Specialist and Generalist Fungal Parasites Induce Distinct Biochemical Changes in the Mandible Muscles of Their Host

 ,
, 
Abstract
:1. Introduction
2. Results
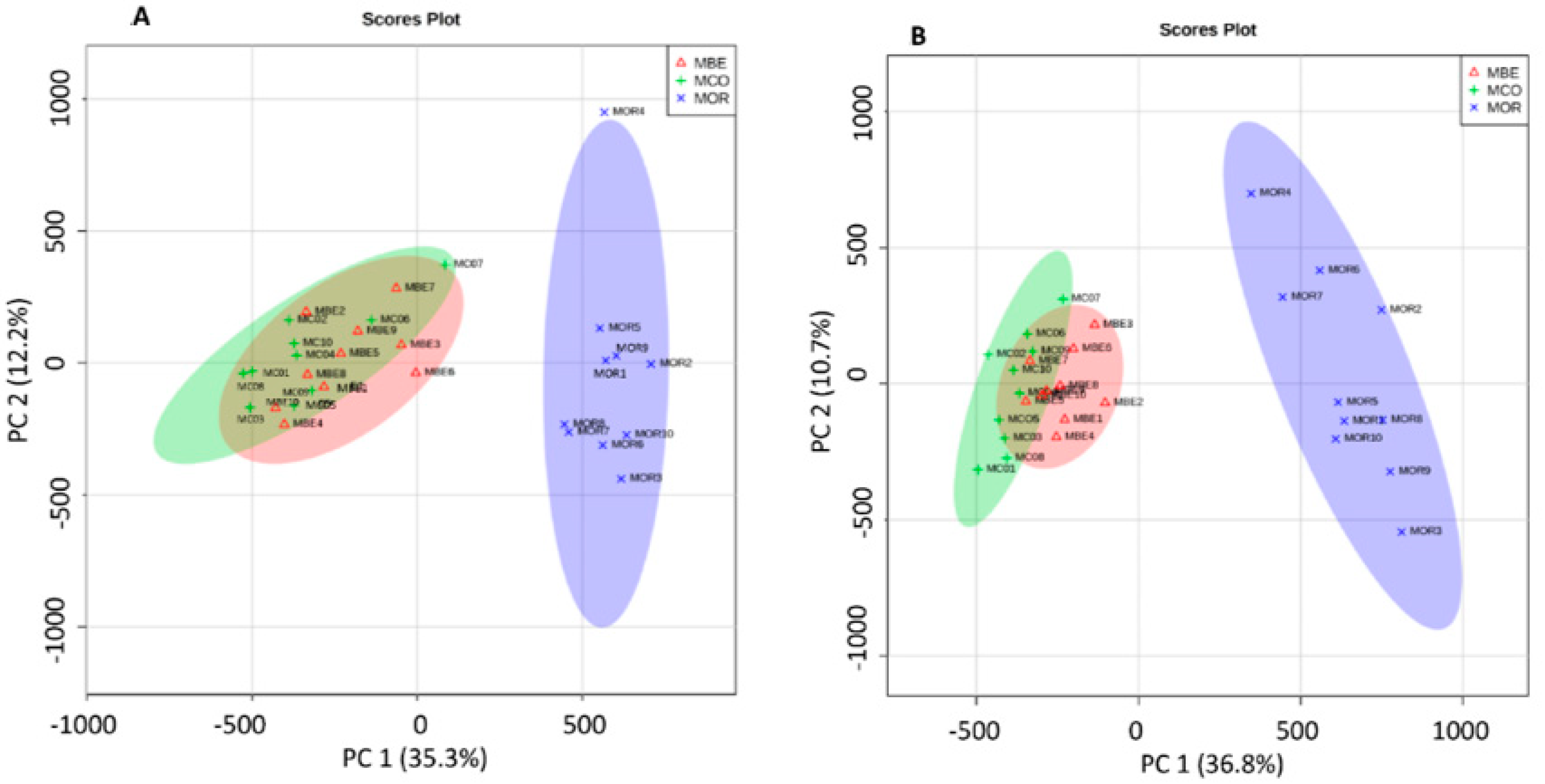
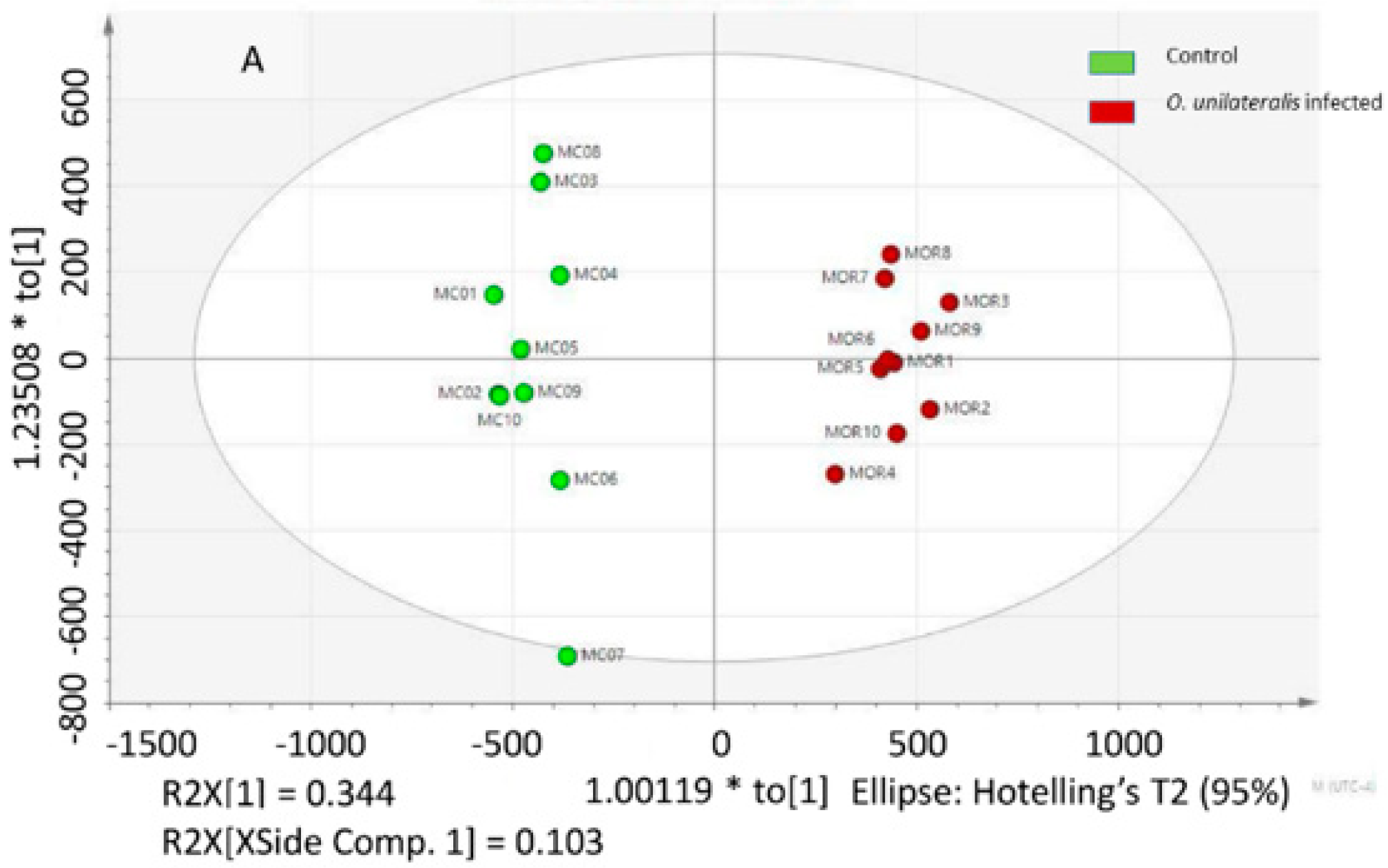
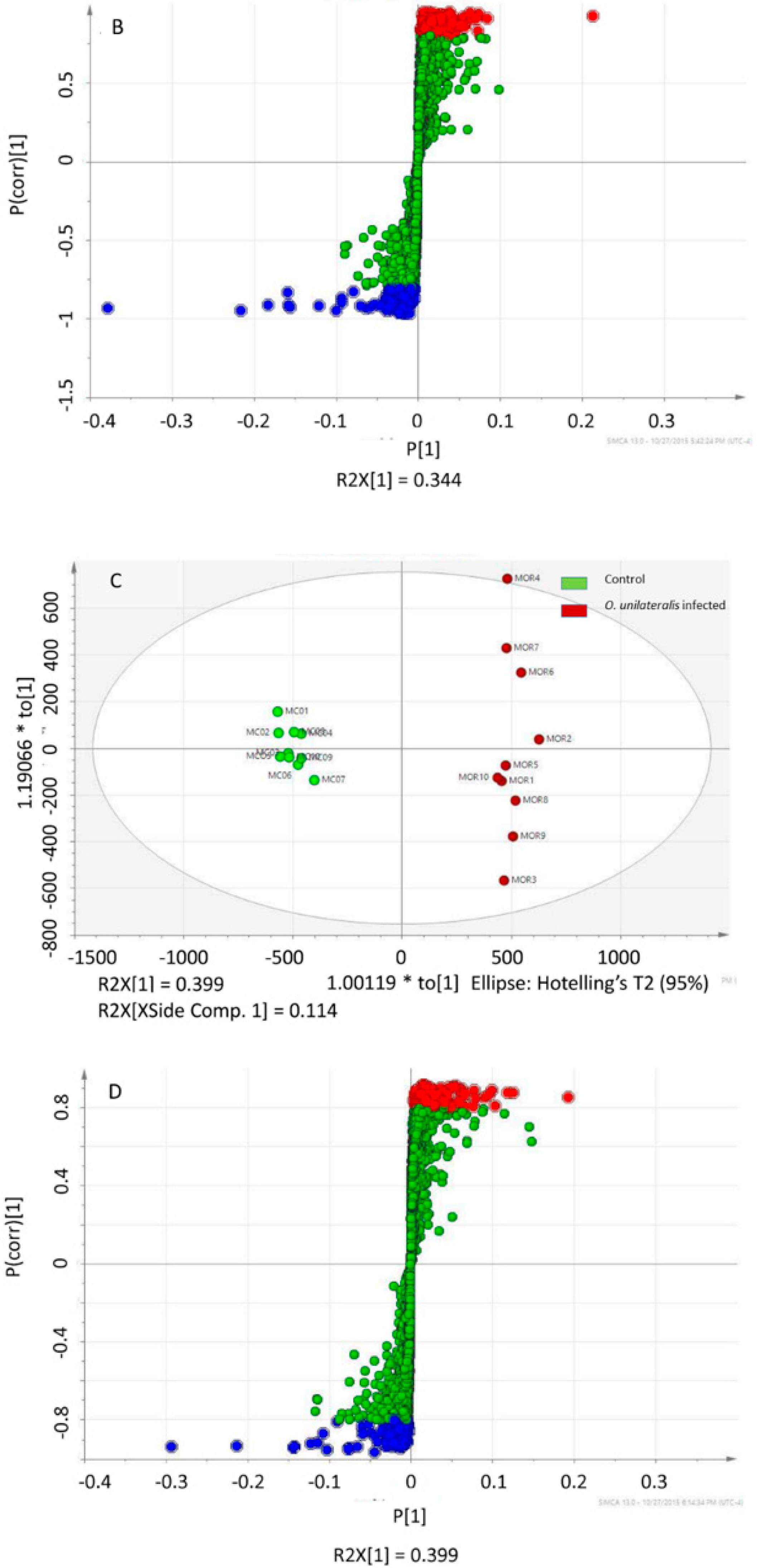
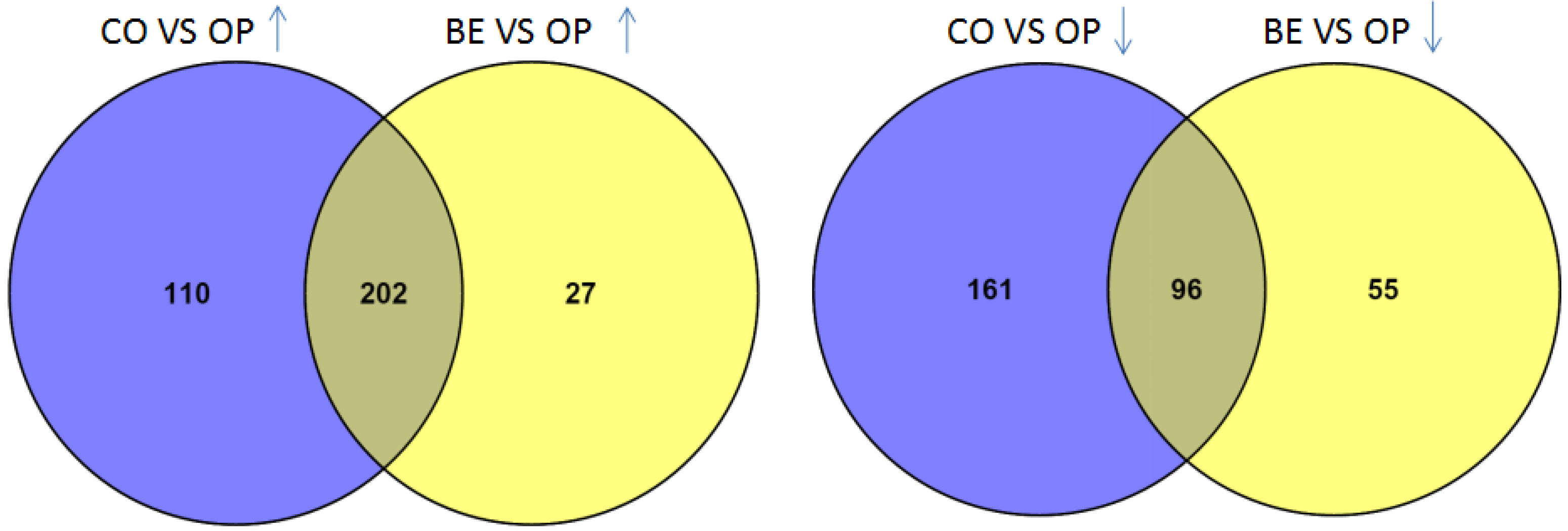
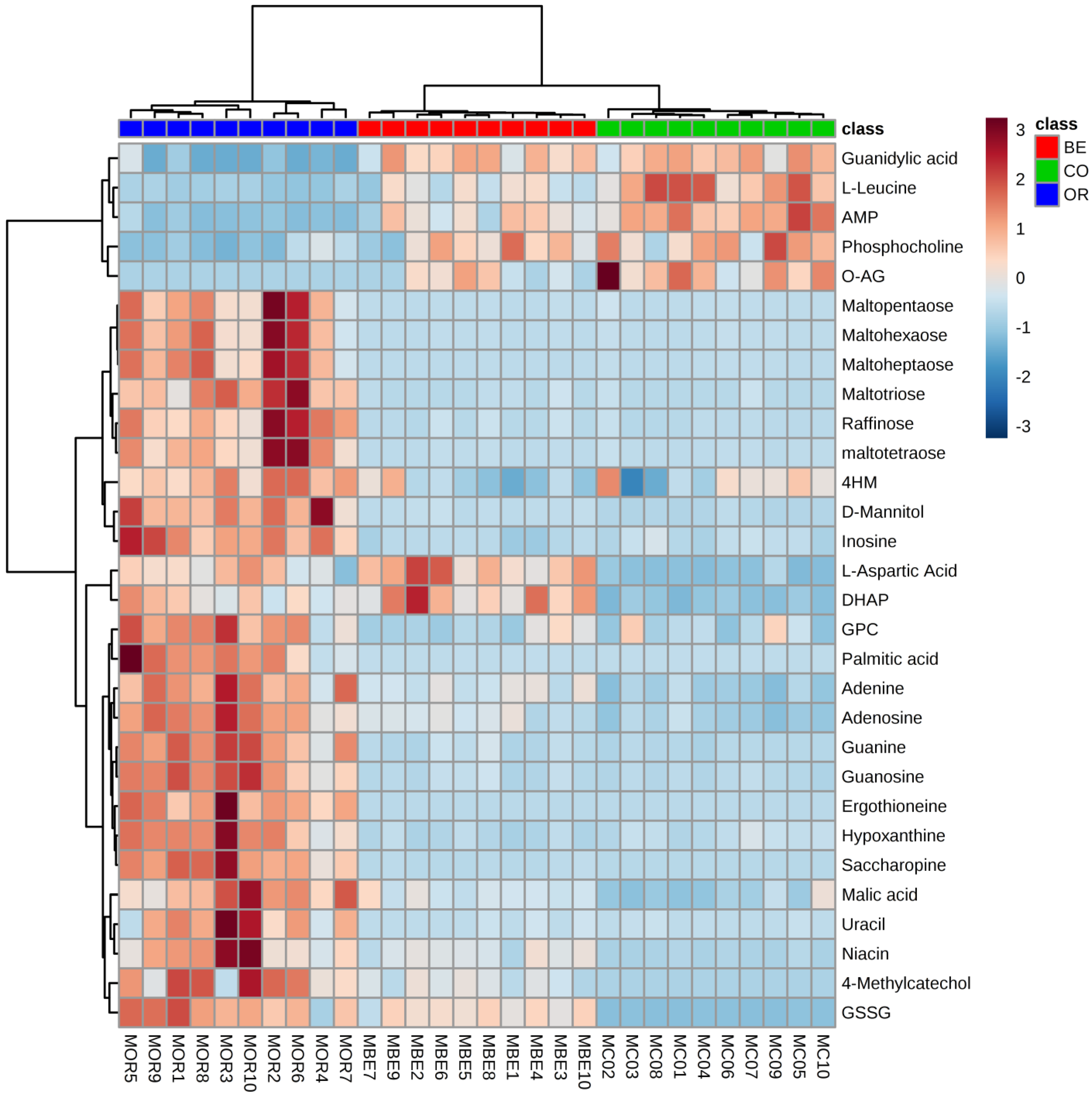
2.1. Difference between the Metabolome of Fungal-Infected Ant Muscles
2.2. Identifying Tissues Metabolites between Behaviorally Manipulated Ants and Controls
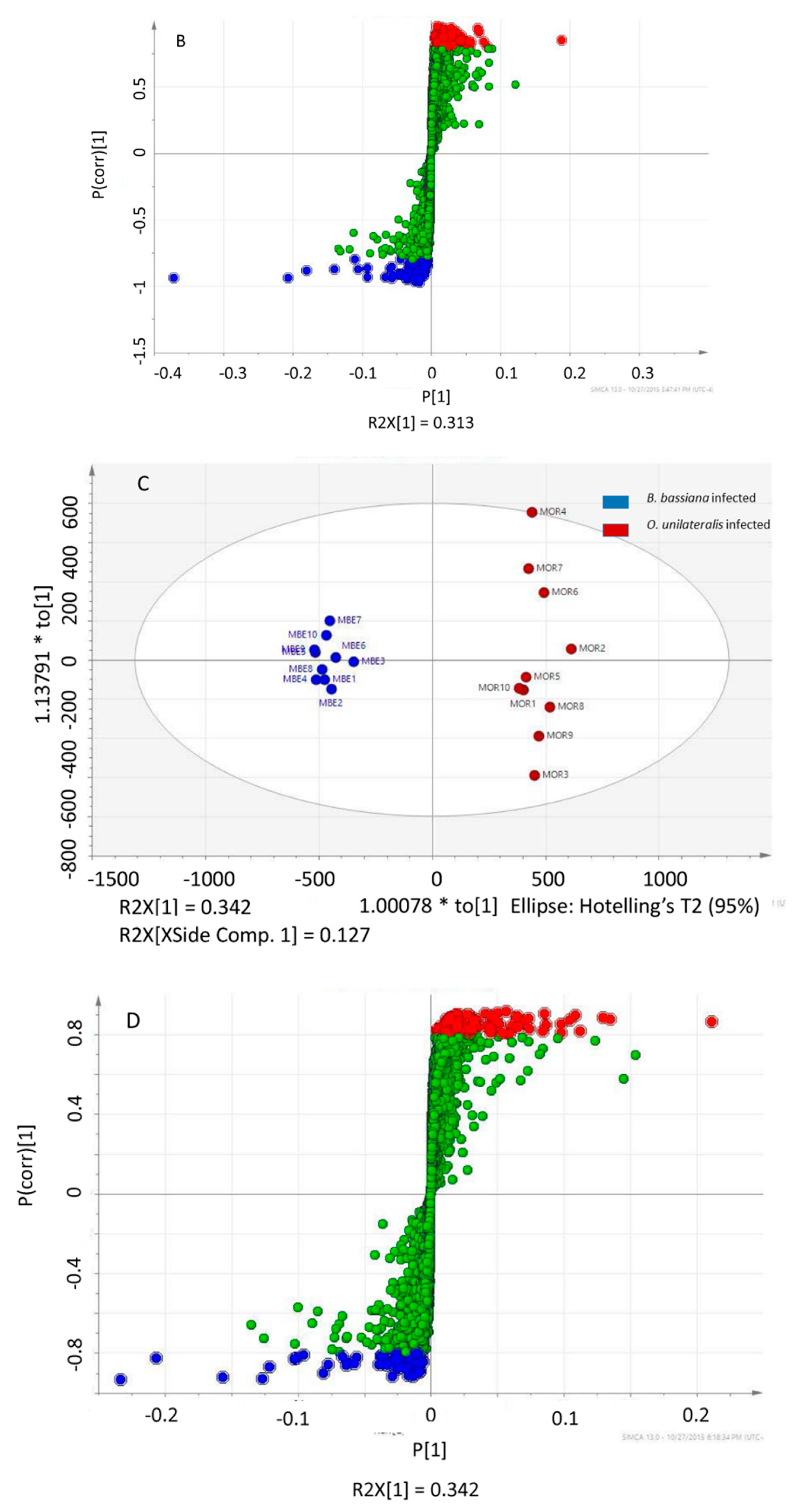
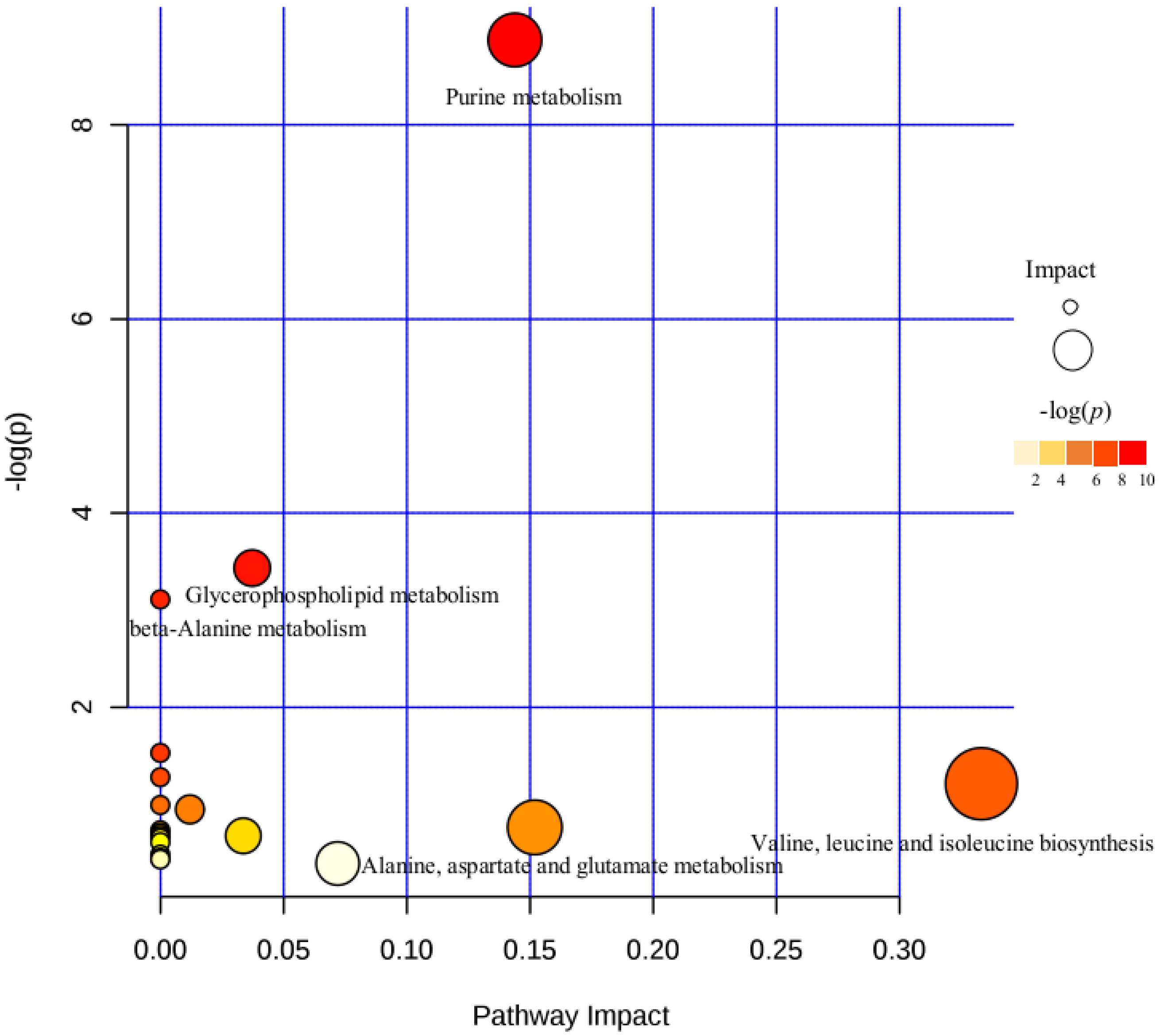
2.3. Different Metabolites in the Comparison between Ophiocordyceps unilateralis s.l. and Beauveria bassiana-Infected Ants
3. Materials and Methods
3.1. Camponotus Castaneus Ant-Rearing and Treatments
3.2. Ophiocordyceps unilateralis s.l. Infection
3.3. Beauveria Bassiana Infection
3.4. Healthy Control Ants
3.5. Metabolite Assay
3.6. Statistics Analysis
3.7. Metabolite Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, J. Parasites and the behavior of animals. Zool. Afr. 2002, 37, 265. [Google Scholar]
- Bekker, C.D.; Ohm, R.A.; Loreto, R.G.; Sebastian, A.; Albert, I.; Merrow, M.; Brachmann, A.; Hughes, D.P. Gene expression during zombie ant biting behavior reflects the complexity underlying fungal parasitic behavioral manipulation. BMC Genom. 2015, 16, 620. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.P.; Araújo, J.P.; Loreto, R.G.; Quevillon, L.; De, B.C.; Evans, H.C. From so simple a beginning: The evolution of behavioral manipulation by fungi. Adv. Genet. 2016, 94, 437. [Google Scholar] [PubMed]
- Loreto, R.G.; Elliot, S.L.; Freitas, M.L.; Pereira, T.M.; Hughes, D.P. Long-term disease dynamics for a specialized parasite of ant societies: A field study. PLoS ONE 2014, 9, e103516. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.B.; Gerritsma, S.; Yusah, K.M.; Mayntz, D.; Hyweljones, N.L.; Billen, J.; Boomsma, J.J.; Hughes, D.P. The life of a dead ant: The expression of an adaptive extended phenotype. Am. Nat. 2009, 174, 424–433. [Google Scholar] [CrossRef]
- Hughes, D.P.; Andersen, S.B.; Hywel-Jones, N.L.; Himaman, W.; Billen, J.; Boomsma, J.J. Behavioral mechanisms and morphological symptoms of zombie ants dying from fungal infection. BMC Ecol. 2011, 11, 13. [Google Scholar] [CrossRef]
- Molnár, I.; Gibson, D.M.; Krasnoff, S.B. Secondary metabolites from entomopathogenic hypocrealean fungi. Cheminform 2010, 27, 1241–1275. [Google Scholar] [CrossRef]
- Kloehn, J.; Blume, M.; Cobbold, S.A.; Saunders, E.C.; Dagley, M.J.; Mcconville, M.J. Using metabolomics to dissect host-parasite interactions. Curr. Opin. Microbiol. 2016, 32, 59–65. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Boguś, M.I. Harman and norharman, metabolites of the entomopathogenic fungus Conidiobolus coronatus (Entomophthorales), affect the serotonin levels and phagocytic activity of hemocytes, insect immunocompetent cells, in Galleria mellonella (Lepidoptera). Cell Biosci. 2019, 9, 29. [Google Scholar] [CrossRef]
- Boyce, G.R.; Gluck-Thaler, E.; Slot, J.C.; Stajich, J.E.; Davis, W.J.; James, T.Y.; Cooley, J.R.; Panaccione, D.G.; Eilenberg, J.; De Fine Licht, H.H.; et al. Psychoactive plant- and mushroom-associated alkaloids from two behavior modifying cicada pathogens. Fungal Ecol. 2019, 41, 147–164. [Google Scholar] [CrossRef]
- Bekker, C.D.; Quevillon, L.E.; Smith, P.B.; Fleming, K.R.; Ghosh, D.; Patterson, A.D.; Hughes, D.P. Species-specific ant brain manipulation by a specialized fungal parasite. BMC Evol. Biol. 2014, 14, 166. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.; Alfred, R.; Götz, S.; Hans, S. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar]
- Warrack, B.M.; Hnatyshyn, S.; Ott, K.H.; Reily, M.D.; Sanders, M.; Zhang, H.; Drexler, D.M. Normalization strategies for metabonomic analysis of urine samples. J. Chromatogr. B 2009, 877, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Wold, S.; Esbensen, K.; Geladi, P. Principal component analysis. Chemom. Intell. Lab. Syst. 1987, 2, 37–52. [Google Scholar] [CrossRef]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemom. 2010, 16, 119–128. [Google Scholar] [CrossRef]
- Anthony, J.C.; Yoshizawa, F.; Anthony, T.G.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr. 2000, 130, 2413–2419. [Google Scholar] [CrossRef]
- Buse, M.G.; Reid, S.S. Leucine. A possible regulator of protein turnover in muscle. J. Clin. Investig. 1975, 56, 1250–1261. [Google Scholar] [CrossRef]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. A high proportion of leucine is required for optimal stimulation of the rate of muscle protein synthesis by essential amino acids in the elderly. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E381–E387. [Google Scholar] [CrossRef] [Green Version]
- Rieu, I.; Balage, M.; Sornet, C.; Debras, E.; Ripes, S.; Rochon-Bonhomme, C.; Pouyet, C.; Grizard, J.; Dardevet, D. Increased availability of leucine with leucine-rich whey proteins improves postprandial muscle protein synthesis in aging rats. Nutrition 2007, 23, 323–331. [Google Scholar] [CrossRef]
- Saha, A.K.; Xu, J.; Ebony, L.; Rosangela, D.; Brandon, A.E.; Kraegen, E.W.; Ruderman, N.B. Downregulation of AMPK accompanies leucine- and glucose-induced increases in protein synthesis and insulin resistance in rat skeletal muscle. Diabetes 2010, 59, 2426–2434. [Google Scholar] [CrossRef]
- Ventrucci, G.; Mello, M.A.R.; Gomes-Marcondes, M.C.C. Leucine-rich diet alters the eukaryotic translation initiation factors expression in skeletal muscle of tumour-bearing rats. BMC Cancer 2007, 7, 42. [Google Scholar] [CrossRef]
- Chulay, J.D.; Haynes, J.D.; Diggs, C.L. Plasmodium falciparum: Assessment of in vitro growth by [3H]hypoxanthine incorporation. Exp. Parasitol. 1983, 55, 138–146. [Google Scholar] [CrossRef]
- Dangoudoubiyam, S.; Zhang, Z.; Howe, D.K.; Hemphill, A. Purine salvage in the apicomplexan Sarcocystis neurona, and generation of hypoxanthine-xanthine-guanine phosphoribosyltransferase-deficient clones for positive-negative selection of transgenic parasites. Parasitology 2014, 141, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Morse, S.A.; Bartenstein, L. Purine metabolism in Neisseria gonorrhoeae: The requirement for hypoxanthine. Can. J. Microbiol. 1980, 26, 13–20. [Google Scholar] [CrossRef]
- Ullman, B.; Carter, D. Hypoxanthine-guanine phosphoribosyltransferase as a therapeutic target in protozoal infections. Infect. Agents Dis. 1995, 4, 29–40. [Google Scholar]
- Saugstad, O.D. Hypoxanthine as a measurement of hypoxia. Pediatric Res. 1975, 9, 158–161. [Google Scholar] [CrossRef]
- Simmonds, R.J.; Coade, S.B.; Harkness, R.A.; Drury, L.; Hytten, F.E. Nucleotide, nucleoside and purine base concentrations in human placentae. Placenta 1982, 3, 29–37. [Google Scholar] [CrossRef]
- Feet, B.A.; Yu, X.Q.; Rootwelt, T.; Oyasaeter, S.; Saugstad, O.D. Effects of hypoxemia and reoxygenation with 21% or 100% oxygen in newborn piglets: Extracellular hypoxanthine in cerebral cortex and femoral muscle. Crit. Care Med. 1997, 25, 1384–1391. [Google Scholar] [CrossRef]
- Phillis, J.W.; O’Regan, M.H.; Walter, G.A. Effects of deoxycoformycin on adenosine, inosine, hypoxanthine, xanthine, and uric acid release from the hypoxemic rat cerebral cortex. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 1988, 8, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Arruda, P.; Kemper, E.L.; Papes, F.; Leite, A. Regulation of lysine catabolism in higher plants. Trends Plant Sci. 2000, 5, 324–330. [Google Scholar] [CrossRef]
- Arruda, P.; Sodek, L.; Silva, W.J. Lysine-ketoglutarate reductase activity in developing maize endosperm. Plant Physiol. 1982, 69, 988–989. [Google Scholar] [CrossRef]
- Serrano, G.C.D.M.; Figueira, T.R.E.S.; Kiyota, E.; Zanata, N.; Arruda, P. Lysine degradation through the saccharopine pathway in bacteria: LKR and SDH in bacteria and its relationship to the plant and animal enzymes. FEBS Lett. 2012, 586, 905–911. [Google Scholar] [CrossRef] [Green Version]
- Higgins, A.D.; Silverstein, J.T.; Engles, J.; Wilson, M.E.; Rexroad, C.E.; Blemings, K.P. Starvation induced alterations in hepatic lysine metabolism in different families of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2005, 31, 33–44. [Google Scholar] [CrossRef]
- Trupin, J.S. Lysine biosynthesis in Neurospora crassa. Fed. Proc. 1963, 22, 243–250. [Google Scholar]
- Liebmann, B.; Mühleisen, T.W.; Müller, M.; Hecht, M.; Weidner, G.; Braun, A.; Brock, M.; Brakhage, A.A. Deletion of the Aspergillus fumigatus lysine biosynthesis gene lysF encoding homoaconitase leads to attenuated virulence in a low-dose mouse infection model of invasive aspergillosis. Arch. Microbiol. 2004, 181, 378–383. [Google Scholar] [CrossRef]
- Fu, X.; Gao, H.; Tian, F.; Gao, J.; Lou, L.; Liang, Y.; Ning, Q.; Luo, X. Mechanistic effects of amino acids and glucose in a novel glutaric aciduria type 1 cell model. PLoS ONE 2014, 9, e110181. [Google Scholar] [CrossRef]
- Higashino, K.; Tsukada, K.; Lieberman, I. Saccharopine, a product of lysine breakdown by mammalian liver. Biochem. Biophys. Res. Commun. 1965, 20, 285–290. [Google Scholar] [CrossRef]
- Johansson, E.; Steffens, J.J.; Lindqvist, Y.; Schneider, G. Crystal structure of saccharopine reductase from Magnaporthe grisea, an enzyme of the α-aminoadipate pathway of lysine biosynthesis. Structure 2000, 8, 1037–1047. [Google Scholar] [CrossRef]
- Jones, E.E.; Broquist, H.P. Saccharopine, an intermediate of the aminoadipic acid pathway of lysine biosynthesis. Ii. Studies in Saccharomyces cereviseae. J. Biol. Chem. 1965, 240, 2531. [Google Scholar]
- Aoyagi, Y.; Sasaki, H.; Sugahara, T. Isolation and Identification of Saccharopine from Lentinus edodes. J. Agric. Chem. Soc. Jpn. 1978, 42, 1941–1942. [Google Scholar] [CrossRef]
- Hallen, A.; Jamie, J.F.; Cooper, A.J.L. Lysine metabolism in mammalian brain: An update on the importance of recent discoveries. Amino Acids 2013, 45, 1249–1272. [Google Scholar] [CrossRef]
- Hutzler, J.; Dancis, J. Conversion of lysine to saccharopine by human tissues. Biochim. Biophys. Acta 1968, 158, 62–69. [Google Scholar] [CrossRef]
- Hassan, Y.; Layla, K.; Osta, M.A. The mosquito melanization response is implicated in defense against the entomopathogenic fungus Beauveria bassiana. PLoS Pathog. 2012, 8, e1003029. [Google Scholar]
- Thierry, D.; Richard, B.; Elisabeth, G.; Pascale, C. Amino acid changes during sunflower infection by the necrotrophic fungus Botrytis cinerea. Plant Signal. Behav. 2009, 4, 859–861. [Google Scholar]
- Ulrich, H.; Fouad, O.; Franz-Josef, M.; Hermann, B. The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiol. Lett. 2006, 254, 258–267. [Google Scholar]
- Mchunu, N.P.; Permaul, K.; Alam, M.; Singh, S. Carbon utilization profile of a thermophilic fungus, Thermomyces lanuginosus using phenotypic microarray. Adv. Biosci. Biotechnol. 2013, 4, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Genghof, D.S.; Vandamme, O. Biosynthesis of ergothioneine and hercynine by mycobacteria. J. Bacteriol. 1964, 87, 852–862. [Google Scholar]
- Dirk, G.; Stephanie, H.; Stefan, G.; Andreas, G.; Andreas, L.; Reinhard, B.; Norma, J.; Andrea, R.; Edgar, S.M. Discovery of the ergothioneine transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [Green Version]
- Melville, D.B.; Horner, W.H.; Otken, C.C.; Ludwig, M.L. Studies on the origin of ergothioneine in animals. J. Biol. Chem. 1955, 213, 61–68. [Google Scholar]
- Cumming, B.M.; Chinta, K.C.; Reddy, V.P.; Steyn, A.J. Role of ergothioneine in microbial physiology and pathogenesis. Antioxid. Redox Signal. 2018, 28, 431–444. [Google Scholar] [CrossRef]
- Askari, A.; Melville, D.B. The reaction sequence in ergothioneine biosynthesis: Hercynine as an intermediate. J. Biol. Chem. 1962, 237, 1615–1618. [Google Scholar]
- Seebeck, F.P. In vitro reconstitution of Mycobacterial ergothioneine biosynthesis. J. Am. Chem. Soc. 2010, 132, 6632–6633. [Google Scholar] [CrossRef] [PubMed]
- Servillo, L.; Castaldo, D.; Casale, R.; D’Onofrio, N.; Giovane, A.; Cautela, D.; Balestrieri, M.L. An uncommon redox behavior sheds light on the cellular antioxidant properties of ergothioneine. Free Radic. Biol. Med. 2015, 79, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.Z.; Mao, L.; Frei, B. Ergothioneine prevents copper-induced oxidative damage to DNA and protein by forming a redox-inactive ergothioneine-copper complex. Chem. Res. Toxicol. 2011, 24, 30. [Google Scholar] [CrossRef] [PubMed]
- Akanmu, D.; Cecchini, R.; Aruoma, O.I.; Halliwell, B. The antioxidant action of ergothioneine. Arch. Biochem. Biophys. 1991, 288, 10–16. [Google Scholar] [CrossRef]
- Franzoni, F.; Colognato, R.; Galetta, F.; Laurenza, I.; Barsotti, M.; Stefano, R.D.; Bocchetti, R.; Regoli, F.; Carpi, A.; Balbarini, A.; et al. An in vitro study on the free radical scavenging capacity of ergothioneine: Comparison with reduced glutathione, uric acid and trolox. Biomed. Pharmacother. 2006, 60, 453–457. [Google Scholar] [CrossRef]
- Chaudière, J.; Ferrari-Iliou, R. Intracellular antioxidants: From chemical to biochemical mechanisms. Food Chem. Toxicol. 1999, 37, 949–962. [Google Scholar] [CrossRef]
- Bello, M.H.; Viviana, B.P.; Dexter, M.; Lynn, E. The Neurospora crassa mutant NcΔEgt−1 identifies an ergothioneine biosynthetic gene and demonstrates that ergothioneine enhances conidial survival and protects against peroxide toxicity during conidial germination. Fungal Genet. Biol. 2012, 49, 160–172. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | p(corr) | m/z_Retention Time (Sample) | Sigma Cas | m/z_Retention Time (Standard) | Ionization |
|---|---|---|---|---|---|
| Guanidylic acid | −0.892268 | 364.0638_1.16 | No | No | Positive |
| L-Leucine | −0.862599 | 132.1050_1.98 | L8000 | 132.1019_1.84 | Positive |
| Phosphocholine | −0.820645 | 184.0729_0.98 | P0378 | 184.0717_0.95 | Positive |
| O-Arachidonoyl Glycidol (O-AG) | −0.809626 | 361.2721_13.63 | No | No | Positive |
| Phosphocholine | −0.82065 | 184.0733_0.98 | A1752 | 184.0717_0.95 | Positive |
| Niacin | 0.807232 | 124.0395_1.45 | No | No | Positive |
| Uracil | 0.808126 | 113.0350_1.61 | U0750 | 113.0357_1.45 | Positive |
| Palmitic acid | 0.814864 | 279.2302_13.84 | No | No | Positive |
| Glycerophosphocholine | 0.852189 | 258.1107_0.99 | 111–02−4 | 258.1095_0.94 | Positive |
| Maltotriose | 0.863712 | 505.1750_1.07 | No | No | Positive |
| Ergothioneine | 0.915323 | 230.0957_1.03 | E7521 | 230.0957_1.01 | Positive |
| Guanosine | 0.928987 | 284.0967_1.06 | G6753 | 284.1002_1.15 | Positive |
| Saccharopine | 0.933777 | 299.1281_1.00 | S1634 | 299.1280_0.92 | Positive |
| L-Glutathione (Oxidized) (GSSG) | 0.941697 | 613.1566_1.08 | 150568 | 613.1733_1.03 | Positive |
| Adenosine | 0.944343 | 268.1037_1.06 | A9251 | 268.1029_1.02 | Positive |
| Adenine | 0.95219 | 136.0630_1.08 | A8626 | 136.0643_1.12 | Positive |
| Guanine | 0.961177 | 152.0574_1.08 | G11950 | 152.0558_1.10 | Positive |
| 4-Methylcatechol | 0.793135 | 123.0454_3.90 | 452–86−8 | 123.0457_5.83(Isomer) | Negative |
| L-Aspartic Acid | 0.804651 | 132.0312_1.18 | A9256 | 132.0417_1.06 | Negative |
| Maltohexaose | 0.804924 | 989.3323_1.26 | M9153 | 989.3382_1.03(Isomer) | Negative |
| Maltotetraose | 0.809164 | 665.2176_1.23 | No | No | Negative |
| Malic acid | 0.832735 | 133.0150_1.42 | No | No | Negative |
| Dihydroxy acetone phosphate (DHAP) | 0.845409 | 168.9923_1.31 | D7137 | 168.9923_1.10 | Negative |
| Raffinose | 0.851847 | 503.1649_1.22 | R0514 | 503.1649_0.97(Isomer) | Negative |
| GSSG | 0.866878 | 611.1549_1.25 | G4626 | 611.1553_1.28 | Negative |
| D-Mannitol | 0.871107 | 181.0722_1.20 | 443907(EMD) | 181.0729_0.97 | Negative |
| Guanosine | 0.883834 | 282.0838_1.26 | G6752 | 282.0849_1.52 | Negative |
| Inosine | 0.898411 | 267.0734_1.89 | I4125 | 267.0762_1.55 | Negative |
| Compounds | p(corr) | m/z_Retention Time (Sample) | Sigma Cas | m/z_Retention Time (Standard) | Ionization |
|---|---|---|---|---|---|
| Guanine | −0.915064 | 152.0570_1.09 | G11950 | 152.0560_1.04 | Positive |
| Ergothioneine | −0.908576 | 230.0964_1.01 | E7521 | 230.0957_1.01 | Positive |
| Guanosine | −0.88987 | 284.0967_1.06 | G6753 | 284.1002_1.15 | Positive |
| Hypoxanthine | −0.885579 | 137.0459_1.06 | H9377 | 137.0461_1.03 | Positive |
| Adenosine | −0.850788 | 268.1037_1.06 | 116833 | 268.1029_1.02 | Positive |
| Adenine | −0.842341 | 136.0621_1.06 | A8626 | 136.0643_1.12 | Positive |
| 4-Hydroxycinnamoylmethane(4-HM) | −0.805273 | 163.0735_9.68 | No | No | Positive |
| Glycemphosphocholine | −0.803298 | 258.1095_0.99 | 258.1095_0.94 | Positive | |
| AMP | 0.800485 | 348.0707_1.08 | A1752 | 348.0688_1.03 | Positive |
| Guanidylic acid | 0.883806 | 364.0638_1.16 | No | No | Positive |
| Maltopentaose | 0.806347 | 827.2799_1.16 | No | No | Negative |
| Maltoteraose | 0.814304 | 665.2176_1.23 | No | No | Negative |
| Maltoheptaose | 0.837664 | 1151.3911_1.16 | no | No | Negative |
| Raffinose | 0.851338 | 503.1664_1.22 | R0514 | 503.1664_0.97(Isomer) | Negative |
| D-Mannitol | 0.855712 | 181.0722_1.20 | 443907(EMD) | 181.0729_0.97 | Negative |
| Inosine | 0.904759 | 267.0734_1.89 | I4125 | 267.0762_1.55 | Negative |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, S.; Loreto, R.; Smith, P.; Patterson, A.; Hughes, D.; Wang, L. Specialist and Generalist Fungal Parasites Induce Distinct Biochemical Changes in the Mandible Muscles of Their Host. Int. J. Mol. Sci. 2019, 20, 4589. https://doi.org/10.3390/ijms20184589
Zheng S, Loreto R, Smith P, Patterson A, Hughes D, Wang L. Specialist and Generalist Fungal Parasites Induce Distinct Biochemical Changes in the Mandible Muscles of Their Host. International Journal of Molecular Sciences. 2019; 20(18):4589. https://doi.org/10.3390/ijms20184589
Chicago/Turabian StyleZheng, Shanshan, Raquel Loreto, Philip Smith, Andrew Patterson, David Hughes, and Liande Wang. 2019. "Specialist and Generalist Fungal Parasites Induce Distinct Biochemical Changes in the Mandible Muscles of Their Host" International Journal of Molecular Sciences 20, no. 18: 4589. https://doi.org/10.3390/ijms20184589
APA StyleZheng, S., Loreto, R., Smith, P., Patterson, A., Hughes, D., & Wang, L. (2019). Specialist and Generalist Fungal Parasites Induce Distinct Biochemical Changes in the Mandible Muscles of Their Host. International Journal of Molecular Sciences, 20(18), 4589. https://doi.org/10.3390/ijms20184589