SNPs rs11240569, rs708727, and rs823156 in SLC41A1 Do Not Discriminate Between Slovak Patients with Idiopathic Parkinson’s Disease and Healthy Controls: Statistics and Machine-Learning Evidence

 ,
, 
Abstract
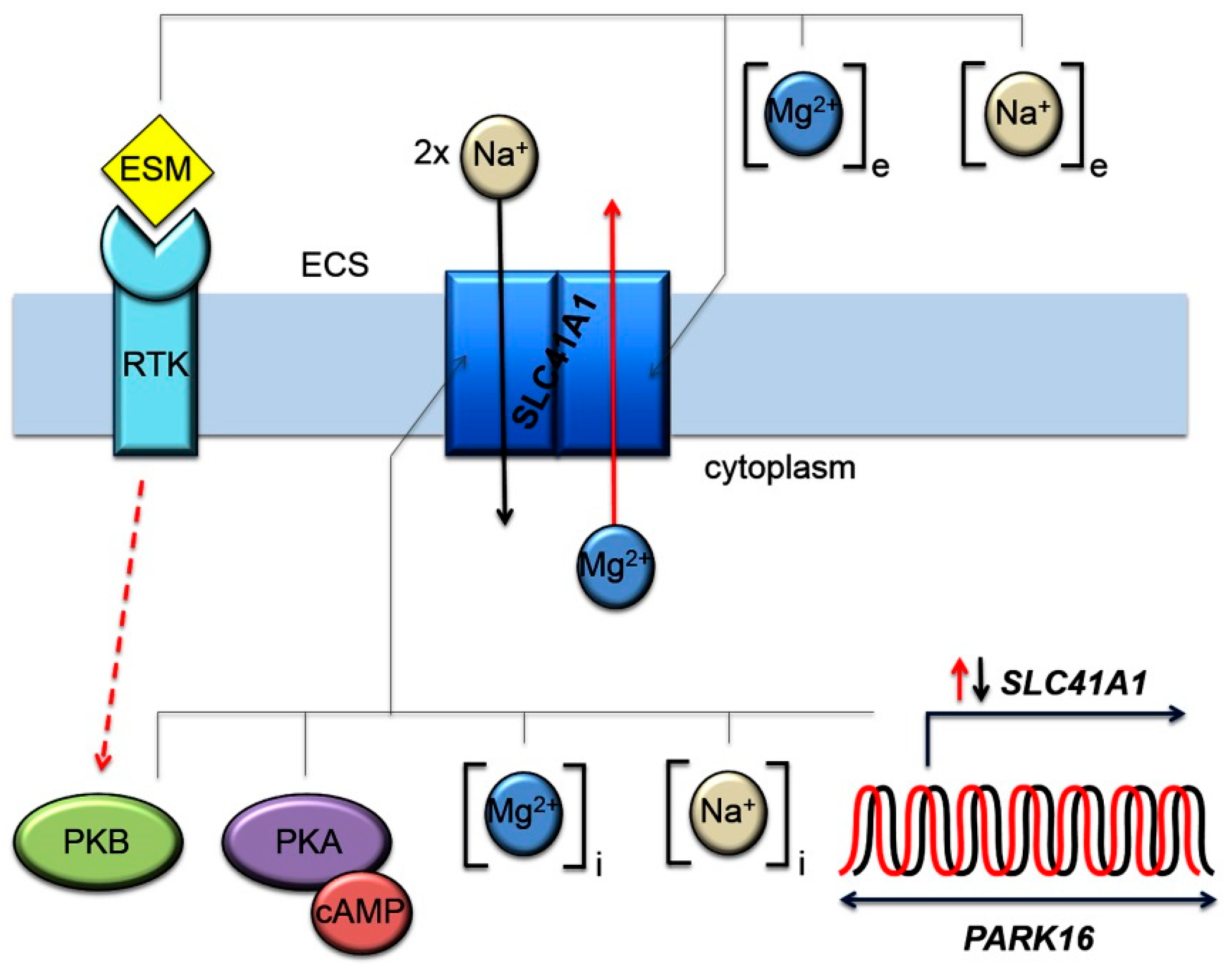
1. Introduction
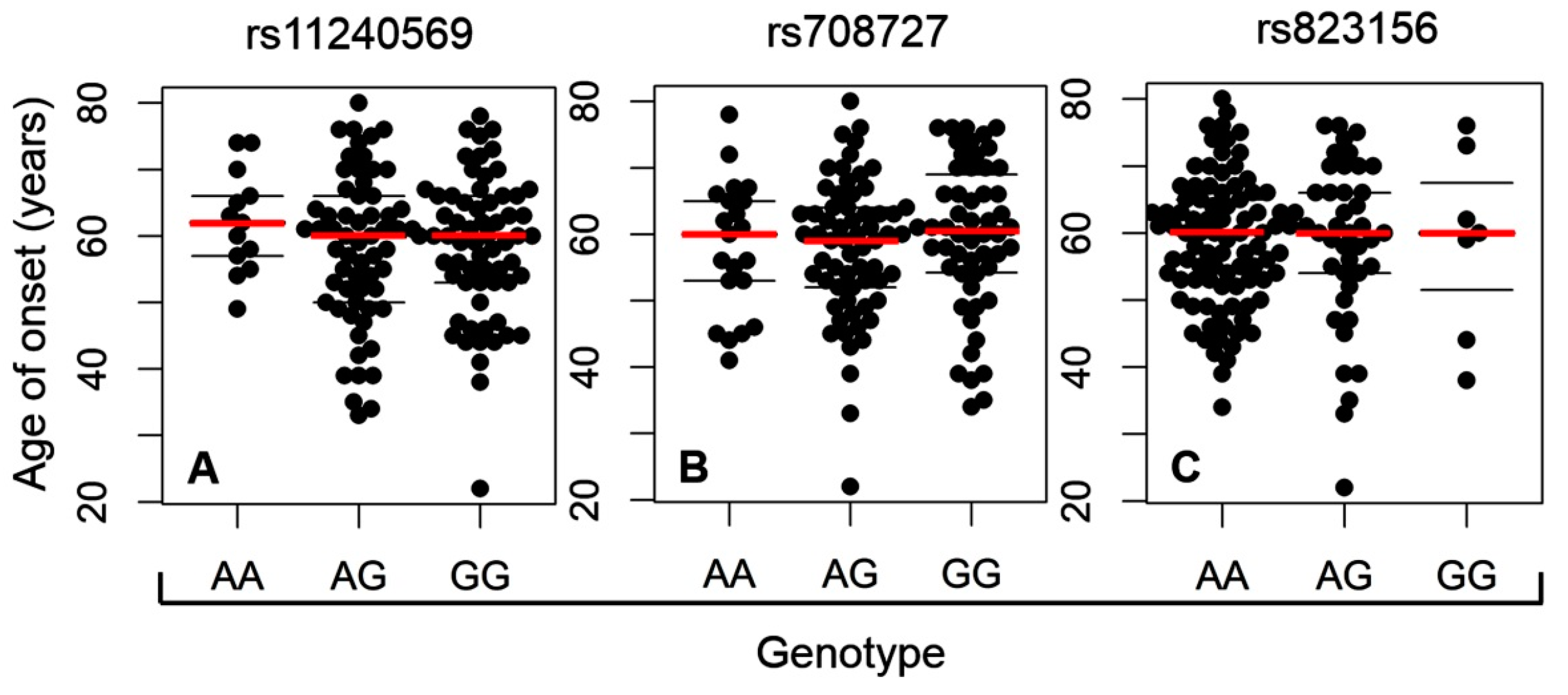
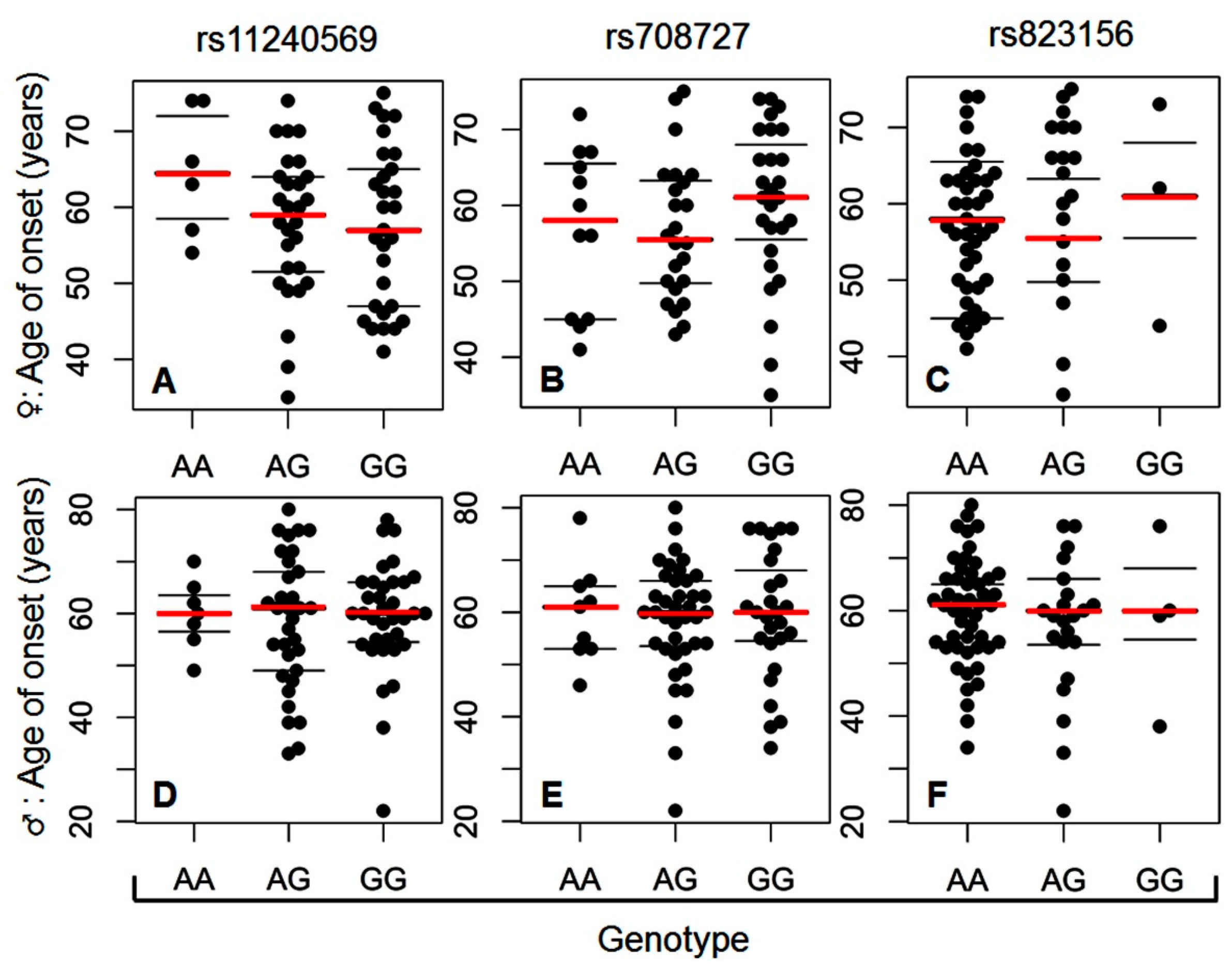
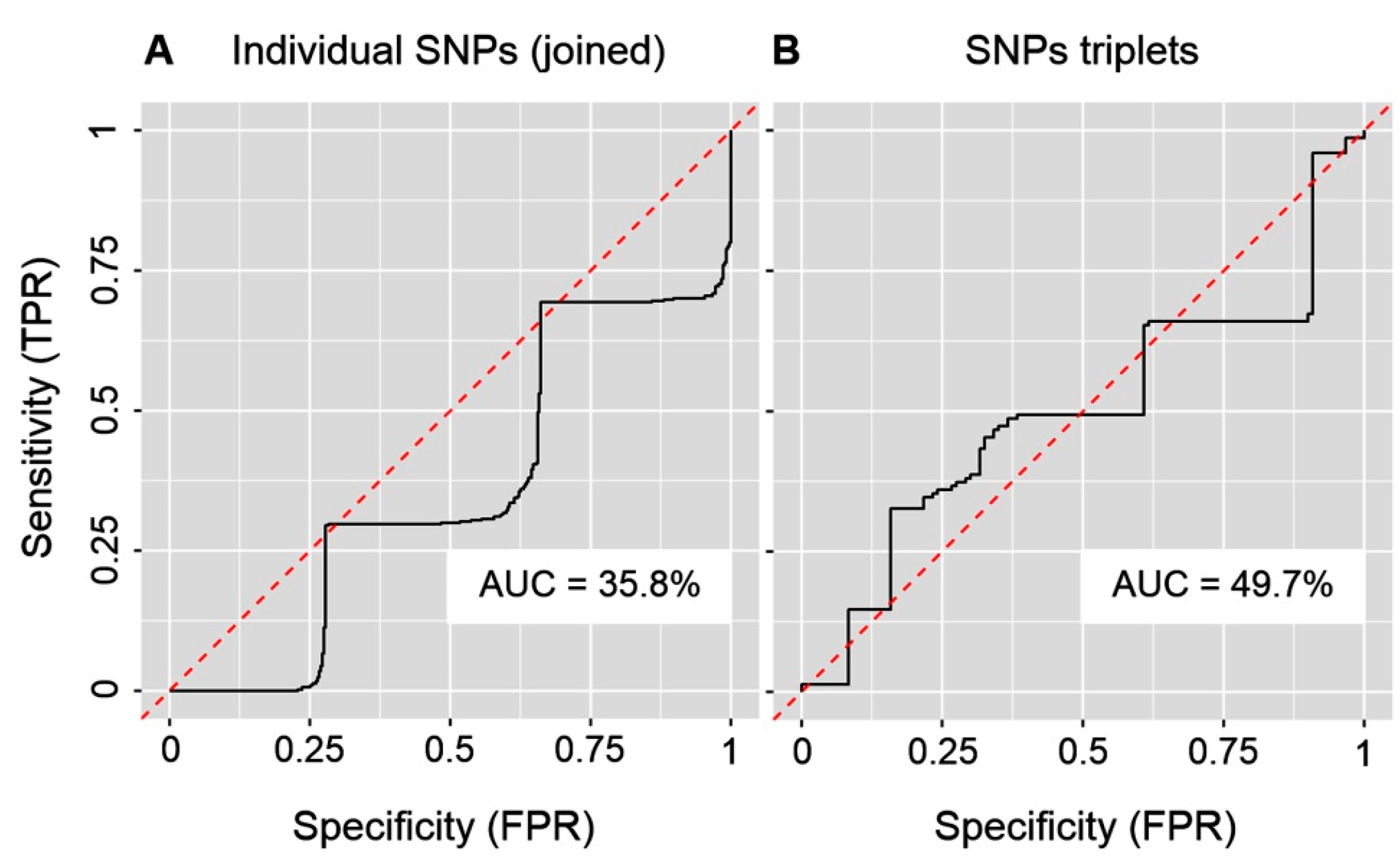
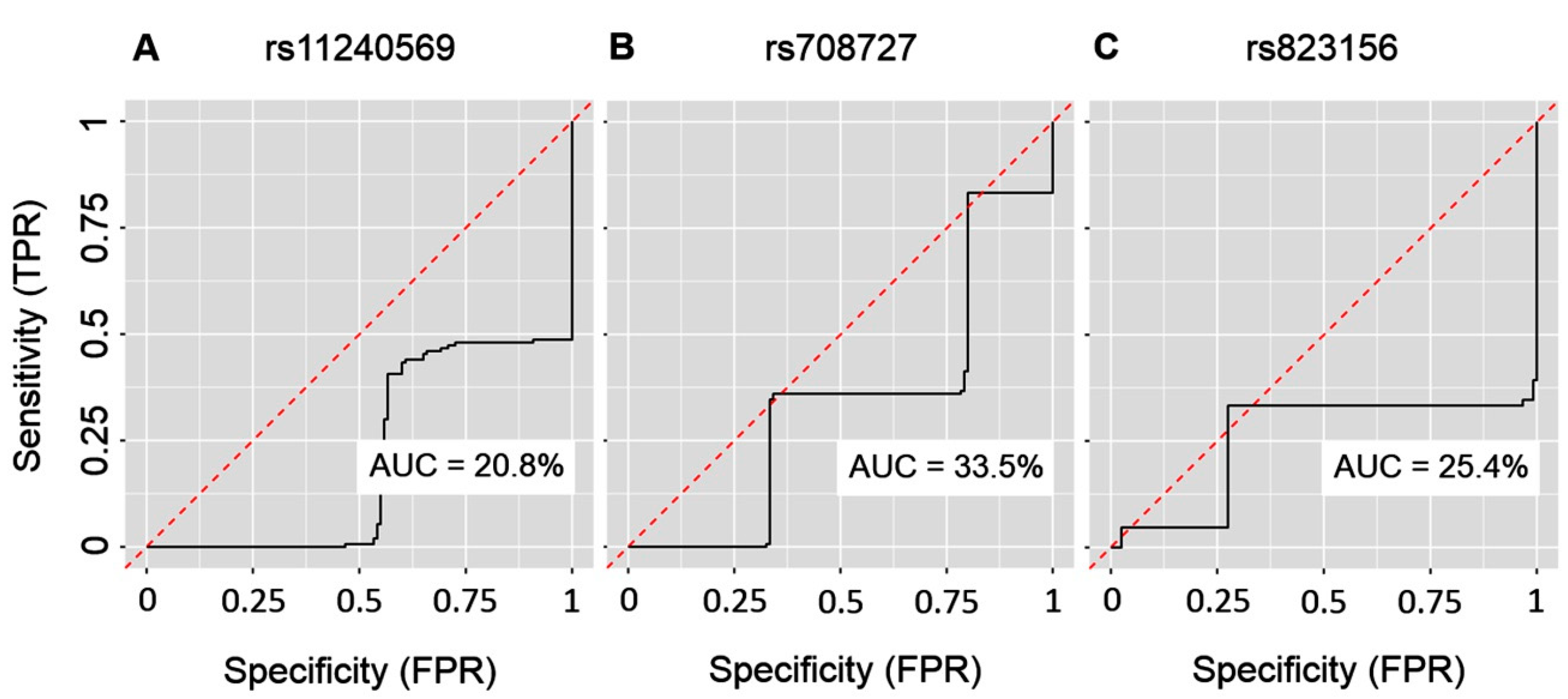
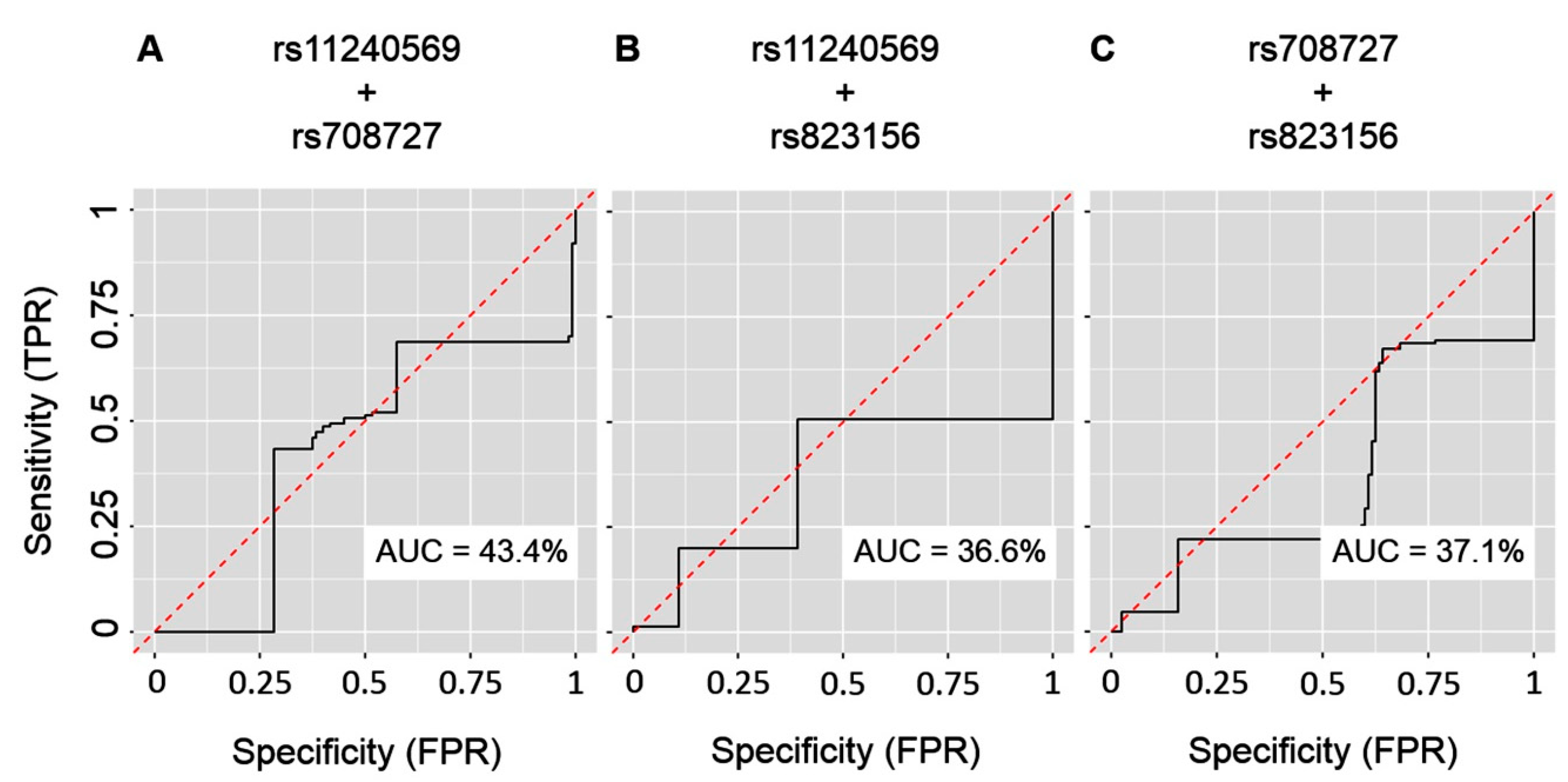
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Participants
4.2. Sample Processing
4.3. Genotyping
4.4. Biostatistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| A1 | SLC41A1 |
| Akt/PKB | Akt/Protein kinase B |
| AUC | Area under curve |
| cAMP | 3’,5’-cyclic adenosine monophosphate |
| CI | Confidence interval |
| DBS | Deep brain stimulation |
| EGF | Epidermal growth factor |
| fq | Frequency |
| HWE | Hardy-Weinberg equilibrium |
| JNK | c-Jun N-terminal kinase |
| MAPFR | Minor allele population frequency range |
| MATPFR | Minor allele total population frequency range |
| ML | Machine-learning |
| MSA | Multiple system atrophy |
| NME | Na+/Mg2+ exchanger |
| NUCKS1 | Nuclear ubiquitous casein and cyclin-dependent kinases substrate |
| OR | Odds ratio |
| OSSE | Online sample size estimator |
| PARK19 | PD-associated locus 16 |
| PD | Parkinson’s disease |
| Pde3b | Phosphodiesterase 3b |
| PDGF | Platelet-derived growth factor |
| PI3K | Phosphoinositide 3-kinase (phosphatidylinositol 3-kinase) |
| PKA | Protein kinase A |
| PM20D1 | Peptidase M20 domain containing 1 |
| p-val | p value |
| RAB7L1 | Ras-related protein Rab-7L1 |
| RF | Random Forest |
| ROC | Receiver operating characteristic |
| RTK | Receptor tyrosine kinases |
| SLC41A1 | Solute carrier family 41, member A1 |
| SLC45A3 | Solute carrier family 45, member A3 (prostate cancer-associated protein 6/prostein) |
| SNP | Single nucleotide polymorphism |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motor Complications of PD | |
| Early | Late |
| Difficult turning in bed | Motor fluctuations |
| Frozen shoulder Stiffness, numbness or pain in limb Micrographia | Choreiformic dyskinesia Gait freezing Falls |
| Difficulty with fine finger movements Tremor of hand, jaw, foot Decreased facial expression Decreased arm swing, dragging a leg Soft voice | |
| Non-motor complications of PD | |
| Early | Late |
| Constipation | Dysphagia |
| REM sleep behaviour disorder | Neuropsychiatric symptoms |
| Depression | Autonomic disturbances |
| Olfaction impairment | Seborrheic dermatitis |
| PD Cohort | C Cohort | Arcsine Transformation * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | N | I | II | N | X2 | df | p-val | d | N | sl | pw |
| AA | AA | 0 | AA | AA | 0 | |||||||
| AA | AG | 0 | AA | AG | 0 | |||||||
| AA | GG | 13 | AA | GG | 11 | 0.00# | 1 | 1.00 | 0.02 | 25493 | 0.5 | 0.8 |
| AG | AA | 0 | AG | AA | 0 | |||||||
| AG | AG | 35 | AG | AG | 37 | 1.55 | 1 | 0.21 | 0.17 | 274 | 0.5 | 0.8 |
| AG | GG | 29 | AG | GG | 15 | 1.81 | 1 | 0.18 | 0.19 | 223 | 0.5 | 0.8 |
| GG | AA | 25 | GG | AA | 24 | 0.30 | 1 | 0.58 | 0.09 | 1056 | 0.5 | 0.8 |
| GG | AG | 36 | GG | AG | 19 | 2.26 | 1 | 0.13 | 0.21 | 186 | 0.5 | 0.8 |
| GG | GG | 12 | GG | GG | 14 | 0.65 | 1 | 0.42 | 0.12 | 513 | 0.5 | 0.8 |
| I | III | I | III | |||||||||
| AA | AA | 11 | AA | AA | 11 | 0.11 | 1 | 0.75 | 0.07 | 1762 | 0.5 | 0.8 |
| AA | AG | 2 | AA | AG | 0 | |||||||
| AA | GG | 0 | AA | GG | 0 | |||||||
| AG | AA | 43 | AG | AA | 42 | 0.96 | 1 | 0.33 | 0.14 | 424 | 0.5 | 0.8 |
| AG | AG | 21 | AG | AG | 10 | 1.59 | 1 | 0.21 | 0.18 | 239 | 0.5 | 0.8 |
| AG | GG | 0 | AG | GG | 0 | |||||||
| GG | AA | 46 | GG | AA | 34 | 0.08 | 1 | 0.78 | 0.05 | 2997 | 0.5 | 0.8 |
| GG | AG | 20 | GG | AG | 20 | 0.35 | 1 | 0.55 | 0.09 | 898 | 0.5 | 0.8 |
| GG | GG | 7 | GG | GG | 3 | 0.38 | 1 | 0.54 | 0.12 | 564 | 0.5 | 0.8 |
| II | III | II | III | |||||||||
| AA | AA | 25 | AA | AA | 24 | 0.30 | 1 | 0.58 | 0.09 | 1055 | 0.5 | 0.8 |
| AA | AG | 0 | AA | AG | 0 | |||||||
| AA | GG | 0 | AA | GG | 0 | |||||||
| AG | AA | 54 | AG | AA | 42 | 2 × 10−3 | 1 | 0.97 | 0.02 | 17971 | 0.5 | 0.8 |
| AG | AG | 17 | AG | AG | 14 | 0.00 ## | 1 | 1.00 | 0.01 | 71890 | 0.5 | 0.8 |
| AG | GG | 0 | AG | GG | 0 | |||||||
| GG | AA | 21 | GG | AA | 21 | 0.38 | 1 | 0.54 | 0.10 | 847 | 0.5 | 0.8 |
| GG | AG | 26 | GG | AG | 16 | 0.54 | 1 | 0.46 | 0.11 | 634 | 0.5 | 0.8 |
| GG | GG | 7 | GG | GG | 3 | 0.38 | 1 | 0.54 | 0.12 | 564 | 0.5 | 0.8 |


References
- Mhyre, T.R.; Boyd, J.T.; Hamill, R.W.; Maguire-Zeiss, K.A. Parkinson’s disease. Subcell. Biochem. 2012, 65, 389–455. [Google Scholar] [PubMed]
- Lebouvier, T.; Chaumette, T.; Paillusson, S.; Duyckaerts, C.; Bruley des Varannes, S.; Neunlist, M.; Derkinderen, P. The second brain and Parkinson’s disease. Eur. J. Neurosci. 2009, 30, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Romani, A. Regulation of magnesium homeostasis and transport in mammalian cells. Arch. Biochem. Biophys. 2007, 458, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Romani, A.M. Cellular magnesium homeostasis. Arch. Biochem. Biophys. 2011, 512, 1–23. [Google Scholar] [CrossRef]
- Langley, W.F.; Mann, D. Central nervous system magnesium deficiency. Arch. Intern. Med. 1991, 151, 593–596. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, A.E.; Sarlo, G.L.; Holton, K.F. The Role of Magnesium in Neurological Disorders. Nutrition 2018, 10, 730. [Google Scholar] [CrossRef]
- Durlach, J. Neurodegenerative Diseases and Magnesium Status. In Magnesium: Current Status and New Developments; Theophanides, T., Anastassopoulou, J., Eds.; Springer: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Kolisek, M.; Sponder, G.; Mastrototaro, L.; Smorodchenko, A.; Launay, P.; Vormann, J.; Schweigel-Röntgen, M. Substitution p.A350V in Na+/Mg2+ exchanger SLC41A1, potentially associated with Parkinson’s disease, is a gain-of-function mutation. PLoS ONE 2013, 8, e71096. [Google Scholar] [CrossRef]
- Kolisek, M.; Launay, P.; Beck, A.; Sponder, G.; Serafini, N.; Brenkus, M.; Froschauer, E.M.; Martens, H.; Fleig, A.; Schweigel, M. SLC41A1 is a novel mammalian Mg2+ carrier. J. Biol. Chem. 2008, 283, 16235–16247. [Google Scholar] [CrossRef]
- Kolisek, M.; Nestler, A.; Vormann, J.; Schweigel-Röntgen, M. Human gene SLC41A1 encodes for the Na+/Mg2+ exchanger. Am. J. Physiol. Cell. Physiol. 2012, 302, 318–326. [Google Scholar] [CrossRef]
- Goytain, A.; Quamme, G.A. Functional characterization of human SLC41A1, a Mg2+ transporter with similarity to prokaryotic MgtE Mg2+ transporters. Physiol. Genom. 2005, 21, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Kolisek, M.; Sponder, G.; Pilchova, I.; Cibulka, M.; Tatarkova, Z.; Werner, T.; Racay, P. Magnesium Extravaganza: A Critical Compendium of Current Research into Cellular Mg2+ Transporters Other than TRPM6/7. Rev. Physiol. Biochem. Pharm. 2019, 176, 65–105. [Google Scholar]
- Mastrototaro, L.; Tietjen, U.; Sponder, G.; Vormann, J.; Aschenbach, J.R.; Kolisek, M. Insulin Modulates the Na+/Mg2+ Exchanger SLC41A1 and Influences Mg2+ Efflux from Intracellular Stores in Transgenic HEK293 Cells. J. Nutr. 2015, 145, 2440–2447. [Google Scholar] [CrossRef] [PubMed]
- Schweigel-Röntgen, M.; Kolisek, M. SLC41 transporters—Molecular identification and functional role. Curr. Top. Membr. 2014, 73, 383–410. [Google Scholar] [PubMed]
- Sponder, G.; Abdulhanan, N.; Fröhlich, N.; Mastrototaro, L.; Aschenbach, J.R.; Röntgen, M.; Pilchova, I.; Cibulka, M.; Racay, P.; Kolisek, M. Overexpression of Na+/Mg2+ exchanger SLC41A1 attenuates pro-survival signaling. Oncotarget 2017, 9, 5084–5104. [Google Scholar] [PubMed]
- Timmons, S.; Coakley, M.F.; Molonay, A.M.; O’Neill, C. Akt signal transduction dysfunction in Parkinson’s disease. Neurosci. Lett. 2009, 467, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, S.J.; Rosas-Hernandez, H.; Cuevas, E.; Lantz, S.M.; Barger, S.W.; Sarkar, S.; Paule, M.G.; Ali, S.F.; Imam, S.Z. Protein Kinases and Parkinson’s Disease. Int. J. Mol. Sci. 2016, 17, 1585. [Google Scholar] [CrossRef]
- Pihlstrøm, L.; Rengmark, A.; Bjørnarå, K.A.; Dizdar, N.; Fardell, C.; Forsgren, L.; Holmberg, B.; Larsen, J.P.; Linder, J.; Nissbrandt, H.; et al. Fine mapping and resequencing of the PARK16 locus in Parkinson’s disease. J. Hum. Genet. 2015, 60, 357–362. [Google Scholar] [CrossRef]
- Tucci, A.; Nalls, M.A.; Houlden, H.; Revesz, T.; Singleton, A.B.; Wood, N.W.; Hardy, J.; Paisán-Ruiz, C. Genetic variability at the PARK16 locus. Eur. J. Hum. Genet. 2010, 18, 1356–1359. [Google Scholar] [CrossRef]
- Kolisek, M.; Montezano, A.C.; Sponder, G.; Anagnostopoulou, A.; Vormann, J.; Touyz, R.M.; Aschenbach, J.R. PARK7/DJ-1 dysregulation by oxidative stress leads to magnesium deficiency: Implications in degenerative and chronic diseases. Clin. Sci. (Lond.) 2015, 129, 1143–1150. [Google Scholar] [CrossRef]
- Lin, C.H.; Wu, Y.R.; Chen, W.L.; Wang, H.C.; Lee, C.M.; Lee-Chen, G.J.; Chen, C.M. Variant R244H in Na+/Mg2+ exchanger SLC41A1 in Taiwanese Parkinson’s disease is associated with loss of Mg2+ efflux function. Parkinsonism Relat. Disord. 2014, 20, 600–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, L.; Li, N.N.; Yu, W.J.; Sun, X.Y.; Peng, R. Genetic analysis of SLC41A1 in Chinese Parkinson’s disease patients. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2015, 168, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Madadi, F.; Khaniani, M.S.; Shandiz, E.E.; Ayromlou, H.; Najmi, S.; Emamalizadeh, B.; Taghavi, S.; Jamshidi, J.; Tafakhori, A.; Shahidi, G.A.; et al. Genetic Analysis of the ZNF512B, SLC41A1, and ALDH2 Polymorphisms in Parkinson’s Disease in the Iranian Population. Genet. Test Mol. Biomark. 2016, 20, 629–632. [Google Scholar] [CrossRef]
- Sanchez-Mut, J.V.; Heyn, H.; Silva, B.A.; Dixsaut, L.; Garcia-Esparcia, P.; Vidal, E.; Sayols, S.; Glauser, L.; Monteagudo-Sánchez, A.; Perez-Tur, J.; et al. PM20D1 is a quantitative trait locus associated with Alzheimer’s disease. Nat. Med. 2018, 24, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Simón-Sánchez, J.; Schulte, C.; Bras, J.M.; Sharma, M.; Gibbs, J.R.; Berg, D.; Paisan-Ruiz, C.; Lichtner, P.; Scholz, S.W.; Hernandez, D.G.; et al. Genome-wide association study reveals genetic risk underlying Parkinson’s disease. Nat. Genet. 2009, 41, 1308–1312. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Kwok, H.H.; Tan, L.C.; Zhao, W.T.; Prakash, K.M.; Au, W.L.; Pavanni, R.; Ng, Y.Y.; Satake, W.; Zhao, Y.; et al. Analysis of GWAS-linked loci in Parkinson disease reaffirms PARK16 as a susceptibility locus. Neurology 2010, 75, 508–512. [Google Scholar] [CrossRef]
- Chang, X.L.; Mao, X.Y.; Li, H.H.; Zhang, J.H.; Li, N.N.; Burgunder, J.M.; Peng, R.; Tan, E.K. Association of GWAS loci with PD in China. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2011, 156, 334–339. [Google Scholar] [CrossRef]
- Yan, Y.P.; Mo, X.Y.; Tian, J.; Zhao, G.H.; Yin, X.Z.; Jin, F.Y.; Zhang, B.R. An association between the PARK16 locus and Parkinson’s disease in a cohort from eastern China. Parkinsonism Relat. Disord. 2011, 17, 737–739. [Google Scholar] [CrossRef]
- Miyake, Y.; Tanaka, K.; Fukushima, W.; Kiyohara, C.; Sasaki, S.; Tsuboi, Y.; Oeda, T.; Shimada, H.; Kawamura, N.; Sakae, N.; et al. PARK16 polymorphisms, interaction with smoking, and sporadic Parkinson’s disease in Japan. J. Neurol. Sci. 2016, 362, 47–52. [Google Scholar] [CrossRef]
- Chung, S.J.; Jung, Y.; Hong, M.; Kim, M.J.; You, S.; Kim, Y.J.; Kim, J.; Song, K. Alzheimer’s disease and Parkinson’s disease genome-wide association study top hits and risk of Parkinson’s disease in Korean population. Neurobiol. Aging 2013, 34, 2695.e1–7. [Google Scholar] [CrossRef]
- Mata, I.F.; Yearout, D.; Alvarez, V.; Coto, E.; de Mena, L.; Ribacoba, R.; Lorenzo-Betancor, O.; Samaranch, L.; Pastor, P.; Cervantes, S.; et al. Replication of MAPT and SNCA, but not PARK16-18, as susceptibility genes for Parkinson’s disease. Mov. Disord. 2011, 26, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Gopalai, A.A.; Ahmad-Annuar, A.; Li, H.H.; Zhao, Y.; Lim, S.Y.; Tan, A.H.; Lim, T.T.; Eow, G.B.; Santhi, P.; Shanthi, V.; et al. PARK16 is associated with PD in the Malaysian population. Am. J. Med. Gene B Neuropsychiatr. Genet. 2016, 171, 839–847. [Google Scholar] [CrossRef]
- Petrejcikova, E.; Carnogurska, J.; Hronska, D.; Bernasovska, J.; Boronova, I.; Gabrikova, D.; Bozikova, A.; Macekova, S. Y-SNP analysis versus Y-haplogroup predictor in the Slovak population. Anthr. Anz. 2014, 71, 275–285. [Google Scholar] [CrossRef]
- Reeve, A.; Simcox, E.; Turnbull, D. Ageing and Parkinson’s disease: Why is advancing age the biggest risk factor? Ageing Res. Rev. 2014, 14, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Moisan, F.; Kab, S.; Mohamed, F.; Canonico, M.; Le Guern, M.; Quintin, C.; Carcaillon, L.; Nicolau, J.; Duport, N.; Singh-Manoux, A.; et al. Parkinson disease male-to-female ratios increase with age: French nationwide study and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2016, 87, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Deo, R.C. Machine Learning in Medicine. Circulation 2015, 132, 1920–1930. [Google Scholar] [CrossRef] [PubMed]
- Rajkomar, A.; Dean, J.; Kohane, I. Machine learning in medicine. N. Engl. J. Med. 2019, 380, 1347–1358. [Google Scholar] [CrossRef] [PubMed]
- Oyanagi, K.; Hashimoto, T. Magnesium in Parkinson’s disease: An update in clinical and basic aspects. In Magnesium in the Central Nervous System; Vink, R., Nechifor, M., Eds.; University of Adelaide Press: Adelaide, Australia, 2011. [Google Scholar]
- Uitti, R.J.; Rajput, A.H.; Rozdilsky, B.; Bickis, M.; Wollin, T.; Yuen, W.K. Regional metal concentrations in Parkinson’s disease, other chronic neurological diseases, and control brains. Can. J. Neurol. Sci. 1989, 16, 310–314. [Google Scholar] [CrossRef]
- Barbiroli, B.; Martinelli, P.; Patuelli, A.; Lodi, R.; Iotti, S.; Cortelli, P.; Montagna, P. Phosphorus magnetic resonance spectroscopy in multiple system atrophy and Parkinson’s disease. Mov. Disord. 1999, 14, 430–435. [Google Scholar] [CrossRef]
- Bocca, B.; Alimonti, A.; Senofonte, O.; Pino, A.; Violante, N.; Petrucci, F.; Sancesario, G.; Forte, G. Metal changes in CSF and peripheral compartments of parkinsonian patients. J. Neurol. Sci. 2006, 248, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Bali, V.; Bebok, Z. Decoding Mechanisms by which Silent Codon Changes Influence Protein Biogenesis and Function. Int. J. Biochem. Cell Biol. 2015, 64, 58–74. [Google Scholar] [CrossRef] [PubMed]
- Mandt, T.; Song, Y.; Scharenberg, A.M.; Sahni, J. SLC41A1 Mg2+ transport is regulated via Mg2+-dependent endosomal recycling through its N-terminal cytoplasmic domain. Biochem. J. 2011, 439, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Dong, L.; Huang, X.; Zheng, S.; Qiu, P.; Lan, F. Associations of rs823128, rs1572931, and rs823156 polymorphisms with reduced Parkinson’s disease risks. Neuroreport 2017, 28, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Visel, A.; Rubin, E.M.; Pennacchio, L.A. Genomic views of distant-acting enhancers. Nature 2009, 461, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ruan, X.; Auerbach, R.K.; Sandhu, K.S.; Zheng, M.; Wang, P.; Poh, H.M.; Goh, Y.; Lim, J.; Zhang, J.; et al. Extensive Promoter-Centered Chromatin Interactions Provide a Topological Basis for Transcription Regulation. Cell 2012, 148, 84–98. [Google Scholar] [CrossRef]
- Yan, Y.; Tian, J.; Mo, X.; Zhao, G.; Yin, X.; Pu, J.; Zhang, B. Genetic Variants in the RAB7L1 and SLC41A1 Genes of the PARK16 Locus in Chinese Parkinson’s Disease Patients. Int. J. Neurosci. 2011, 121, 632–636. [Google Scholar] [CrossRef]
- Lin, M.; Lucas, H.C., Jr.; Shmueli, G. Research commentary—Too big to fail: Large samples and the p-value problem. Inf. Syst. Res. 2013, 24, 906–917. [Google Scholar] [CrossRef]
- Lo, A.; Chernoff, H.; Zheng, T.; Lo, S.H. Why significant variables aren’t automatically good predictors. Proc. Natl. Acad. Sci. USA 2015, 112, 13892–13897. [Google Scholar] [CrossRef]
- Breiman, L. Statistical modeling: The two cultures (with comments and a rejoinder by the author). Stat. Sci. 2001, 16, 199–231. [Google Scholar] [CrossRef]
- Devroye, L.; Györfi, L.; Lugosi, G. Vapnik-Chervonenkis Theory. In A Probabilistic Theory of Pattern Recognition; Devroye, L., Györfi, L., Lugosi, G., Eds.; Stochastic Modelling and Applied Probability; Springer: New York, NY, USA, 1996. [Google Scholar]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. 2015, 30, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, M.M.; Yahr, M.D. Parkinsonism: Onset, progression and mortality. Neurology 1967, 17, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Gaunt, T.R.; Day, I.N. Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am. J. Epidemiol. 2009, 169, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Schrider, D.R.; Kern, A.D. Supervised Machine Learning for Population Genetics: A New Paradigm. Trends Genet. 2018, 34, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.A.; Zhang, W.; Balding, D.J. Approximate Bayesian computation in population genetics. Genetics 2002, 162, 2025–2035. [Google Scholar]
- Rizek, P.; Kumar, N.; Jog, M.S. An update on the diagnosis and treatment of Parkinson disease. Cmaj 2016, 188, 1157–1165. [Google Scholar] [CrossRef] [PubMed]




| SNP | Cohort | Alelle | Count | fq(%) | Genotype | Count | fq(%) |
|---|---|---|---|---|---|---|---|
| rs11240569 | PD | G | 210 | 70 | GG | 73 | 48 |
| A | 90 | 30 | AG | 64 | 43 | ||
| AA | 13 | 9 | |||||
| C | G | 166 | 69 | GG | 57 | 48 | |
| A | 74 | 31 | AG | 52 | 43 | ||
| AA | 11 | 9 | |||||
| rs708727 | PD | G | 179 | 60 | GG | 54 | 36 |
| A | 121 | 40 | AG | 71 | 47 | ||
| AA | 25 | 17 | |||||
| C | G | 136 | 57 | GG | 40 | 33 | |
| A | 104 | 43 | AG | 56 | 47 | ||
| AA | 24 | 20 | |||||
| rs823156 | PD | A | 243 | 81 | AA | 100 | 67 |
| G | 57 | 19 | AG | 43 | 29 | ||
| GG | 7 | 5 | |||||
| C | A | 204 | 85 | AA | 87 | 73 | |
| G | 36 | 15 | AG | 30 | 25 | ||
| GG | 3 | 2 |
| SNP | rs11240569 (G > A) Cohort | rs708727 (G > A) Cohort | rs823156 (A > G) Cohort | |||
|---|---|---|---|---|---|---|
| PD N (O/E) | C N (O/E) | PD N (O/E) | C N (O/E) | PD N (O/E) | C N (O/E) | |
| GG (com.) | 73/73.5 | 57/57.4 | ||||
| AG | 64/63 | 52/51.2 | ||||
| AA (rar.) | 13/13.5 | 11/11.4 | ||||
| X2 | 0.04 | 0.03 | ||||
| p-val | 0.85 | 0.86 | ||||
| GG (com.) | 54/53.4 | 40/38.53 | ||||
| AG | 71/72.2 | 56/58.93 | ||||
| AA (rar.) | 25/24.4 | 24/22.54 | ||||
| X2 | 0.04 | 0.30 | ||||
| p-val | 0.84 | 0.59 | ||||
| AA (com.) | 100/98.42 | 87/86.7 | ||||
| AG | 43/46.16 | 30/30.6 | ||||
| GG (rar.) | 7/5.42 | 3/2.7 | ||||
| X2 | 0.71 | 0.05 | ||||
| p-val | 0.40 | 0.83 | ||||
| 95% CI | 95% CI | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SNP | MA | OR | ll | ul | p-val | genotype | OR | ll | uL | p-val |
| rs11240569 (G > A) | A | 1.04 | 0.71 | 1.53 | 0.85 | AA | 1.08 | 0.41 | 2.84 | 1.00 |
| AG | 1.04 | 0.61 | 1.78 | 0.90 | ||||||
| rs708727 (G > A) | A | 1.13 | 0.79 | 1.62 | 0.48 | AA | 1.29 | 0.61 | 2.75 | 0.48 |
| AG | 1.06 | 0.60 | 1.89 | 0.89 | ||||||
| rs823156 (A > G) | G | 1.33 | 0.82 | 2.17 | 0.25 | GG | 2.02 | 0.45 | 12.5 | 0.35 |
| AG | 1.62 | 0.34 | 10.5 | 0.73 | ||||||
| PD Cohort | C Cohort | Arcsine Transformation * | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | N | I | II | III | N | X2 | df | p-val | d | N | sl | pw |
| AA | AA | AA | 0 | AA | AA | AA | 0 | |||||||
| AG | AA | AA | 0 | AG | AA | AA | 0 | |||||||
| GG | AA | AA | 25 | GG | AA | AA | 24 | 0.30 | 1 | 0.58 | 0.09 | 1056 | 0.05 | 0.8 |
| AA | AG | AA | 0 | AA | AG | AA | 0 | |||||||
| AG | AG | AA | 34 | AG | AG | AA | 36 | 1.51 | 1 | 0.22 | 0.17 | 282 | 0.05 | 0.8 |
| GG | AG | AA | 20 | GG | AG | AA | 6 | 4.41 | 1 | 0.04 | 0.30 | 89 | 0.05 | 0.8 |
| AA | GG | AA | 11 | AA | GG | AA | 11 | 0.11 | 1 | 0.75 | 0.07 | 1762 | 0.05 | 0.8 |
| AG | GG | AA | 9 | AG | GG | AA | 6 | 0.01 | 1 | 0.93 | 0.04 | 4071 | 0.05 | 0.8 |
| GG | GG | AA | 1 | GG | GG | AA | 4 | 1.35 | 1 | 0.25 | 0.20 | 189 | 0.05 | 0.8 |
| AA | AA | AG | 0 | AA | AA | AG | 0 | |||||||
| AG | AA | AG | 0 | AG | AA | AG | 0 | |||||||
| GG | AA | AG | 0 | GG | AA | AG | 0 | |||||||
| AA | AG | AG | 0 | AA | AG | AG | 0 | |||||||
| AG | AG | AG | 1 | AG | AG | AG | 1 | 0.00 | 1 | 1 | 0.02 | 20968 | 0.05 | 0.8 |
| GG | AG | AG | 16 | GG | AG | AG | 13 | 0.00 | 1 | 1 | 0.01 | 271094 | 0.05 | 0.8 |
| AA | GG | AG | 2 | AA | GG | AG | 0 | |||||||
| AG | GG | AG | 20 | AG | GG | AG | 9 | 1.80 | 1 | 0.18 | 0.19 | 211 | 0.05 | 0.8 |
| GG | GG | AG | 4 | GG | GG | AG | 7 | 1.00 | 1 | 0.32 | 0.16 | 307 | 0.05 | 0.8 |
| AA | AA | GG | 0 | AA | AA | GG | 0 | |||||||
| AG | AA | GG | 0 | AG | AA | GG | 0 | |||||||
| GG | AA | GG | 0 | GG | AA | GG | 0 | |||||||
| AA | AG | GG | 0 | AA | AG | GG | 0 | |||||||
| AG | AG | GG | 0 | AG | AG | GG | 0 | |||||||
| GG | AG | GG | 0 | GG | AG | GG | 0 | |||||||
| AA | GG | GG | 0 | AA | GG | GG | 0 | |||||||
| AG | GG | GG | 0 | AG | GG | GG | 0 | |||||||
| GG | GG | GG | 7 | GG | GG | GG | 3 | 0.38 | 1 | 0.54 | 0.12 | 564 | 0.05 | 0.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cibulka, M.; Brodnanova, M.; Grendar, M.; Grofik, M.; Kurca, E.; Pilchova, I.; Osina, O.; Tatarkova, Z.; Dobrota, D.; Kolisek, M. SNPs rs11240569, rs708727, and rs823156 in SLC41A1 Do Not Discriminate Between Slovak Patients with Idiopathic Parkinson’s Disease and Healthy Controls: Statistics and Machine-Learning Evidence. Int. J. Mol. Sci. 2019, 20, 4688. https://doi.org/10.3390/ijms20194688
Cibulka M, Brodnanova M, Grendar M, Grofik M, Kurca E, Pilchova I, Osina O, Tatarkova Z, Dobrota D, Kolisek M. SNPs rs11240569, rs708727, and rs823156 in SLC41A1 Do Not Discriminate Between Slovak Patients with Idiopathic Parkinson’s Disease and Healthy Controls: Statistics and Machine-Learning Evidence. International Journal of Molecular Sciences. 2019; 20(19):4688. https://doi.org/10.3390/ijms20194688
Chicago/Turabian StyleCibulka, Michal, Maria Brodnanova, Marian Grendar, Milan Grofik, Egon Kurca, Ivana Pilchova, Oto Osina, Zuzana Tatarkova, Dusan Dobrota, and Martin Kolisek. 2019. "SNPs rs11240569, rs708727, and rs823156 in SLC41A1 Do Not Discriminate Between Slovak Patients with Idiopathic Parkinson’s Disease and Healthy Controls: Statistics and Machine-Learning Evidence" International Journal of Molecular Sciences 20, no. 19: 4688. https://doi.org/10.3390/ijms20194688
APA StyleCibulka, M., Brodnanova, M., Grendar, M., Grofik, M., Kurca, E., Pilchova, I., Osina, O., Tatarkova, Z., Dobrota, D., & Kolisek, M. (2019). SNPs rs11240569, rs708727, and rs823156 in SLC41A1 Do Not Discriminate Between Slovak Patients with Idiopathic Parkinson’s Disease and Healthy Controls: Statistics and Machine-Learning Evidence. International Journal of Molecular Sciences, 20(19), 4688. https://doi.org/10.3390/ijms20194688