Transcriptome Analysis of Sogatella furcifera (Homoptera: Delphacidae) in Response to Sulfoxaflor and Functional Verification of Resistance-Related P450 Genes

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Toxicity of Sulfoxaflor in S. furcifera
2.2. Synergism Experiment
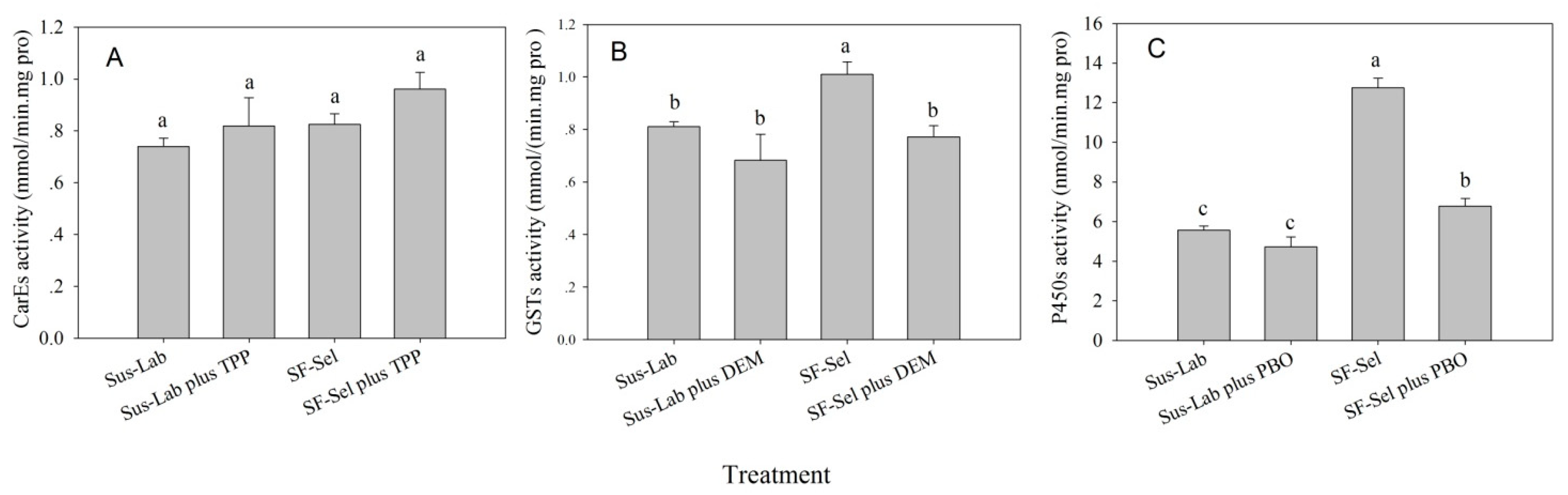
2.3. Detoxification Enzyme Activity
2.4. Illumina Sequencing and Read Assembly
2.5. Transcriptome Data Splicing
2.6. Transcriptome Annotation
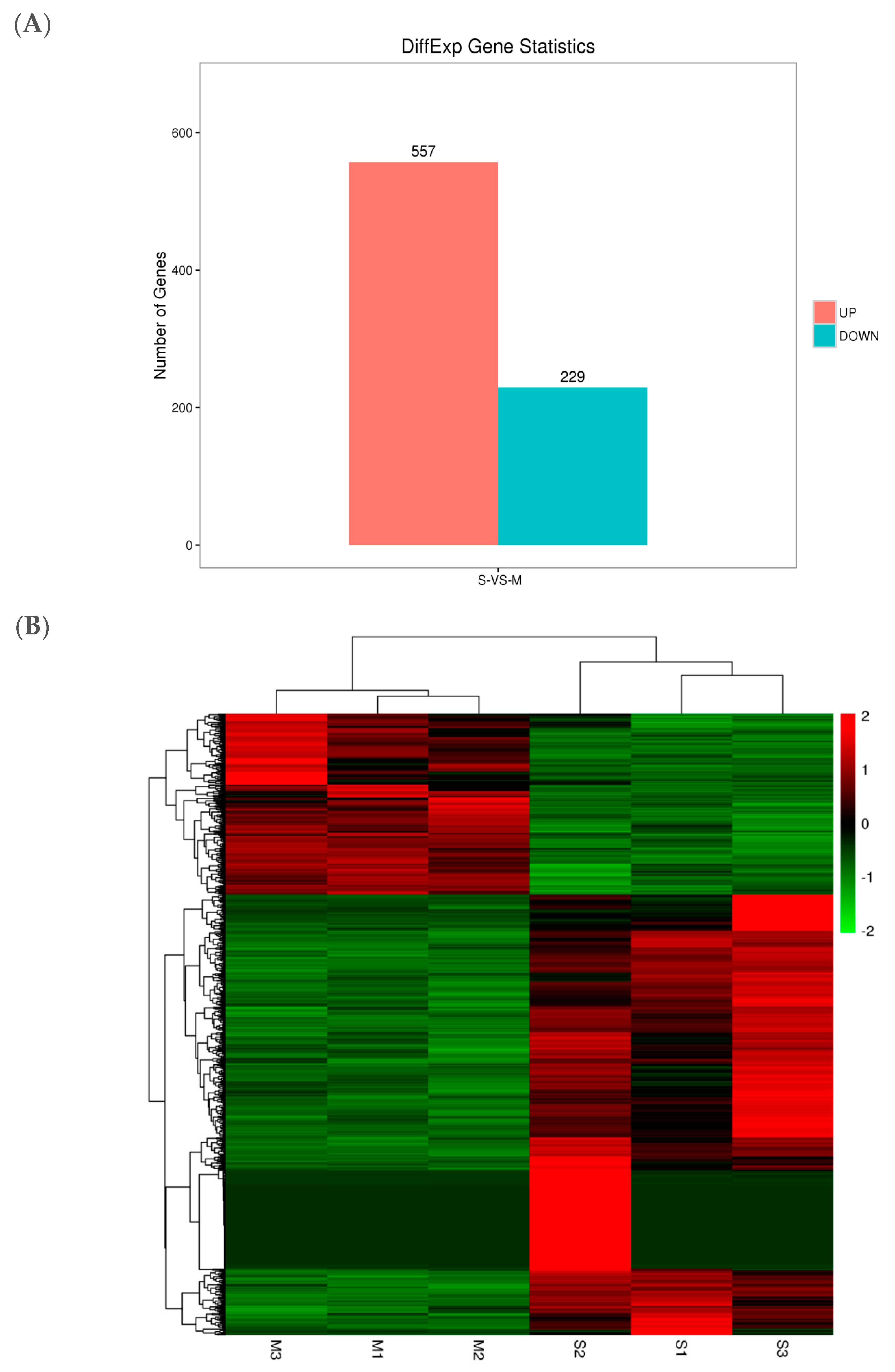
2.7. Analysis of Gene Expression
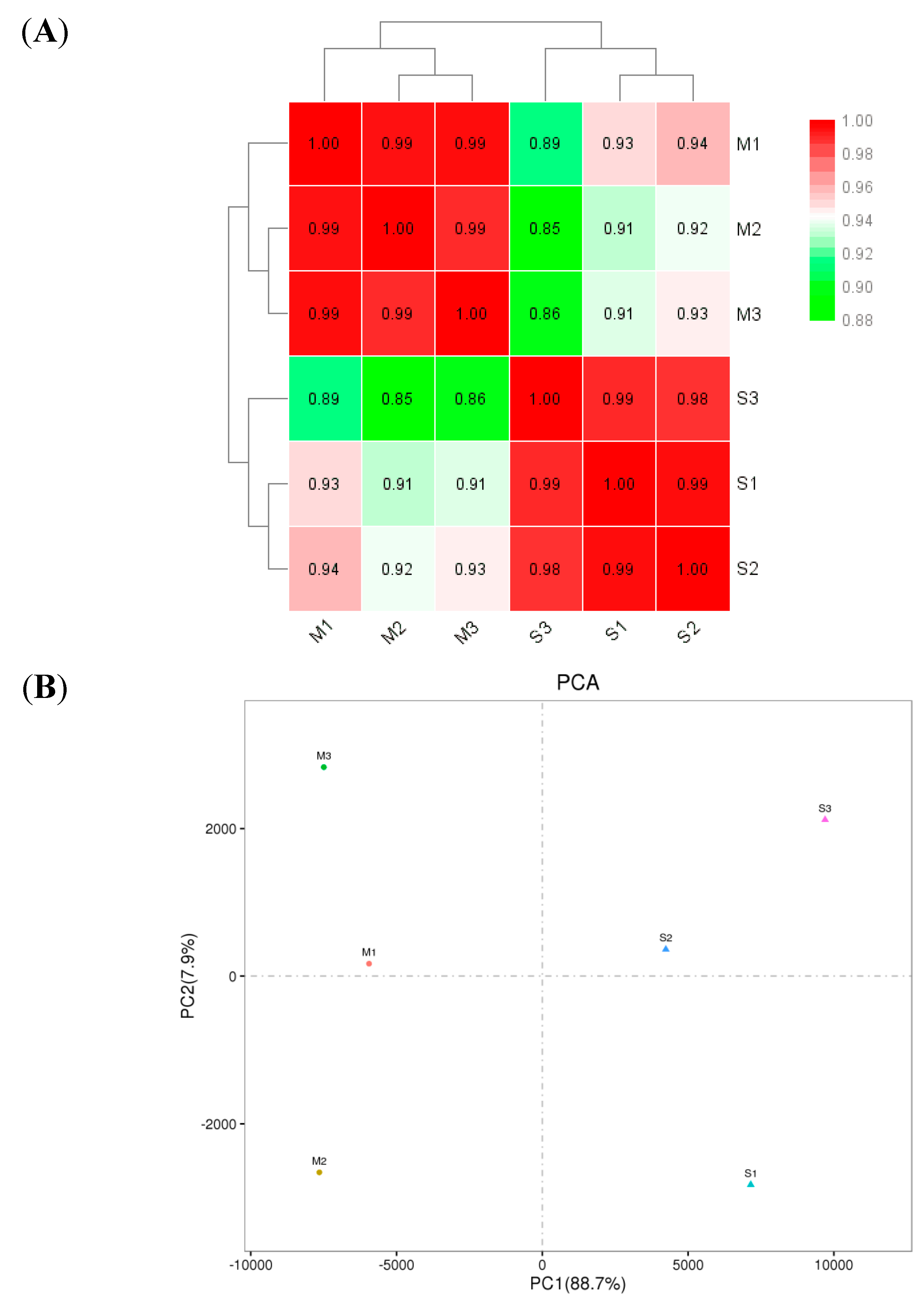
2.8. Cluster Analysis of DEGs
2.8.1. GO Enrich
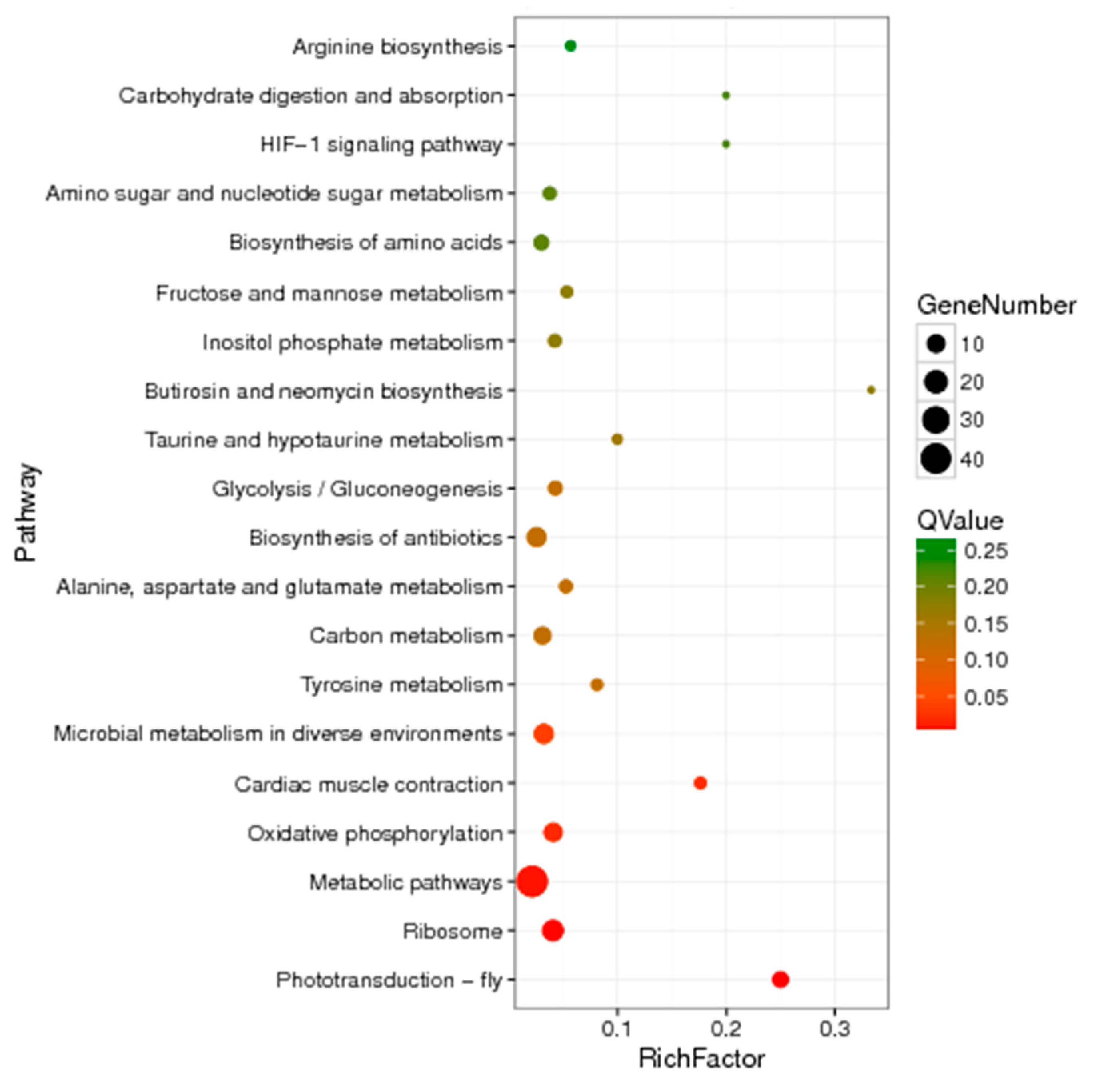
2.8.2. Enrichment of DEGs in the KEGG Database
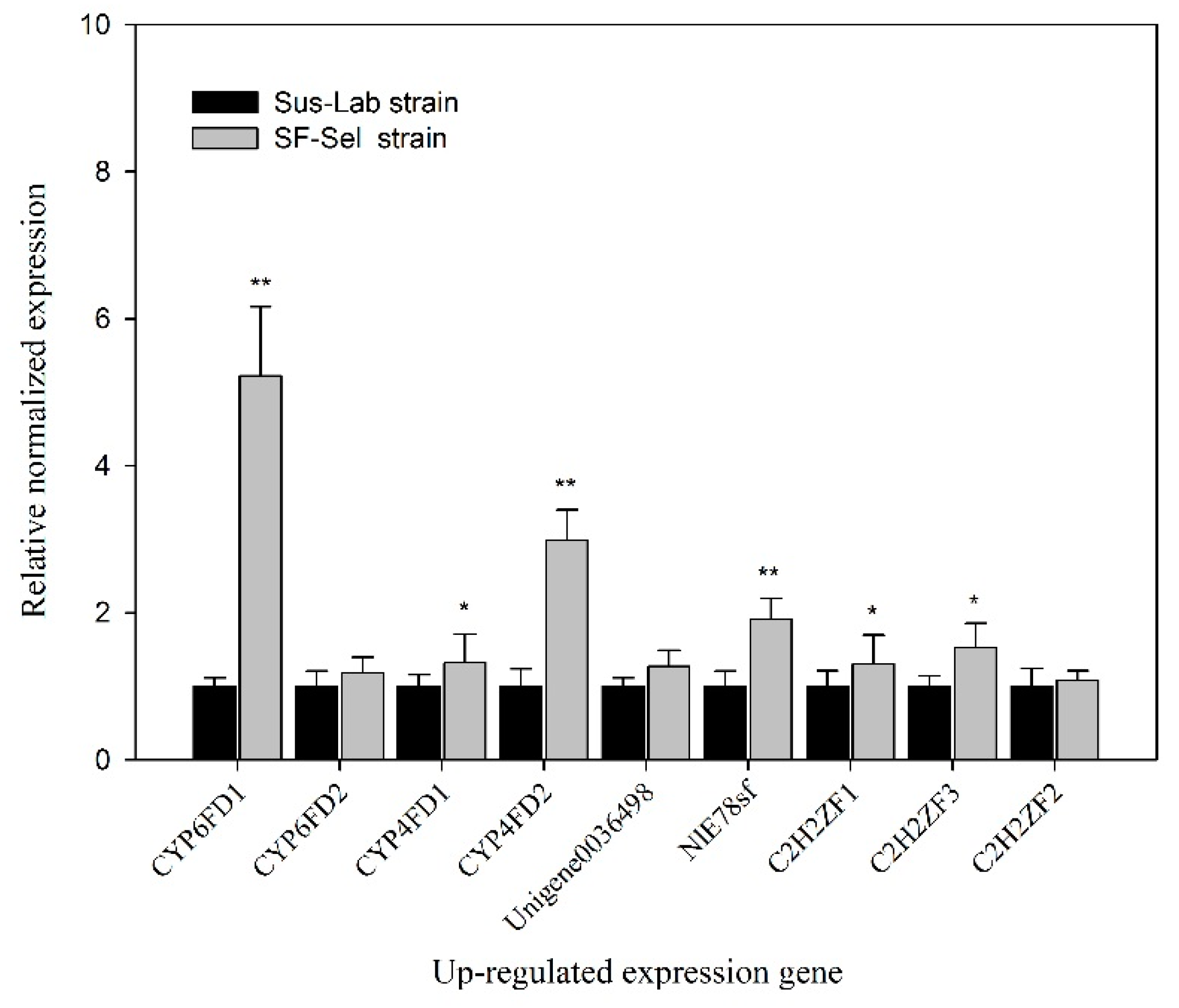
2.9. Screening the Candidate Genes
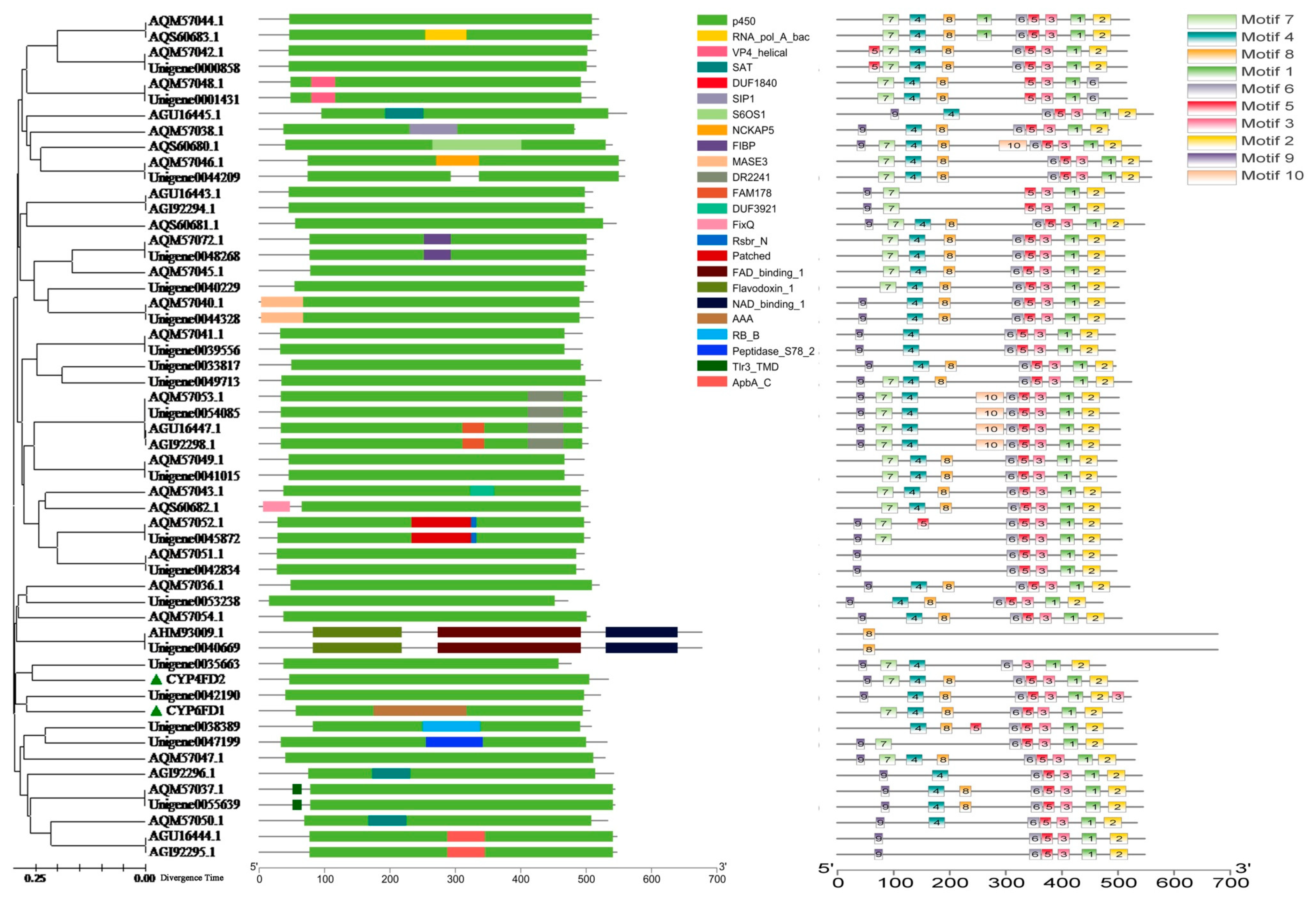
2.10. P450 Diversity Analysis
2.11. Quantitative PCR (qRT-PCR)
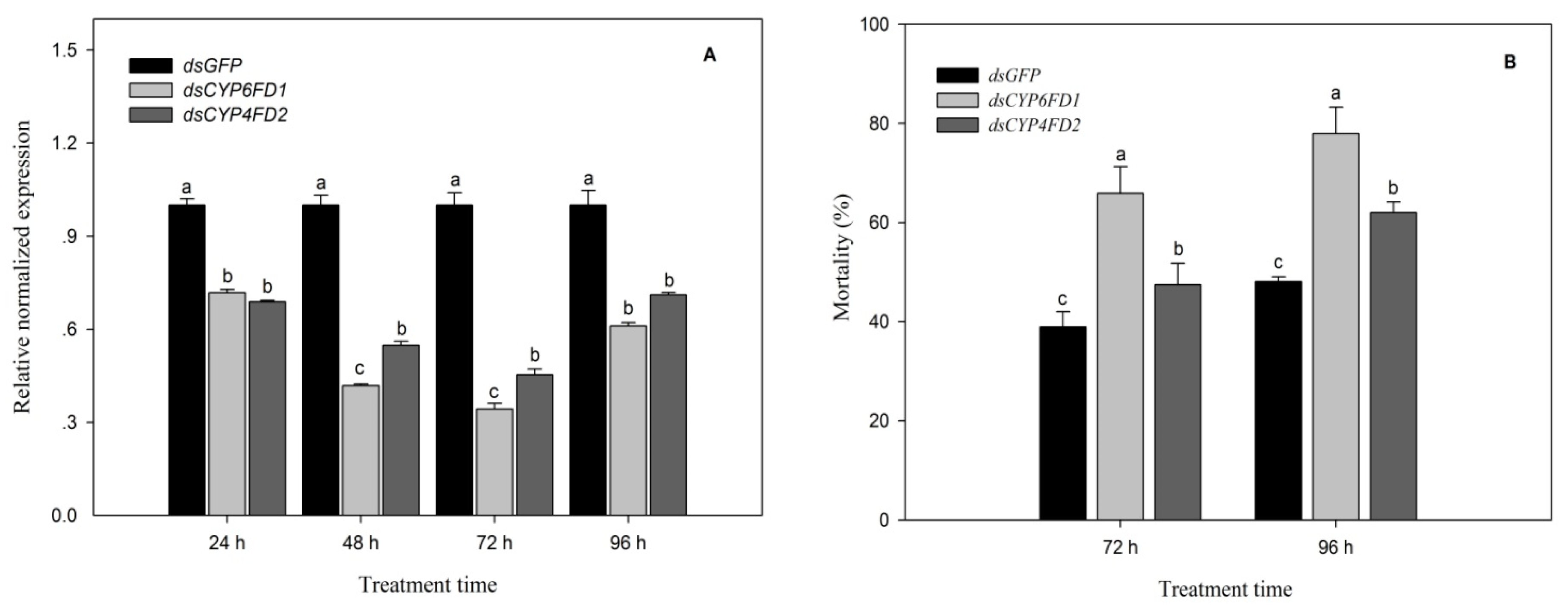
2.12. Functional Analysis of CYP6FD1 and CYP4FD2 via RNAi
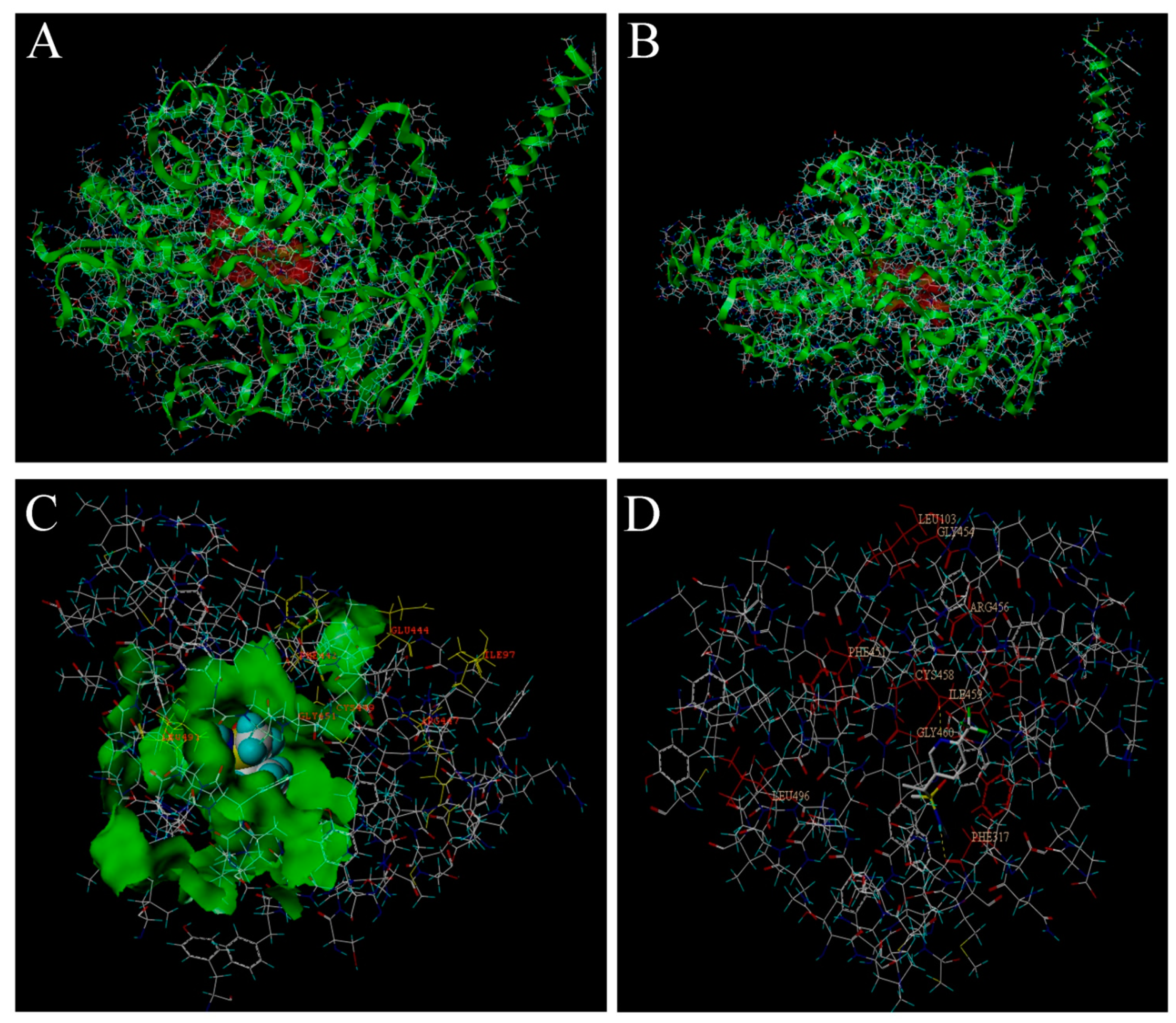
2.13. Interaction of the Tertiary Structure of CYP6FD1 and CYP4FD2 with Sulfoxaflor
3. Discussion
4. Materials and Methods
4.1. Insects and Insecticide
4.2. Selection of the SF-Sel strain with a Sublethal Dose of Sulfoxaflor
4.3. Test for Synergism
4.4. Enzyme Assays
4.5. Transcriptome Analysis
4.5.1. Library Construction and Sequencing, Illumina Read Processing, and Assembly and Annotation of Unigenes
4.5.2. Gene Expression and Differential Gene Enrichment
4.5.3. Diversity and Collinearity of S. furcifera P450 Genes
4.6. Quantitative PCR (qRT-PCR)
4.7. Function Analysis of CYP6FD1 and CYP4FD2 via RNAi
4.8. Prediction the Interaction of Tertiary Structure of CYP6FD1 and CYP4FD2 with Sulfoxaflor
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, X.L.; Liao, X.; Mao, K.K.; Yang, P.; Li, D.Y.; Ali, E.; Wan, H.; Li, J.H. Neonicotinoid insecticide resistance in the field populations of Sogatella furcifera (Horváth) in Central China from 2011 to 2015. J. Asia-Pacific Entomol. 2017, 20, 955–958. [Google Scholar] [CrossRef]
- Horgan, F.G.; Srinivasan, T.S.; Naik, B.S.; Ramal, A.F.; Bernal, C.C.; Almazan, M.L.P. Effects of nitrogen on egg-laying inhibition and ovicidal response in planthopper-resistant rice varieties. Crop. Prot. 2016, 89, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhou, C.; Long, G.Y.; Yang, H.; Jin, D.C. Sublethal effects of buprofezin on development, reproduction, and chitin synthase 1 gene (SfCHS1) expression in the white-backed planthopper, Sogatella furcifera. J. Asia-Pacific Entomol. 2018, 21, 585–591. [Google Scholar] [CrossRef]
- Cheng, Z.N.; Li, S.; Gao, R.Z.; Sun, F.; Liu, W.C.; Zhou, G.H.; Wu, J.X.; Zhou, X.P.; Zhou, Y.J. Distribution and genetic diversity of southern rice black-streaked dwarf virus in China. Virol. J. 2013, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, K.; Towata, T.; Sakai, J.; Onuki, M.; Okuda, M.; Matsumura, M. Dynamics of Southern rice black-streaked dwarf virus in rice and implication for virus acquisition. Phytopathology 2013, 5, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.G.; Jiang, C.X.; Mao, M.; Liu, C.; Li, Q.; Wang, X.G.; Yang, Q.F.; Wang, H.J. Effect of the insecticide dinotefuran on the ultrastructure of the flight muscle of female Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2017, 110, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Endo, S.; Tsurumach, I.M. Insecticide susceptibility of the brown planthopper and the white-backed planthopper collected from Southeast Asia. J. Pestic. Sci. 2001, 26, 82–86. [Google Scholar] [CrossRef]
- Jin, J.X.; Jin, D.C.; Li, W.H.; Cheng, Y.; Li, F.L.; Zhou, Y.H.; Zhang, Y.L.; Liu, L. Status of insecticide resistance in Sogatella furcifera (Horváth)(Hemiptera: Delphacidae) from Guizhou Province. J. Nanjing Agric. Univ. 2017, 40, 258–265. [Google Scholar]
- Matsumura, M.; Takeuchi, H.; Satoh, M.; Sanada-Morimura, S.; Otuka, A.; Watanabe, T.; Thanh, D.V. Species-specific insecticide resistance to imidacloprid and fipronil in the rice planthoppers Nilaparvata lugens and Sogatella furcifera in east and southeast Asia. Pest Manag. Sci. 2008, 64, 1115–1121. [Google Scholar] [CrossRef]
- Tang, J.; Li, J.; Shao, Y.; Yang, B.J.; Liu, Z.W. Fipronil resistance in the whitebacked planthopper (Sogatella furcifera): possible resistance mechanisms and cross-resistance. Pest Manag. Sci. 2010, 66, 121–125. [Google Scholar] [CrossRef]
- Matsumura, M.; Sanada-Morimura, S.; Otuka, A.; Ohtsu, R.; Sakumoto, S.; Takeuchia, H.; Satoha, M. Insecticide susceptibilities in populations of two rice planthoppers, Nilaparvata lugens and Sogatella furcifera, immigrating into Japan in the period 2005−2012. Pest Manag. Sci. 2014, 70, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, W.; Zhang, S.; Wu, S.F.; Ban, L.F.; Su, J.Y.; Gao, C.F. Susceptibility of Sogatella furcifera and Laodelphax striatellus (Hemiptera: Delphacidae) to six insecticides in China. J. Econ. Entomol. 2014, 107, 1916–1922. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.C.; Zhang, W.; Wang, L.X.; Zhang, S.; Zhang, K.; Gao, C.F.; Wu, S.F. Resistance monitoring and cross-resistance patterns of three rice planthoppers, Nilaparvata lugens, Sogatella furcifera and Laodelphax striatellus to dinotefuran in China. Pestic. Biochem. Physiol. 2016, 134, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Gujar, G.T. Local variation in susceptibility of the diamondback moth, Plutella xylostella (Linnaeus) to insecticides and role of detoxification enzymes. Crop Prot. 2009, 22, 495–504. [Google Scholar] [CrossRef]
- Lai, T.C.; Li, J.; Su, J.Y. Monitoring of beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) resistance to chlorantraniliprole in China. Pestic. Biochem. Physiol. 2011, 101, 198–205. [Google Scholar] [CrossRef]
- Wang, X.G.; Xiang, X.; Yu, H.L.; Liu, S.H.; Yin, Y.; Cui, P.; Wu, Y.Q.; Yang, J.; Jiang, C.X.; Yang, Q.F. Monitoring and biochemical characterization of beta-cypermethrin resistance in Spodoptera exigua (Lepidoptera: Noctuidae) in Sichuan Province, China. Pestic. Biochem. Physiol. 2018, 146, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.; Loso, M.; Babcock, J.; Hasler, J.; Letherer, T.; Young, C. Novel nicotinic action of the sulfoximine insecticide sulfoxaflor. Ins. Biochem. Mol. Biol. 2011, 41, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Babcock, J.M.; Gerwick, C.B.; Huang, J.X.; Loso, M.R.; Nakamura, G.; Nolting, S.P.; Rogers, R.B.; Sparks, T.C.; Thomas, J.; Watson, G.B.; et al. Biological characterization of sulfoxaflor, a novel insecticide. Pest Manag. Sci. 2011, 67, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Loso, M.R.; Watson, G.B.; Sparks, T.C.; Rogers, R.B.; Huang, J.X.; Gerwick, B.C.; Babcock, J.M.; Kelley, D.; Hegde, V.B.; et al. Discovery and characterization of sulfoxaflor, a novel insecticide targeting sap-feeding pests. J. Agric. Food Chem. 2011, 59, 2950–2957. [Google Scholar] [CrossRef]
- Longhurst, C.; Babcock, J.M.; Denholm, I.; Gorman, K.; Thomas, J.D.; Sparks, T.C. Cross-resistance relationships of the sulfoximine insecticide sulfoxaflor with neonicotinoids and other insecticides in the whiteflies Bemisia tabaci and Trialeurodes vaporariorum. Pest Manag. Sci. 2013, 69, 809–813. [Google Scholar] [CrossRef]
- Chen, X.; Ma, K.; Li, F.; Liang, P.; Liu, Y.; Guo, T.; Song, D.; Desneux, N.; Gao, X.W. Sublethal and transgenerational effects of sulfoxaflor on the biological traits of the cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). Ecotoxicology 2016, 25, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Cutler, P.; Slater, R.; Edmunds, A.J.; Maienfisch, P.; Hall, R.G.; Earley, F.G.; Pitterna, T.; Pal, S.; Paul, V.L.; Goodchild, J.; et al. Investigating the mode of action of sulfoxaflor: A fourth-generation neonicotinoid. Pest Manag. Sci. 2013, 69, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Mao, K.K.; Ali, E.; Zhang, X.L.; Li, J.H. Temporal variability and resistance correlation of sulfoxaflor susceptibility among Chinese populations of the brown planthopper Nilaparvata lugens (Stål). Crop Prot. 2017, 102, 141–146. [Google Scholar] [CrossRef]
- Liao, X.; Jin, R.H.; Zhang, X.L.; Ali, E.; Mao, K.K.; Xu, P.F.; Li, J.H.; Wan, H.H. Characterization of sulfoxaflor resistance in the brown planthopper, Nilaparvata lugens (Stål). Pest Manag. Sci. 2019, 275, 1646–1654. [Google Scholar] [CrossRef]
- Ma, K.S.; Tang, Q.L.; Zhang, B.Z.; Liang, P.; Wang, B.M.; Gao, X.W. Overexpression of multiple cytochrome P450 genes associated with sulfoxaflor resistance in Aphis gossypii Glover. Pestic. Biochem. Physiol. 2019, 157, 204–210. [Google Scholar] [CrossRef]
- Reyes, M.; Rocha, K.; Alarcón, L.; Siegwart, M.; Sauphanor, B. Metabolic mechanisms involved in the resistance of field populations of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) to spinosad. Pestic. Biochem. Phys. 2012, 102, 45–50. [Google Scholar] [CrossRef]
- Tian, X.R.; Sun, X.X.; Su, J.Y. Biochemical mechanisms for metaflumizone resistance in beet armyworm, Spodoptera exigua. Pestic. Biochem. Phys. 2014, 113, 8–14. [Google Scholar] [CrossRef]
- Chen, E.H.; Dou, W.; Hu, F.; Tang, S.; Wang, J.J. Purification and biochemical characterization of glutathione S-transferases in Bactrocera minax (Diptera: Tephritidae). Fla. Entomol. 2012, 95, 593–601. [Google Scholar] [CrossRef]
- Wei, X.; Pan, Y.; Xin, X.C.; Zheng, C.; Gao, X.W.; Xi, J.H.; Shang, Q.L. Cross-resistance pattern and basis of resistance in a thiamethoxam-resistant strain of Aphis gossypii Glover. Pestic. Biochem. Phys. 2017, 138, 91–96. [Google Scholar] [CrossRef]
- Mao, K.K.; Zhang, X.L.; Ali, E.; Liao, X.; Jin, R.H.; Ren, Z.J.; Wan, H.; Li, J.H. Characterization of nitenpyram resistance in Nilaparvata lugens (Stål). Pestic. Biochem. Phys. 2019, 157, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.B.; Gao, H.L.; Zhang, Y.X.; Fan, D.; Fang, J.; Liu, Z. The roles of CYP6AY1 and CYP6ER1 in imidacloprid resistance in the brown planthopper: Expression levels and detoxification efficiency. Pestic. Biochem. Phys. 2016, 129, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.X.; Jin, D.C.; Li, F.L.; Cheng, Y.; Li, W.H.; Ye, Z.C.; Zhou, Y.H. Expression differences of resistance-related genes induced by cycloxaprid using qRT-PCR in the female adult of Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2017, 110, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, C.T.; Garrood, W.T.; Singh, K.S.; Randall, E.; Lueke, B.; Gutbrod, O.; Matthiesen, S.; Kohler, M.; Nauen, R.; Davies, T.G.E.; et al. Neofunctionalization of duplicated P450 genes drives the evolution of insecticide resistance in the brown planthopper. Curr. Biol. 2018, 28, 268. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Daniels, M.; Andrews, M.; Lind, R.J.; Gorman, K.; Williamson, M.S. Age-specific expression of a P450 monooxygenase (CYP6CM1) correlates with neonicotinoid resistance in Bemisia tabaci. Pestic. Biochem. Phys. 2011, 101, 53–58. [Google Scholar] [CrossRef]
- Ding, Z.P.; Wen, Y.C.; Yang, B.J.; Zhang, Y.X.; Liu, S.H.; Liu, Z.W.; Han, Z.J. Biochemical mechanisms of imidacloprid resistance in Nilaparvata lugens: Over-expression of cytochrome P450 CYP6AY1. Insect Biochem. Molec. 2013, 43, 1021–1027. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y. Identification and characterisation of NADPH-dependent cytochrome P450 reductase gene and cytochrome b5 gene from Plutella xylostella: Possible involvement in resistance to beta-cypermethrin. Gene 2015, 558, 208–214. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Yocum, G.D.; Rinehart, J.P. Hormonal control of diapause. Compr. Mol. Insect Sci. 2005, 615–650. [Google Scholar]
- Stiborová, M.; Indra, R.; Frei, E.; Schmeiser, H.H.; Eckschlager, T.; Adam, V.; Heger, Z.; Arlt, V.M.; Martínek, V. Cytochrome b5 plays a dual role in the reaction cycle of cytochrome P450 3A4 during oxidation of the anticancer drug ellipticine. Monatsh. Chem. 2017, 148, 1983–1991. [Google Scholar] [CrossRef]
- Cui, S.F.; Wang, L.; Ma, L.; Geng, X.Q. P450-mediated detoxification of botanicals in insects. Phytoparasitica 2016, 44, 585–599. [Google Scholar] [CrossRef]
- Lertkiatmongkol, P.; Jenwitheesuk, E.; Rongnoparut, P. Homology modeling of mosquito cytochrome P450 enzymes involved in pyrethroid metabolism, insights into differences in substrate selectivity. BMC Res. Notes 2011, 4, 321. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zhu, Y.C.; Ma, C.; Huang, Y.; Shen, J. Susceptibility to neonicotinoids and risk of resistance development in the brown planthopper, Nilaparvata lugens (stål) (Homoptera: Delphacidae). Pest Manag. Sci. 2010, 64, 1278–1284. [Google Scholar] [CrossRef] [PubMed]
- Van, A.K. A study of housefly esterase by means of sensitive colorimetric-method. J. Insect Physiol. 1962, 8, 401–406. [Google Scholar]
- Rose, R.L.; Barbhaiya, L.; Roe, R.M. Cytochrome P450-associated insecticide resistance and the development of biochemical diagnostic assays in Heliothis virescens. Pestic. Biochem. Phys. 1995, 51, 178–191. [Google Scholar] [CrossRef]
- Bradford, M.M.A. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 25, 248–256. [Google Scholar] [CrossRef]
- Wang, X.G.; Chen, Y.Q.; Gong, C.W.; Yao, X.G.; Jiang, C.X.; Yang, Q.F. Molecular identification of four novel cytochrome P450 genes related to the development of resistance of Spodoptera exigua (Lepidoptera: Noctuidae) to chlorantraniliprole. Pest Manag. Sci. 2018, 74, 1938–1952. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods. 2008, 5, 621–628. [Google Scholar] [CrossRef]
- An, X.K.; Hou, M.L.; Liu, Y.D. Reference gene selection and evaluation for gene expression studies using qRT-PCR in the white-backed Planthopper, Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2016, 109, 879. [Google Scholar] [CrossRef]
- Wang, L.X.; Niu, C.D.; Zhang, Y.; Jia, Y.L.; Zhang, Y.J.; Zhang, Y.; Zhang, Y.Q.; Gao, C.F.; Wu, S.F. The NompC channel regulates Nilaparvata lugens proprioception and gentletouch response. Insect Biochem. Mol. Biol. 2019, 106, 55–63. [Google Scholar] [CrossRef]
- Revankar, H.M.; Kulkarni, M.V.; Joshi, S.D.; More, U.A. Synthesis, biological evaluation and docking studies of 4-aryloxymethyl coumarins derived from substructures and degradation products of vancomycin. Eur. J. Med. Chem. 2013, 70, 750–757. [Google Scholar] [CrossRef]
- Jain, A.N. Scoring noncovalent protein-ligand interactions: Acontinuous differentiable function tuned to compute binding affinities. J. Comput. Aid. Mol. Des. 1996, 10, 427–440. [Google Scholar] [CrossRef]
- Jain, A.N. Surflex: Fully automatic flexible molecular docking using a molecular similarity-based search engine. J. Med. Chem. 2003, 46, 499–511. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Treatment | LC50 (µg/mL) 95%CI | Slope ± SE | χ2 (df) | SR * |
|---|---|---|---|---|---|
| Sus-Lab | sulfoxaflor | 3.544(3.287–3.804) | 6.499 ± 0.722 | 11.226(13) | / |
| sulfoxaflor plus TPP | 2.371(2.051–2.700) | 3.253 ± 0.418 | 4.864(13) | 1.495 | |
| sulfoxaflor plus DEM | 2.940(2.627–3.282) | 4.069 ± 0.512 | 4.997(13) | 1.205 | |
| sulfoxaflor plus PBO | 2.927(2.549–3.212) | 6.893 ± 1.199 | 10.213(13) | 1.211 | |
| SF-Sel | sulfoxaflor | 7.284(6.265–8.385) | 4.101 ± 0.454 | 14.726(13) | / |
| sulfoxaflor plus TPP | 2.709(2.180–3.449) | 1.972 ± 0.248 | 4.292(13) | 2.689 | |
| sulfoxaflor plus DEM | 3.191(2.529–4.061) | 2.200 ± 0.307 | 3.268(13) | 2.283 | |
| sulfoxaflor plus PBO | 1.367(1.129–1.648) | 2.829 ± 0.353 | 4.908(13) | 5.328 |
| GeneID | log2 Ratio(S/M) | S_vs_M Regulated | Gene Type | Annotation |
|---|---|---|---|---|
| Unigene0005814 | 11.17326071 | UP | CYP6FD1 | P450 |
| Unigene0012458 | 10.38247993 | UP | ||
| Unigene0020537 | 9.861035196 | UP | CYP6FD2 | P450 |
| Unigene0020536 | 9.83315364 | UP | ||
| Unigene0069588 | 8.681589817 | UP | CYP4FD1 | P450 |
| Unigene0015479 | 7.934280594 | UP | ||
| Unigene0027543 | 2.07289003 | UP | CYP4FD2 | P450 |
| Unigene0036498 | 8.17697521 | UP | Transporter | |
| Unigene0042782 | 1.136503653 | UP | C2H2ZF2 | Transcription factors |
| Unigene0051504 | 1.044721874 | UP | NlE78sf | Transcription factors |
| Unigene0010562 | 1.411318813 | UP | C2H2ZF1 | Transcription factors |
| Unigene0010210 | 1.036525876 | UP | C2H2ZF3 | Transcription factors |
| Gene Family | Prime | Sequence (5′-3′) | Length |
|---|---|---|---|
| Reference | RPL9-F | TGTGTGACCACCGAGAACAACTCA | 131 |
| RPL9-R | ACGATGAGCTCGTCCTTCTGCTTT | ||
| P450 | CYP6FD1-F | CTTCAACATGCGGTTCACGC | 187 |
| CYP6FD1-R | TTCATCCAAGCTCAACGGCT | ||
| CYP4FD1-F | AACCACTGCATGACTTTGCC | 199 | |
| CYP4FD1-R | TCAGCACCCGCAATGAATGT | ||
| CYP6FD2-F | GAGATGGCACACAAACCGGA | 171 | |
| CYP6FD2-R | GCAGAATCGCGCTAGAATGG | ||
| CYP4FD2-F | CAGCGAATGGTGGCTTCATC | 183 | |
| CYP4FD2-R | ATAGCAGCCATGGTCTCACC | ||
| Transporter | Unigene0036498-F | CCCAAACCCTTCAAGACGGA | 162 |
| Unigene0036498-R | GGCTGGATCGGAAATGCTCT | ||
| Transcription factor | NlE78sf-F | GGAGTGTTGGGGTGGTAGTG | 181 |
| NlE78sf-R | GGTGATGAACACTGCTCCGA | ||
| C2H2ZF1-F | CCATCATCAAGGCGGAACCT | 182 | |
| C2H2ZF1-R | ACCAGCGTTTTCAATGGTGC | ||
| C2H2ZF3-F | GTCGCCTGTGCCTTCTAGTT | 165 | |
| C2H2ZF3-R | AGCGGATGCACCTGATACTG | ||
| C2H2ZF2-F | ACAAGGGCATTCGCAAACAC | 159 | |
| C2H2ZF2-R | ATGTGCCGATCCAGATAGCG |
| Prime | Sequence (5′-3′) |
|---|---|
| T7-GFP-F | TAATACGACTCACTATAGGGAAGGGCGAGGAGCTGTTCACCG |
| T7-GFP-R | TAATACGACTCACTATAGGGCAGCAGGACCATGTGATCGCGC |
| CYP6FD1dsRNAF | TAATACGACTCACTATAGGGAGAAGTCCCAATTTCACAGACGC |
| CYP6FD1dsRNAR | TAATACGACTCACTATAGGGAGAGATTCCGGTCTATGCGCTTC |
| CYP4FD2dsRNAF | TAATACGACTCACTATAGGGAGAAAGGTTTCATCTACAAAGGATTGC |
| CYP4FD2dsRNAR | TAATACGACTCACTATAGGGAGACATCAGTGAAATCGTGCAGAATC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-G.; Ruan, Y.-W.; Gong, C.-W.; Xiang, X.; Xu, X.; Zhang, Y.-M.; Shen, L.-T. Transcriptome Analysis of Sogatella furcifera (Homoptera: Delphacidae) in Response to Sulfoxaflor and Functional Verification of Resistance-Related P450 Genes. Int. J. Mol. Sci. 2019, 20, 4573. https://doi.org/10.3390/ijms20184573
Wang X-G, Ruan Y-W, Gong C-W, Xiang X, Xu X, Zhang Y-M, Shen L-T. Transcriptome Analysis of Sogatella furcifera (Homoptera: Delphacidae) in Response to Sulfoxaflor and Functional Verification of Resistance-Related P450 Genes. International Journal of Molecular Sciences. 2019; 20(18):4573. https://doi.org/10.3390/ijms20184573
Chicago/Turabian StyleWang, Xue-Gui, Yan-Wei Ruan, Chang-Wei Gong, Xin Xiang, Xiang Xu, Yu-Ming Zhang, and Li-Tao Shen. 2019. "Transcriptome Analysis of Sogatella furcifera (Homoptera: Delphacidae) in Response to Sulfoxaflor and Functional Verification of Resistance-Related P450 Genes" International Journal of Molecular Sciences 20, no. 18: 4573. https://doi.org/10.3390/ijms20184573