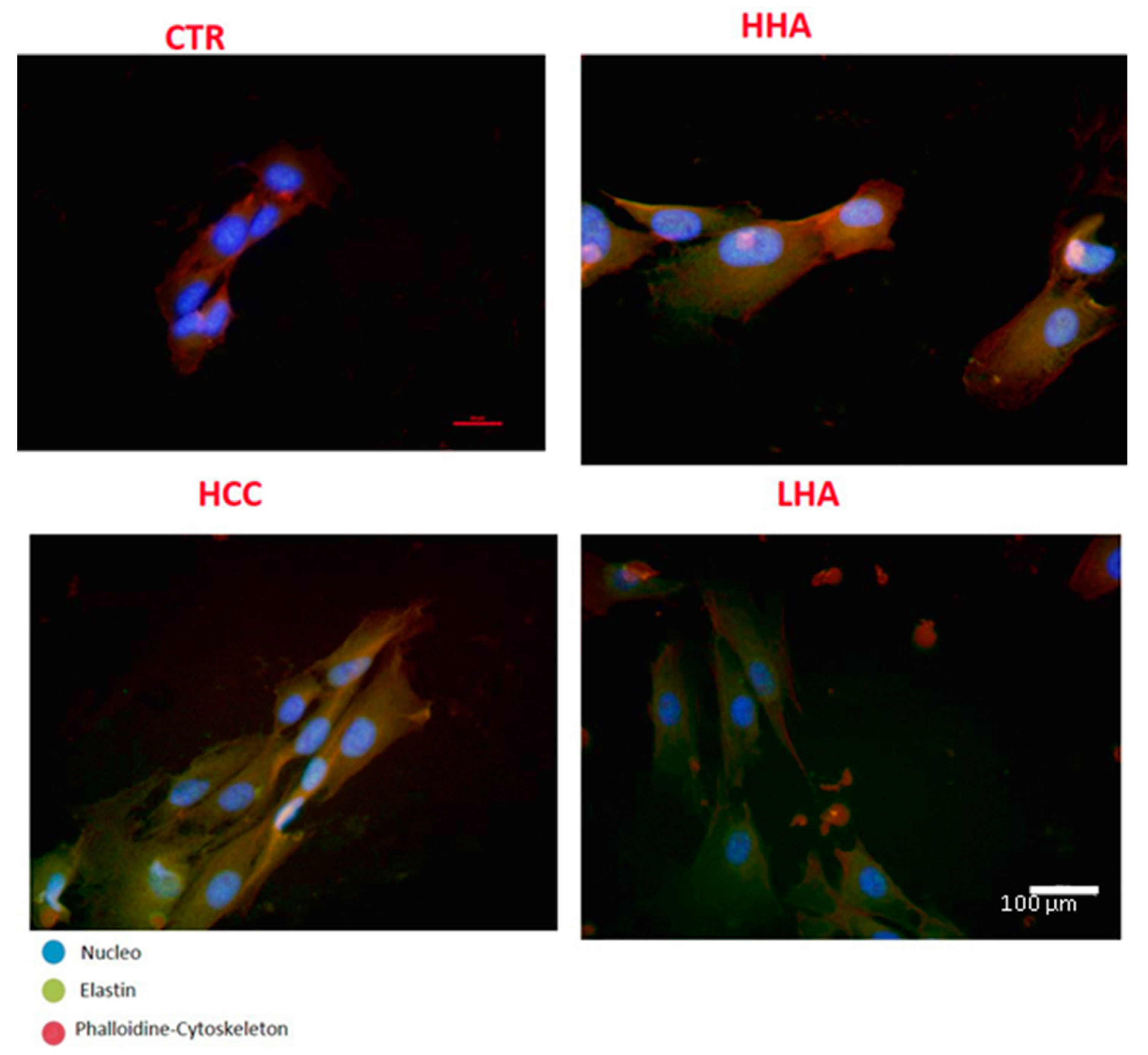
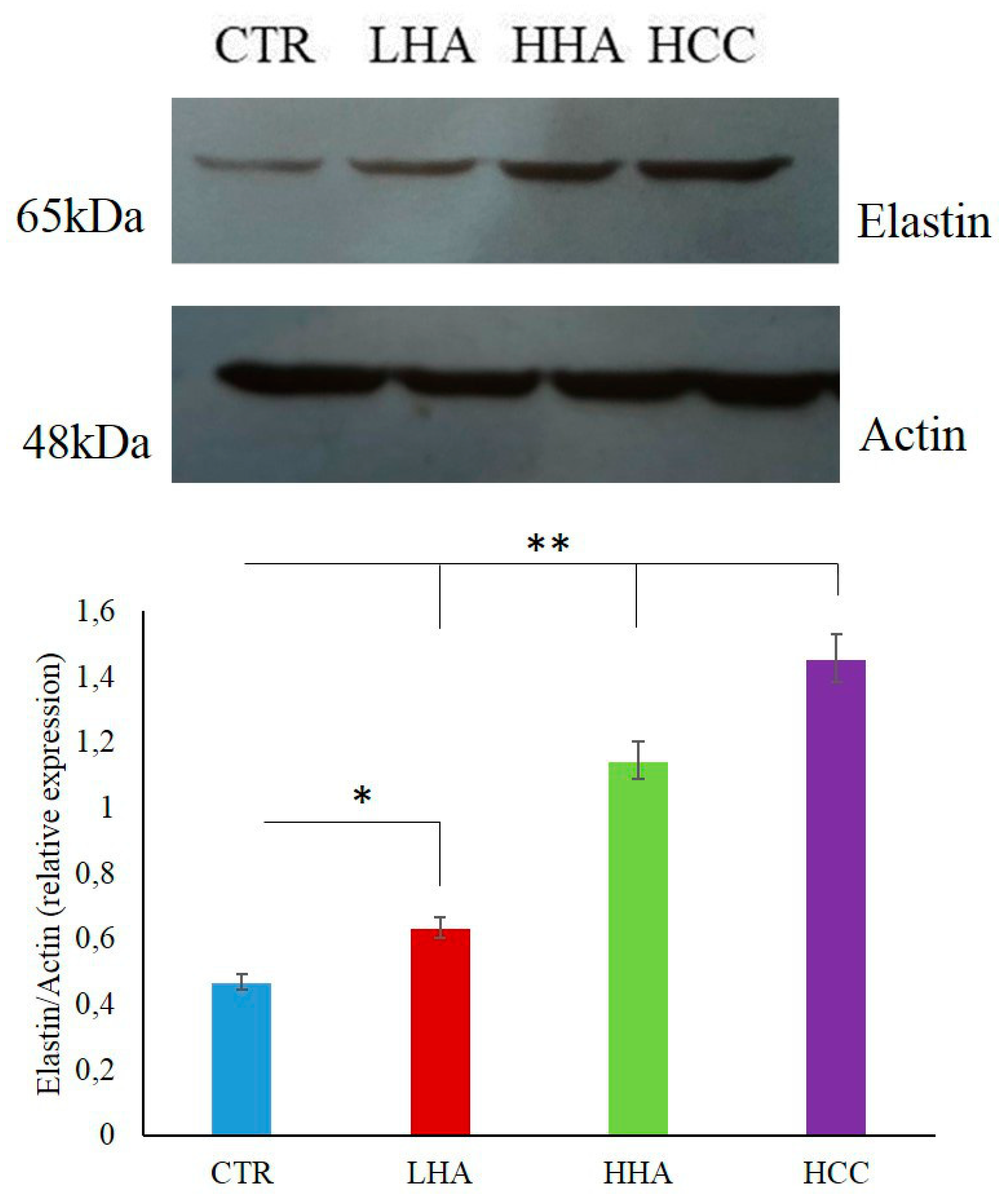
2.2.1. Elastin Expression
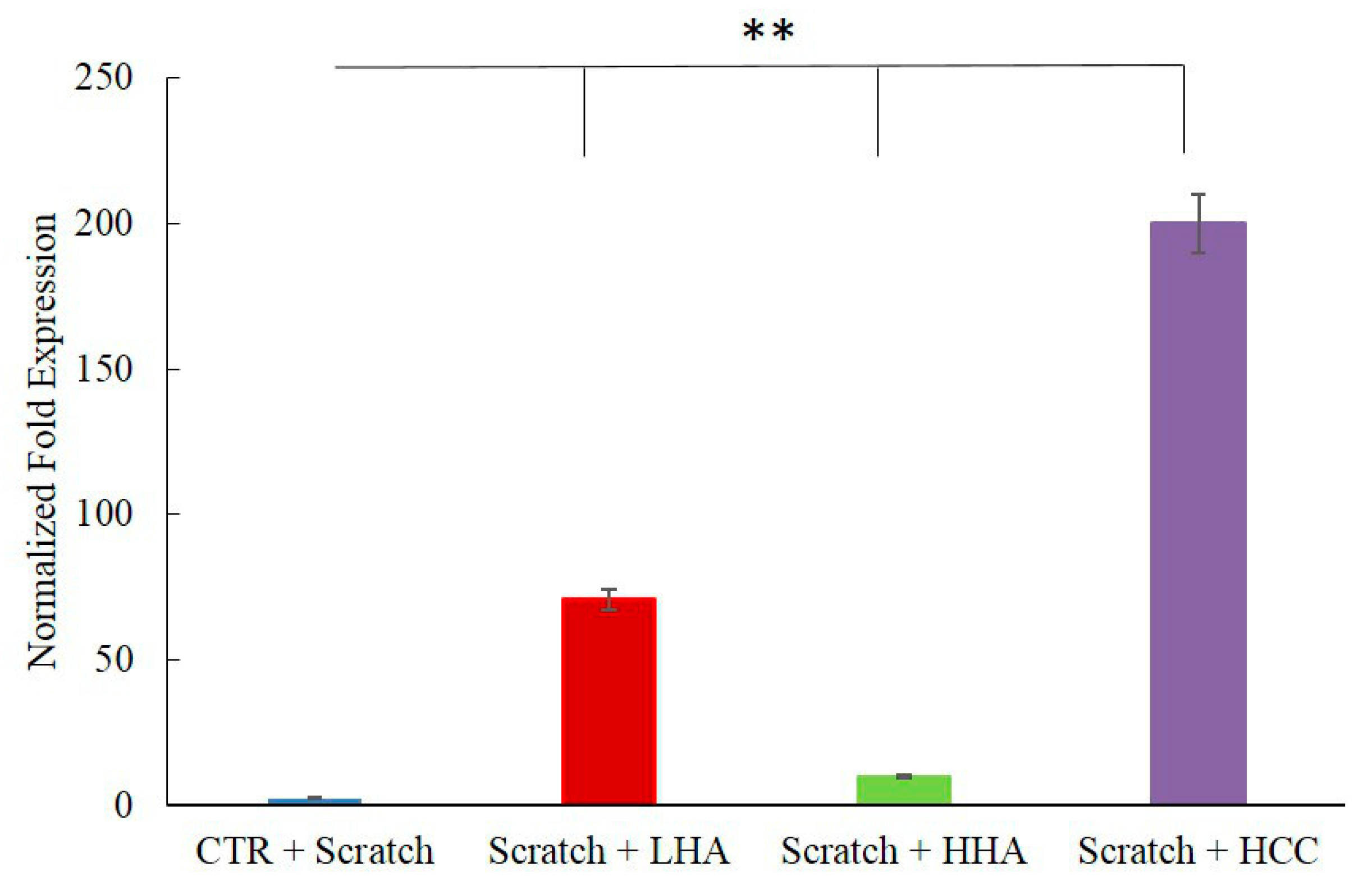
2.2.2. HBD-2 Modulation
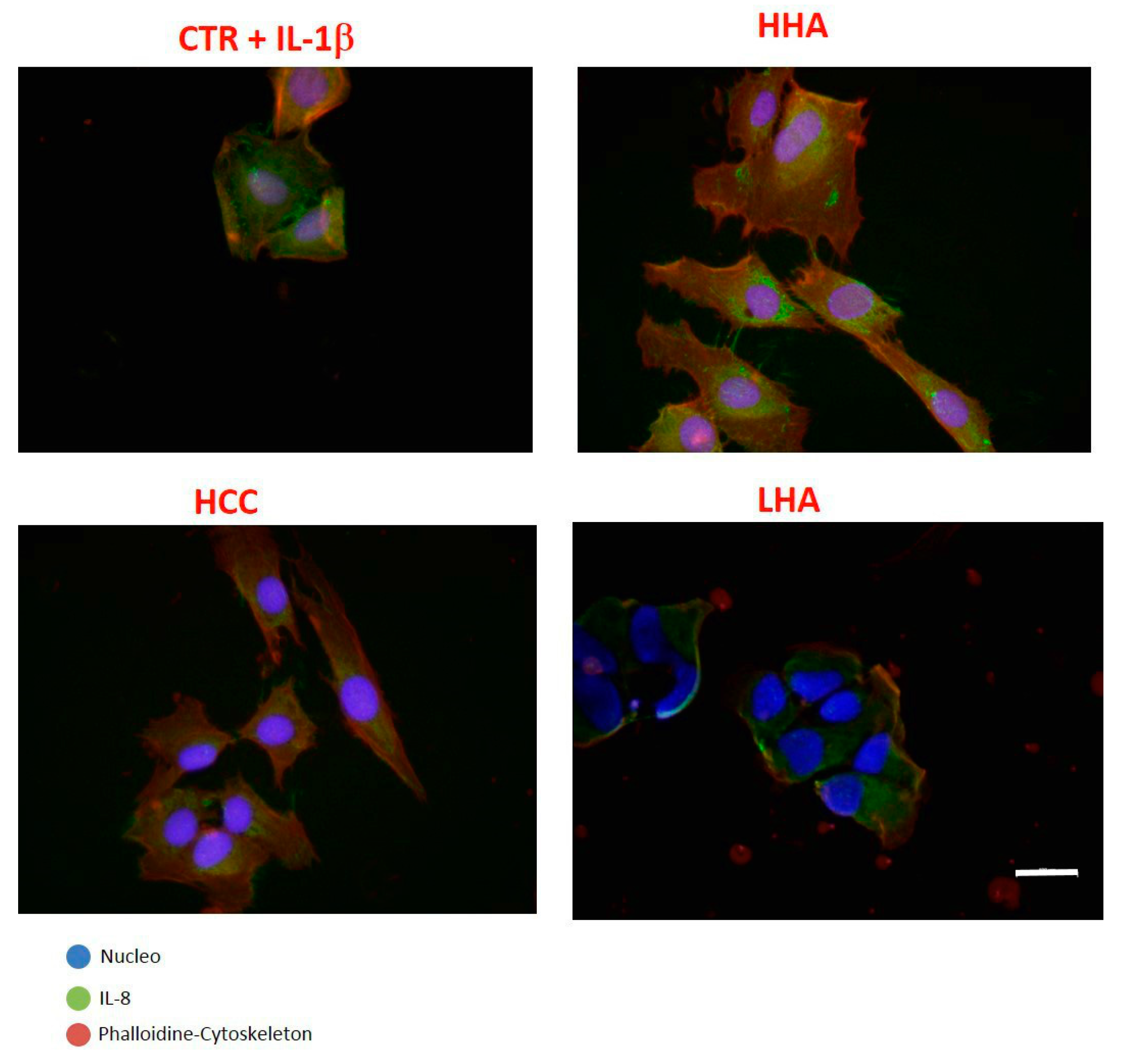
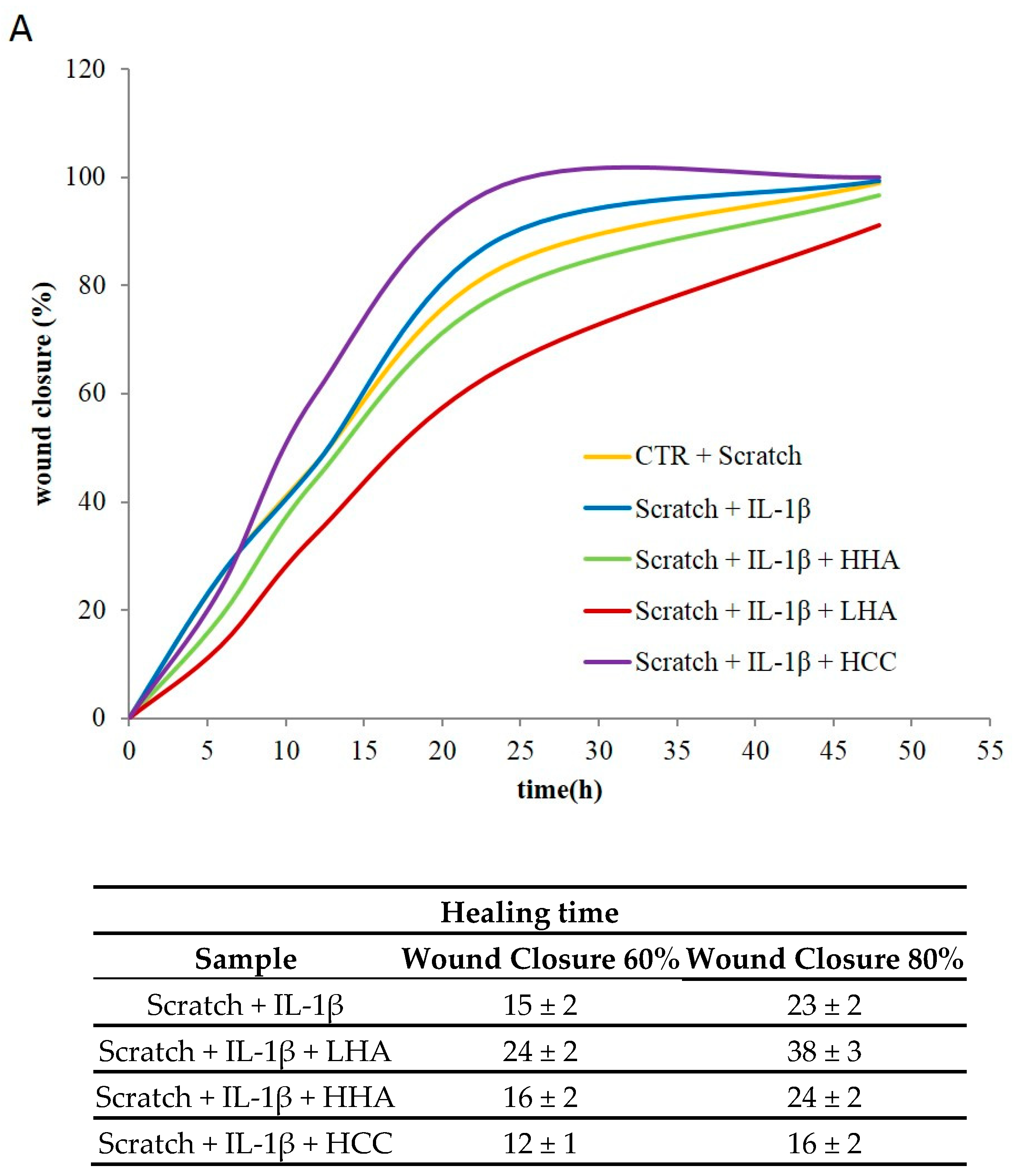
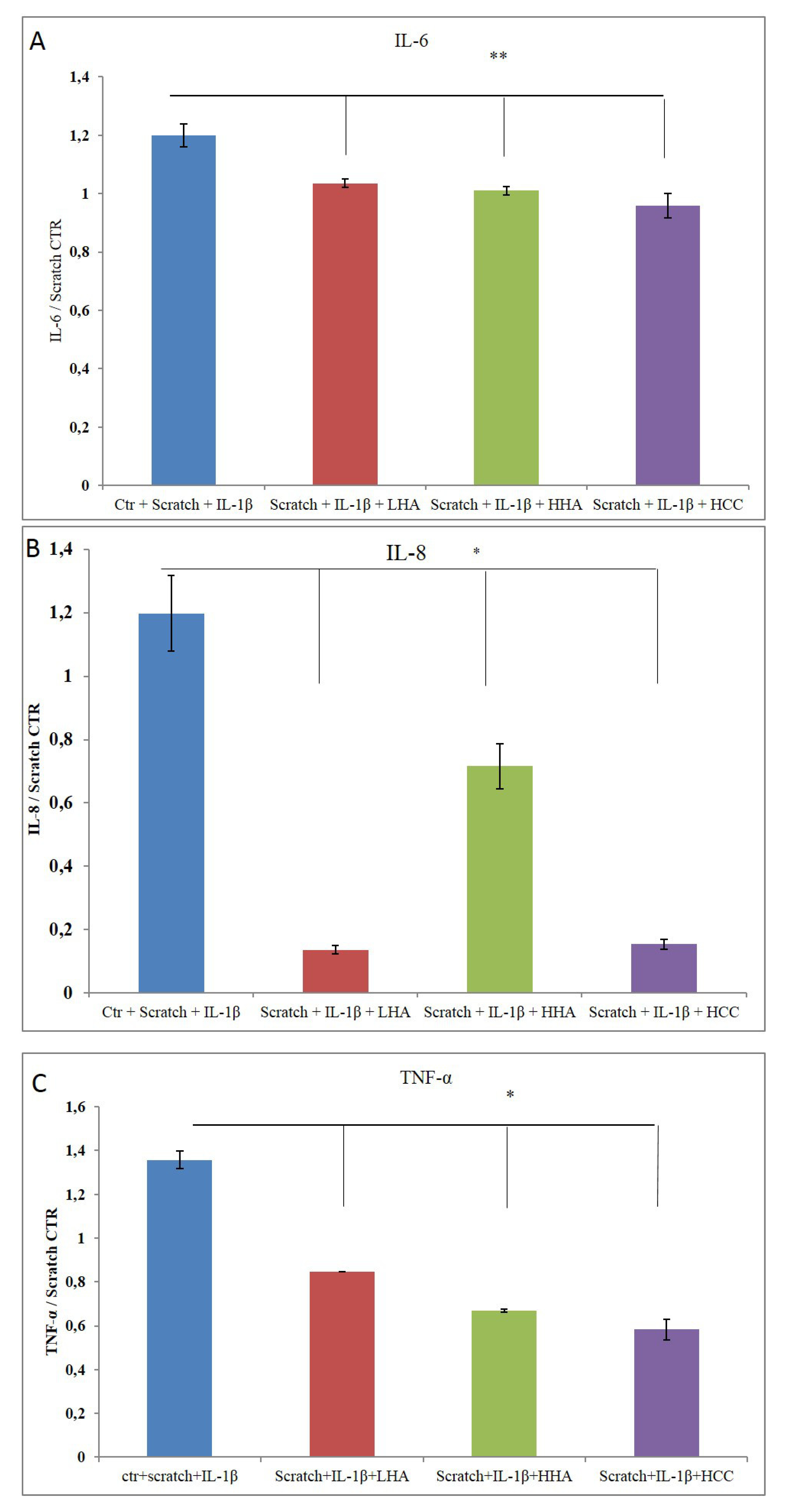
2.2.3. Inflammation and Wound Closure
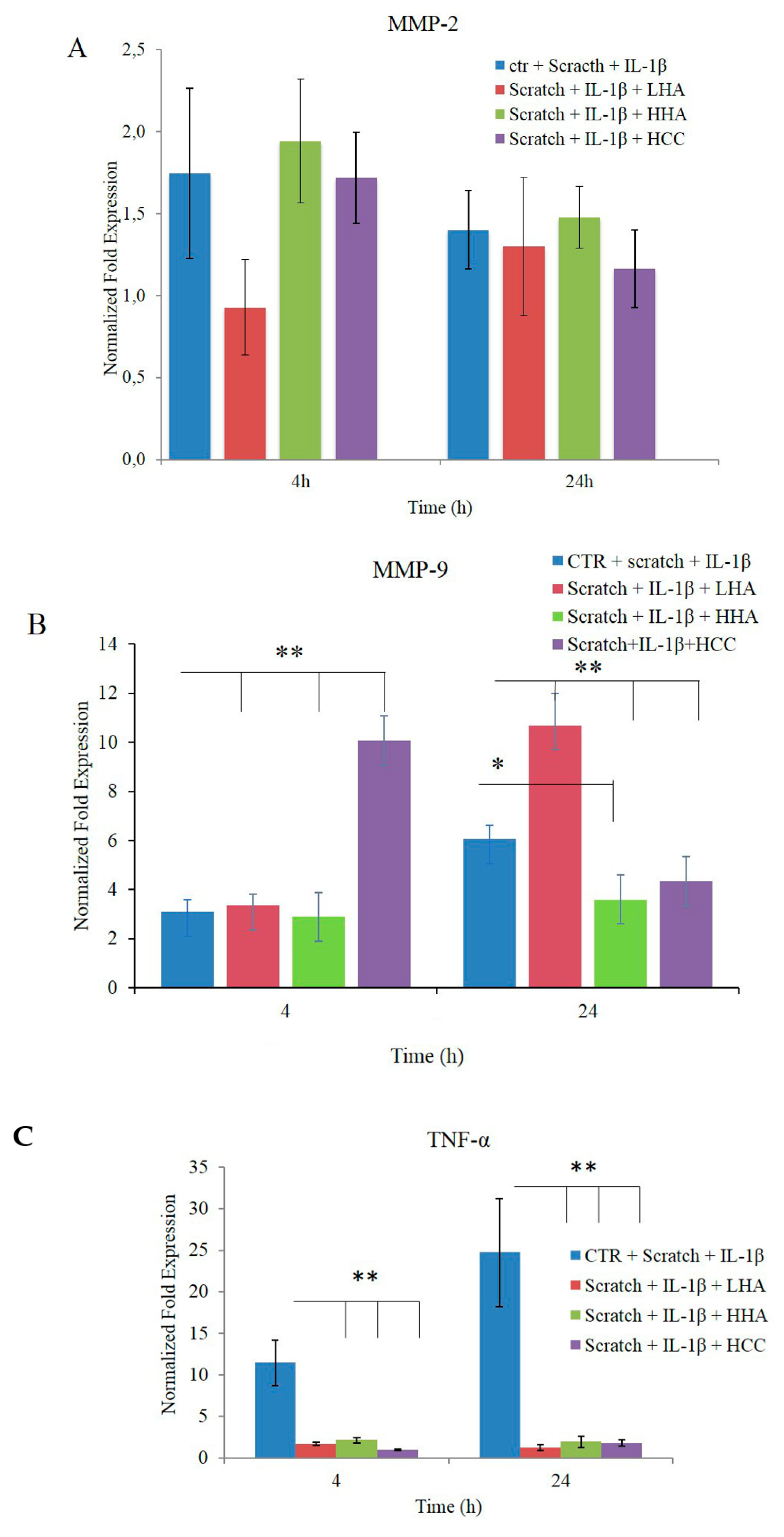
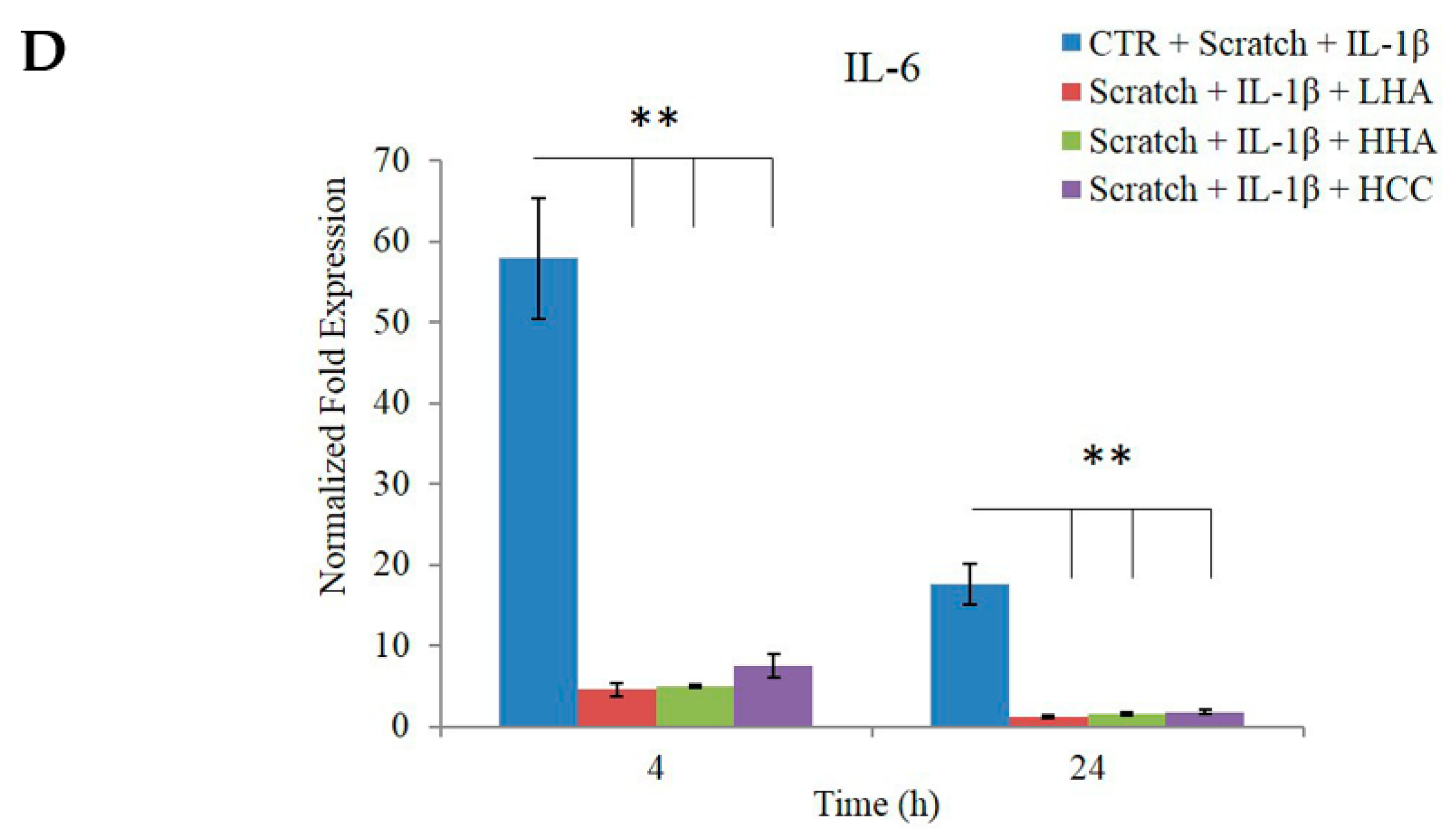
2.2.4. Bio-Plex Assay
3. Discussion
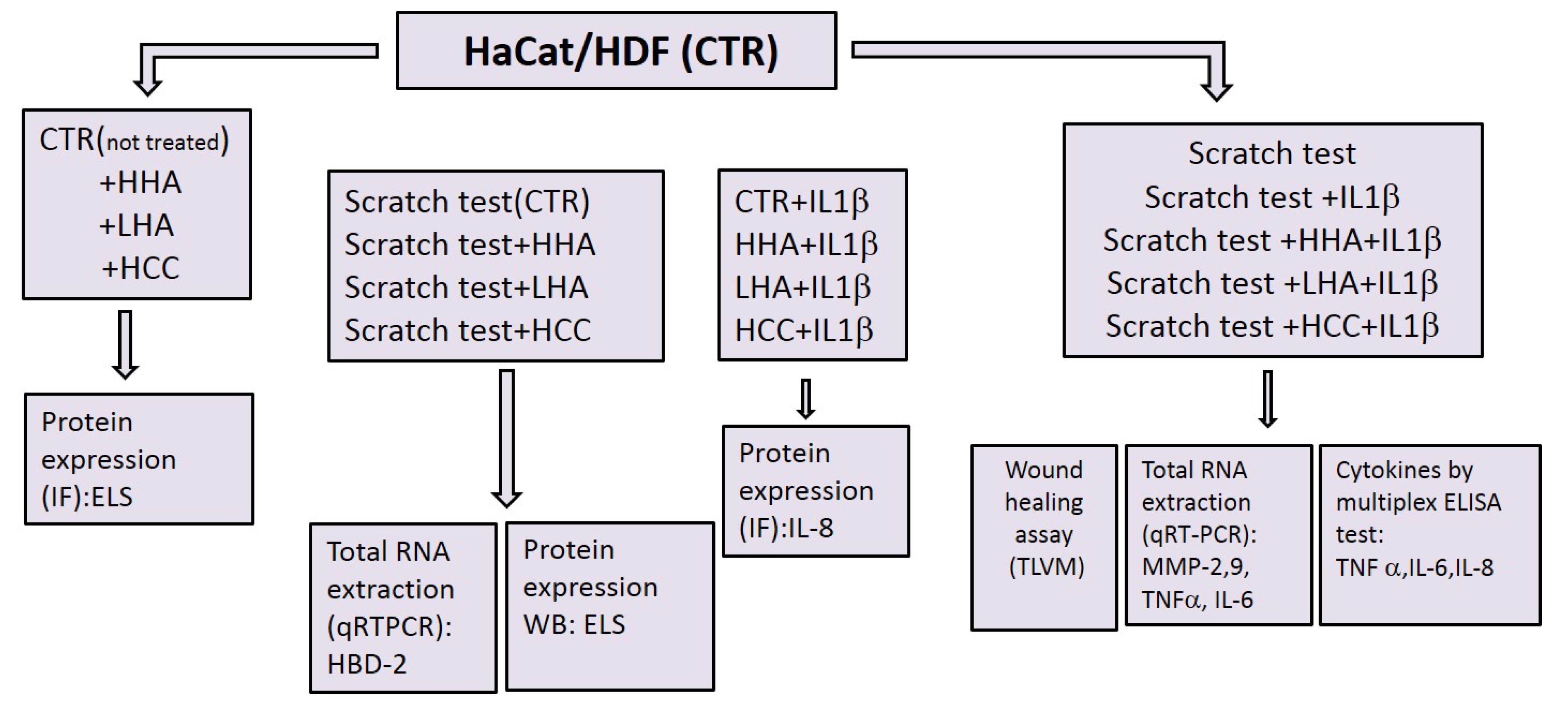
4. Materials and Methods
4.1. Materials
4.2. Sample Preparation
4.3. Hydrodynamic Characterization and Rheological Measurements
4.4. In Vitro Scratch Test
4.5. Quantitative RT-PCR
4.6. Western Blotting
4.7. Immunofluorescent Staining
4.8. Cytokine Quantification Using Bio-Plex
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Balazs, E.A.; Denlinger, J.L. Clinical uses of hyaluronan. Ciba. Found Symp. 1989, 143, 265–280. [Google Scholar] [PubMed]
- La Gatta, A.; Schiraldi, C.; Papa, A.; De Rosa, M. Comparative analysis of commercial dermal fillers based on crosslinked hyaluronan: Physical characterization and in vitro enzymatic degradation. Polym. Degrad. Stabil. 2011, 96, 630–636. [Google Scholar] [CrossRef]
- La Gatta, A.; De Rosa, M.; Frezza, M.A.; Catalano, C.; Meloni, M.; Schiraldi, C. Biophysical and biological characterization of a new line of hyaluronan-based dermal fillers: a scientific rationale to specific clinical indications. Mater. Sci. Eng. C.Mater. Biol. Appl. 2016, 68, 565–572. [Google Scholar] [CrossRef]
- Powers, J.G.; Higham, C.; Broussard, K.; Phillips, T.J. Wound healing and treating wounds: Chronic wound care and management. J. Am. Acad. Dermatol. 2016, 74, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Rowan, M.P.; Cancio, L.C.; Elster, E.A.; Burmeister, D.M.; Rose, L.F.; Natesan, S.; Chung, K.K. Burn wound healing and treatment: review and advancements. Crit. care 2015, 19, 243. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, B.A.; Hoey, C. Topical antimicrobial therapy for treating chronic wounds. Clin. Infect. Dis. 2009, 49, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Atiyeh, B.S.; Costagliola, M.; Hayek, S.N.; Dibo, S.A. Effect of silver on burn wound infection control and healing: review of the literature. Burns 2007, 33, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Gisby, J.; Bryant, J. Efficacy of a new cream formulation of mupirocin: comparison with oral and topical agents in experimental skin infections. Antimicrob. Agents Chemother. 2000, 44, 255–260. [Google Scholar] [CrossRef]
- Mogoşanu, G.D.; Grumezescu, A.M. Natural and synthetic polymers for wounds and burns dressing. Int. J. Pharm. 2014, 463, 127–136. [Google Scholar] [CrossRef]
- Ng, S.F.; Leow, H.L. Development of biofilm-targeted antimicrobial wound dressing for the treatment of chronic wound infections. Drug Dev. Ind. Pharm. 2015, 41, 1902–1909. [Google Scholar] [CrossRef]
- Morsi, N.M.; Abdelbary, G.A.; Ahmed, M.A. Silver sulfadiazine based cubosome hydrogels for topical treatment of burns: development and in vitro/in vivo characterization. Eur. J. Pharm. Biopharm. 2014, 86, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Lira, A.A.M.; Sester, E.A.; Carvalho, A.L.M.; Strattmann, R.R.; Albuquerque, M.M.; Wanderley, A.G.; Santana, D.P. Development of lapachol topical formulation: anti-inflammatory study of a selected formulation. AAPS PharmSciTech. 2008, 9, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Neuman, M.G.; Nanau, R.M.; Oruña-Sanchez, L.; Coto, G. Hyaluronic acid and wound healing. J. Pharm. Pharm. Sci. 2015, 18, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Essendoubi, M.; Gobinet, C.; Reynaud, R.; Angiboust, J.F.; Manfait, M.; Piot, O. Human skin penetration of hyaluronic acid of different molecular weights as probed by Raman spectroscopy. Skin Res. Technol. 2016, 22, 55–62. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, A.; Stellavato, A.; Corsuto, L.; Diana, P.; Filosa, R.; La Gatta, A.; De Rosa, M.; Schiraldi, C. Is molecular size a discriminating factor in hyaluronan interaction with human cells? Carbohydr. Polym. 2017, 157, 21–30. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, M.; D’Agostino, A.; La Gatta, A.; Schiraldi, C. Hybrid cooperative complexes of hyaluronic acid. U.S. Patent Application No. 13/820,838, 2011. [Google Scholar]
- Khameneh, H.J.; Isa, S.A.; Min, L.; Nih, F.W.; Ruedl, C. GM-CSF Signalling Boosts Dramatically IL-1Production. PLoS ONE 2011, 6, e23025. [Google Scholar] [CrossRef] [PubMed]
- Basso, F.G.; Pansani, T.N.; Turrioni, A.P.S.; Soares, D.G.; de Souza Costa, C.A.; Hebling, J. Tumor Necrosis Factor-α and Interleukin (IL)-1β, IL-6, and IL-8 Impair In Vitro Migration and Induce Apoptosis of Gingival Fibroblasts and Epithelial Cells, Delaying Wound Healing. J. Periodontol. 2016, 87, 990–996. [Google Scholar] [CrossRef]
- Gougeon, P.Y.; Lourenssen, S.; Han, T.Y.; Nair, D.G.; Ropeleski, M.J.; Blennerhassett, M.G. The pro-inflammatory cytokines IL-1β and TNFα are neurotrophic for enteric neurons. J. Neurosci. 2013, 33, 3339–3351. [Google Scholar] [CrossRef] [PubMed]
- Stellavato, A.; De Novellis, F.; Reale, S.; De Rosa, M.; Schiraldi, C. Hybrid complexes of high and low molecular weight: evaluation using an in vitro model of osteoarthritis. J. Biol. Regul. Homeost. Agents 2016, 30, 7–16. [Google Scholar] [PubMed]
- Fusco, A.; Savio, V.; Cammarota, M.; Alfano, A.; Schiraldi, C.; Donnarumma, G. Beta-Defensin-2 and Beta-Defensin-3 Reduce Intestinal Damage Caused by Salmonella typhimurium Modulating the Expression of Cytokines and Enhancing the Probiotic Activity of Enterococcus faecium. J. Immunol. Res. 2017, 2017, 6976935. [Google Scholar] [CrossRef]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.; et al. Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Ogawa, H.; Nagaoka, I. Human beta-defensin-2 functions as a chemotactic agent for tumour necrosis factor-alpha-treated human neutrophils. Immunology 2004, 111, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Rohrl, J.; Yang, D.; Oppenheim, J.J.; Hehlgans, T. Human {beta}-Defensin 2 and 3 and Their Mouse Orthologs Induce Chemotaxis through Interaction with CCR2. J. Immunol. 2010, 184, 6688–6694. [Google Scholar] [CrossRef] [PubMed]
- Illescas-Montes, R.; Melguizo-Rodríguez, L.; García-Martínez, O.; de Luna-Bertos, E.; Manzano-Moreno, F.J.; Ruiz, C.; Ramos-Torrecillas, J. Human Fibroblast Gene Expression Modulation Using 940 NM Diode Laser. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Stellavato, A.; Corsuto, L.; D’Agostino, A.; La Gatta, A.; Diana, P.; Bernini, P.; De Rosa, M.; Schiraldi, C. Hyaluronan Hybrid Cooperative Complexes as a Novel Frontier for Cellular Bioprocesses Re-Activation. PloS One 2016, 11, e0163510. [Google Scholar] [CrossRef] [PubMed]
- Stellavato, A.; La Noce, M.; Corsuto, L.; Pirozzi, A.V.A.; De Rosa, M.; Papaccio, G.; Schiraldi, C.; Tirino, V. Hybrid Complexes of High and Low Molecular Weight Hyaluronans Highly Enhance HASCs Differentiation: Implication for Facial Bioremodelling. Cell. Physiol. Biochem. 2017, 44, 1078–1092. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, J.S. The role of hyaluronan in wound healing. Int. Wound J. 2014, 11, 159–163. [Google Scholar] [CrossRef]
- Caló, E.; Khutoryanskiy, V.V. Biomedical applications of hydrogels: A review of patents and commercial products. Eur. Polym. J. 2015, 65, 252–267. [Google Scholar] [CrossRef]
- Almine, J.F.; Wise, S.G.; Weiss, A.S. Elastin signaling in wound repair. Birth Defects Res. Part C: Embryo Today 2012, 96, 248–257. [Google Scholar] [CrossRef]
- Siwik, D.A.; Chang, D.L.; Colucci, W.S. Interleukin-1β and tumor necrosis factor-α decrease collagen synthesis and increase matrix metalloproteinase activity in cardiac fibroblasts in vitro. Circ. Res. 2000, 86, 1259–1265. [Google Scholar] [CrossRef]
- Stellavato, A.; Vassallo, V.; La Gatta, A.; Pirozzi, A.V.A.; De Rosa, M.; Balato, G.; D’Addona, A.; Tirino, V.; Ruosi, C.; Schiraldi, C. Novel Hybrid Gels Made of High and Low Molecular Weight Hyaluronic Acid Induce Proliferation and Reduce Inflammation in an Osteoarthritis In Vitro Model Based on Human Synoviocytes and Chondrocytes. Biomed. Res. Int. 2019, 2019, 4328219. [Google Scholar] [CrossRef] [PubMed]
- Petrey, A.C.; de la Motte, C.A. Hyaluronan, a crucial regulator of inflammation. Front. Immuno. 2014, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Stellavato, A.; Tirino, V.; de Novellis, F.; Della Vecchia, A.; Cinquegrani, F.; De Rosa, M.; Papaccio, G.; Schiraldi, C. Biotechnological chondroitin a novel glycosamminoglycan with remarkable biological function on human primary chondrocytes. J. Cell. Biochem. 2016, 117, 2158–2169. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Ushio, H.; Nakano, N.; Ng, W.; Sayama, K.; Hashimoto, K.; Nagaoka, I.; Okumura, K.; Ogawa, H. Antimicrobial peptides human β-defensins stimulate epidermal keratinocyte migration, proliferation and production of proinflammatory cytokines and chemokines. J. Invest. Dermatol. 2007, 127, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Baroni, A.; Donnarumma, G.; Paoletti, I.; Longanesi-Cattani, I.; Bifulco, K.; Tufano, M.A.; Carriero, M.V. Antimicrobial human beta-defensin-2 stimulates migration, proliferation and tube formation of human umbilical vein endothelial cells. Peptides 2009, 30, 267–272. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples Post Thermal Treatments | Zero-Shear Viscosity (η0) (mPa⋅s) |
|---|---|
| HHA | 94.73 ± 0.46 |
| LHA | 3.95 ± 0.01 |
| HCC | 16.60 ± 0.06 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Agostino, A.; Maritato, R.; La Gatta, A.; Fusco, A.; Reale, S.; Stellavato, A.; Pirozzi, A.V.A.; De Rosa, M.; Donnarumma, G.; Schiraldi, C. In Vitro Evaluation of Novel Hybrid Cooperative Complexes in a Wound Healing Model: A Step Toward Improved Bioreparation. Int. J. Mol. Sci. 2019, 20, 4727. https://doi.org/10.3390/ijms20194727
D’Agostino A, Maritato R, La Gatta A, Fusco A, Reale S, Stellavato A, Pirozzi AVA, De Rosa M, Donnarumma G, Schiraldi C. In Vitro Evaluation of Novel Hybrid Cooperative Complexes in a Wound Healing Model: A Step Toward Improved Bioreparation. International Journal of Molecular Sciences. 2019; 20(19):4727. https://doi.org/10.3390/ijms20194727
Chicago/Turabian StyleD’Agostino, Antonella, Rosa Maritato, Annalisa La Gatta, Alessandra Fusco, Sabrina Reale, Antonietta Stellavato, Anna Virginia Adriana Pirozzi, Mario De Rosa, Giovanna Donnarumma, and Chiara Schiraldi. 2019. "In Vitro Evaluation of Novel Hybrid Cooperative Complexes in a Wound Healing Model: A Step Toward Improved Bioreparation" International Journal of Molecular Sciences 20, no. 19: 4727. https://doi.org/10.3390/ijms20194727
APA StyleD’Agostino, A., Maritato, R., La Gatta, A., Fusco, A., Reale, S., Stellavato, A., Pirozzi, A. V. A., De Rosa, M., Donnarumma, G., & Schiraldi, C. (2019). In Vitro Evaluation of Novel Hybrid Cooperative Complexes in a Wound Healing Model: A Step Toward Improved Bioreparation. International Journal of Molecular Sciences, 20(19), 4727. https://doi.org/10.3390/ijms20194727