Sesquiterpenes Are Agonists of the Pregnane X Receptor but Do Not Induce the Expression of Phase I Drug-Metabolizing Enzymes in the Human Liver

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
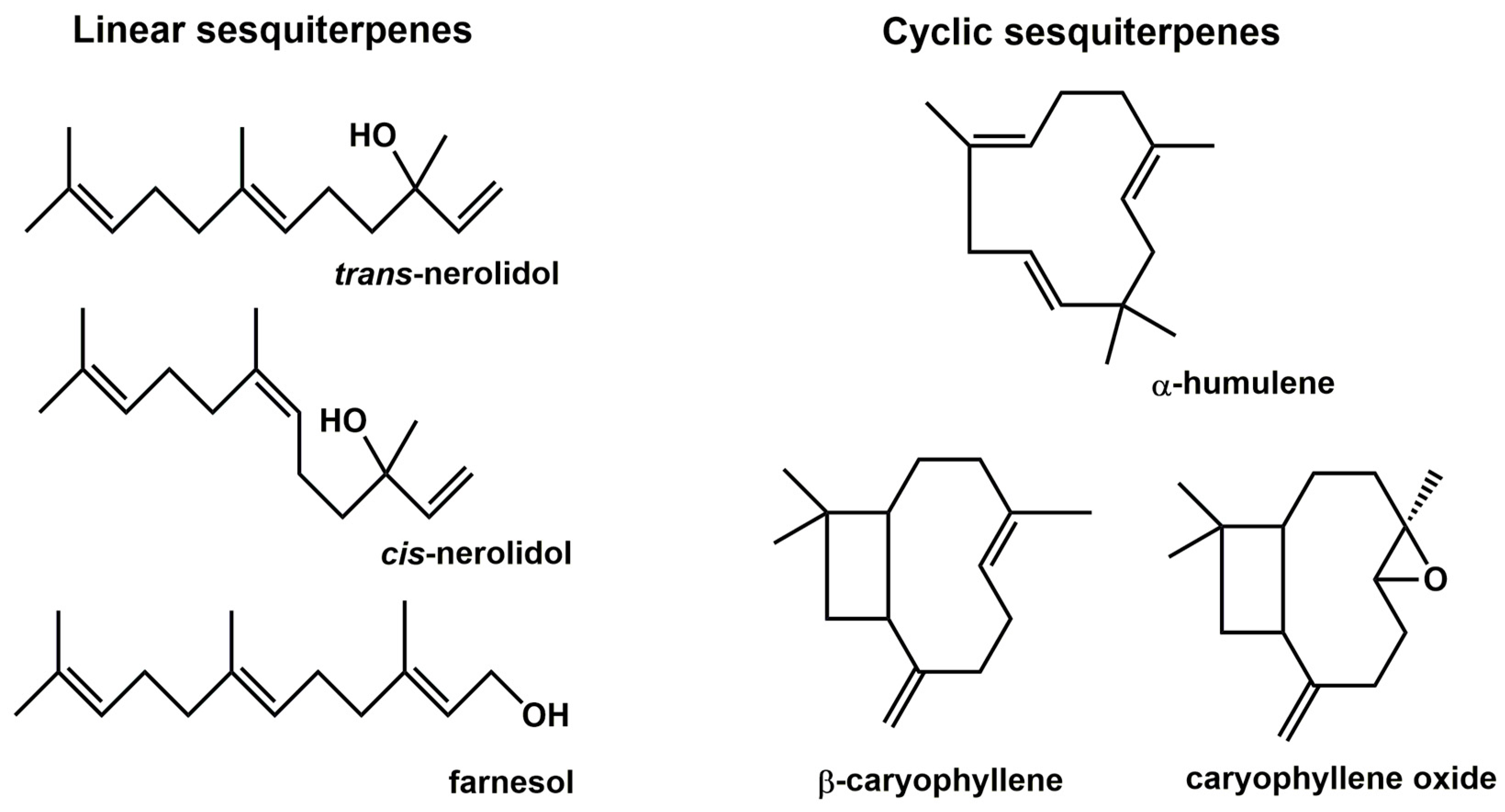
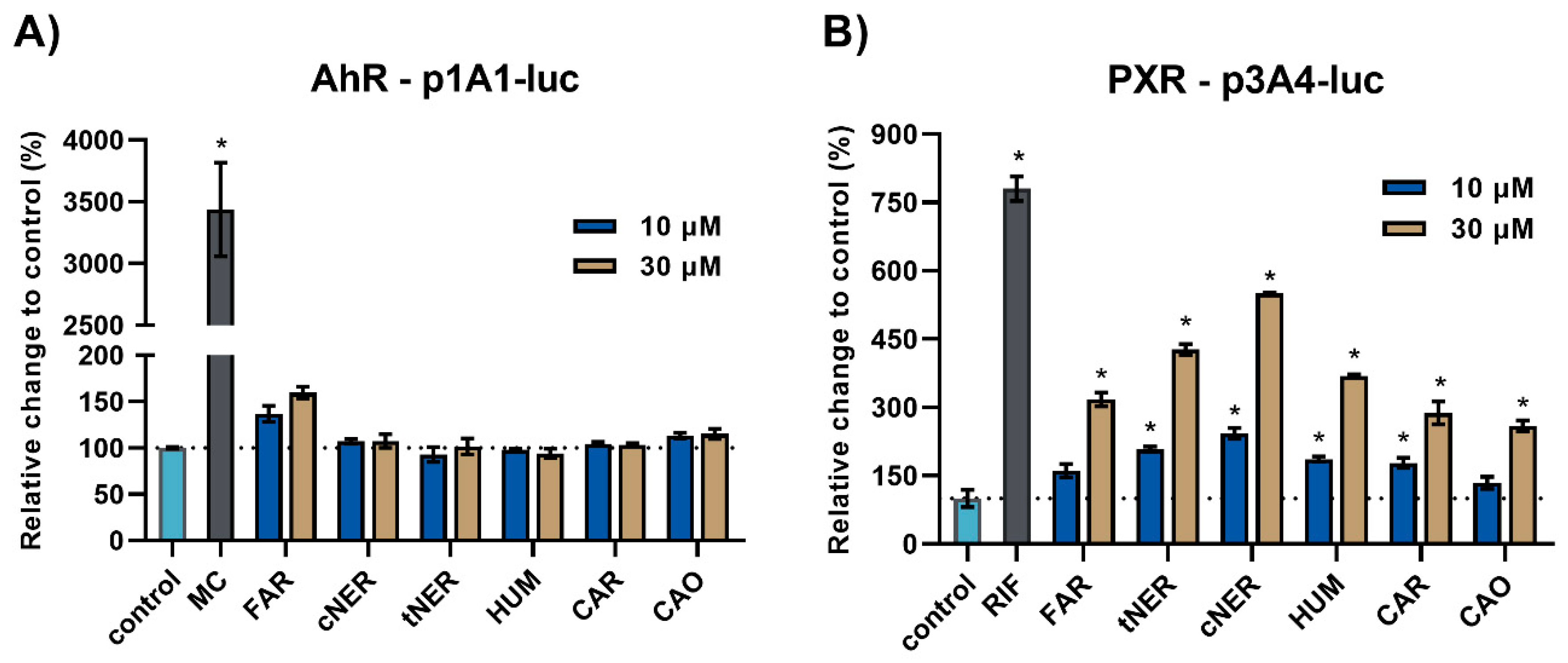
2.1. The Effect of Sesquiterpenes on AhR and PXR Activation
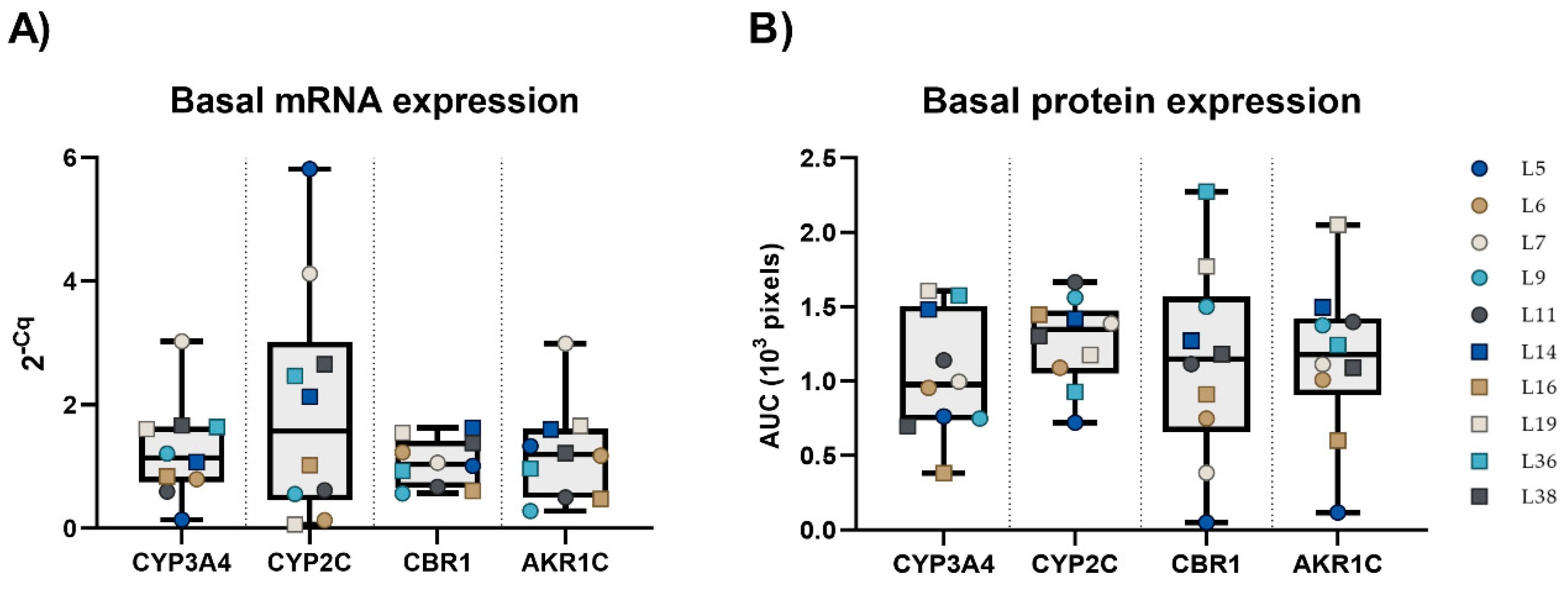
2.2. Basal mRNA and Protein Expression of CYP3A4, CYP2C, CBR1, and AKR1C3 in PCLS
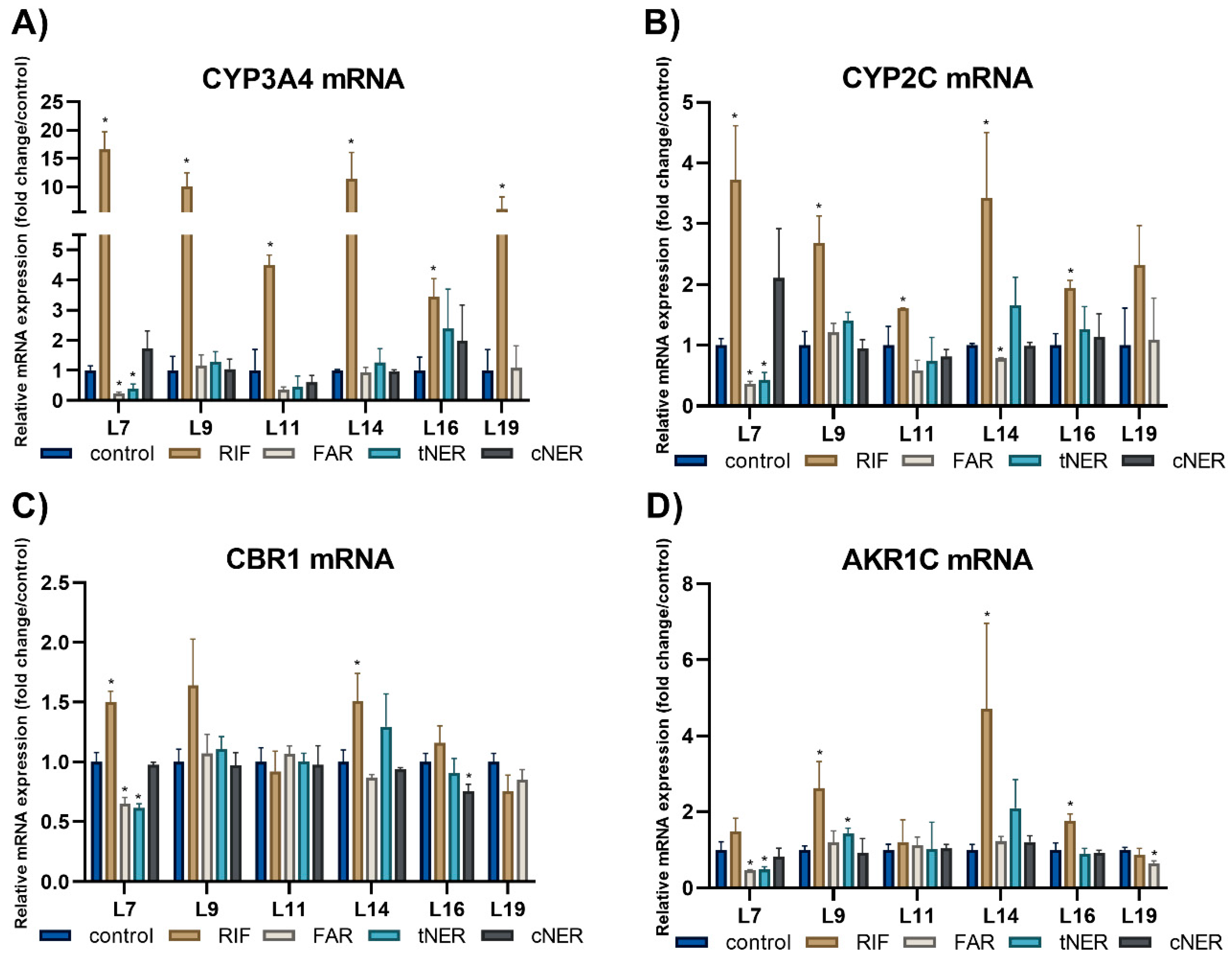
2.3. The Effect of Sesquiterpenes on the mRNA Expression of the Studied Enzymes
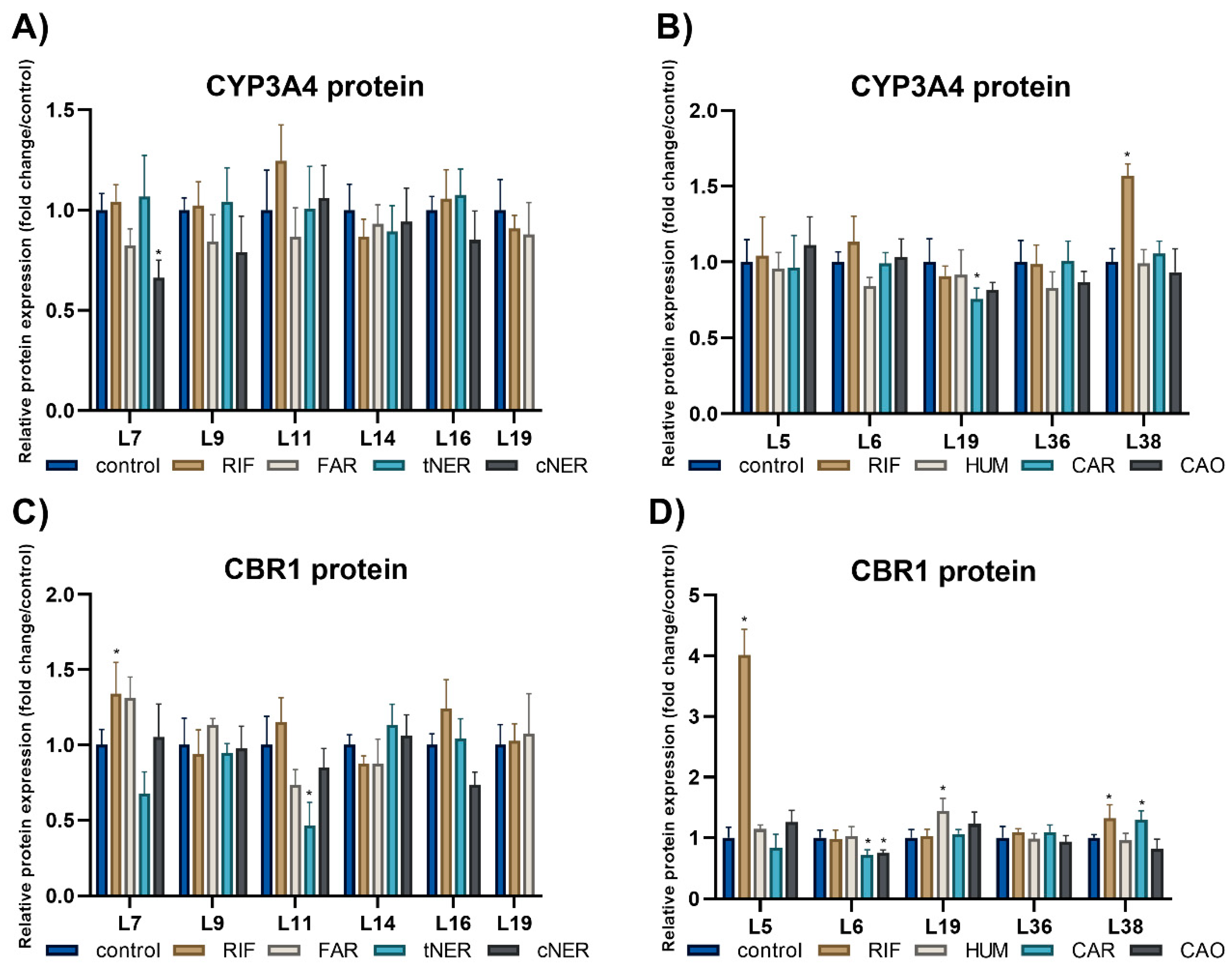
2.4. The Effect of Sesquiterpenes on the Protein Expression of Studied Enzymes
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Cell Culture
3.3. Plasmids
3.4. Luciferase Reporter Gene Assays
3.5. Ethics Committee Statement
3.6. Human Liver Tissue
3.7. Preparation of Precision-Cut Liver Slices and Experimental Treatment
3.8. RNA Isolation, cDNA Synthesis and Quantitative Real-Time PCR (RT-qPCR)
3.9. Western Blotting
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AKR1C | aldo-keto reductase 1C |
| CAO | caryophyllene oxide |
| CAR | β-caryophyllene |
| CBR1 | carbonyl reductase 1 |
| cNER | cis-nerolidol |
| CYP | cytochrome P450 |
| DEPC | diethyl pyrocarbonate |
| DM | diabetes mellitus |
| DME | drug-metabolizing enzyme |
| DMSO | dimethyl sulfoxide |
| FAR | farnesol |
| FBS | fetal bovine serum |
| GADPH | glyceraldehyde 3-phosphate dehydrogenase |
| HLD | hyperlipidemia |
| HUM | α-humulene |
| HTN | hypertension |
| MC | methylcholanthrene |
| RIF | rifampicin |
| RT-qPCR | reverse transcription-quantitative polymerase chain reaction |
| SDHA | succinate dehydrogenase complex, subunit A |
| s/p CVA | status post cerebrovascular accident |
| SDS-PAGE | sodium dodecyl sulfate–polyacrylamide gel electrophoresis |
| TBST-T | TRIS-buffered saline-Tween-20 |
| tNER | trans-nerolidol |
References
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharm. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.A.; Seely, D. Clinically based evidence of drug-herb interactions: A systematic review. Expert. Opin. Drug Saf. 2010, 9, 79–124. [Google Scholar] [CrossRef] [PubMed]
- Awortwe, C.; Bruckmueller, H.; Cascorbi, I. Interaction of herbal products with prescribed medications: A systematic review and meta-analysis. Pharm. Res. 2019, 141, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Agbabiaka, T.B.; Wider, B.; Watson, L.K.; Goodman, C. Concurrent Use of Prescription Drugs and Herbal Medicinal Products in Older Adults: A Systematic Review. Drugs Aging 2017, 34, 891–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonner, P.H.; Kampen, J.; Scholz, J. Pathophysiological changes in the elderly. Best Pr. Res. Clin. Anaesthesiol. 2003, 17, 163–177. [Google Scholar] [CrossRef]
- Bartikova, H.; Hanusova, V.; Skalova, L.; Ambroz, M.; Bousova, I. Antioxidant, pro-oxidant and other biological activities of sesquiterpenes. Curr. Top. Med. Chem. 2014, 14, 2478–2494. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Mysliveckova, Z.; Szotakova, B.; Spicakova, A.; Lnenickova, K.; Ambroz, M.; Kubicek, V.; Krasulova, K.; Anzenbacher, P.; Skalova, L. The inhibitory effects of β-caryophyllene, β-caryophyllene oxide and α-humulene on the activities of the main drug-metabolizing enzymes in rat and human liver in vitro. Chem. Biol. Interact. 2017, 278, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Chaves, J.S.; Leal, P.C.; Pianowisky, L.; Calixto, J.B. Pharmacokinetics and tissue distribution of the sesquiterpene α-humulene in mice. Planta. Med. 2008, 74, 1678–1683. [Google Scholar] [CrossRef]
- Liu, H.; Yang, G.; Tang, Y.; Cao, D.; Qi, T.; Qi, Y.; Fan, G. Physicochemical characterization and pharmacokinetics evaluation of β-caryophyllene/β-cyclodextrin inclusion complex. Int. J. Pharm. 2013, 450, 304–310. [Google Scholar] [CrossRef]
- Saito, A.Y.; Sussmann, R.A.; Kimura, E.A.; Cassera, M.B.; Katzin, A.M. Quantification of nerolidol in mouse plasma using gas chromatography-mass spectrometry. J. Pharm. Biomed. Anal. 2015, 111, 100–103. [Google Scholar] [CrossRef]
- Spicakova, A.; Szotakova, B.; Dimunova, D.; Mysliveckova, Z.; Kubicek, V.; Ambroz, M.; Lnenickova, K.; Krasulova, K.; Anzenbacher, P.; Skalova, L. Nerolidol and Farnesol Inhibit Some Cytochrome P450 Activities but Did Not Affect Other Xenobiotic-Metabolizing Enzymes in Rat and Human Hepatic Subcellular Fractions. Molecules 2017, 22, 590. [Google Scholar] [CrossRef] [PubMed]
- Lnenickova, K.; Svobodova, H.; Skalova, L.; Ambroz, M.; Novak, F.; Matouskova, P. The impact of sesquiterpenes β-caryophyllene oxide and trans-nerolidol on xenobiotic-metabolizing enzymes in mice in vivo. Xenobiotica 2018, 48, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, I.A.; Olinga, P.; de Jager, M.H.; Merema, M.T.; de Kanter, R.; van de Kerkhof, E.G.; Groothuis, G.M. Preparation and incubation of precision-cut liver and intestinal slices for application in drug metabolism and toxicity studies. Nat. Protoc. 2010, 5, 1540–1551. [Google Scholar] [CrossRef] [PubMed]
- Olinga, P.; Schuppan, D. Precision-cut liver slices: A tool to model the liver ex vivo. J. Hepatol. 2013, 58, 1252–1253. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.J.; Price, R.J.; Watts, P.S.; Renwick, A.B.; Tredger, J.M.; Boobis, A.R.; Lake, B.G. Induction of cytochrome P450 enzymes in cultured precision-cut human liver slices. Drug Metab. Dispos. 2003, 31, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Faugeras, L.; Dili, A.; Druez, A.; Krug, B.; Decoster, C.; D’Hondt, L. Treatment options for metastatic colorectal cancer in patients with liver dysfunction due to malignancy. Crit. Rev. Oncol. Hematol. 2017, 115, 59–66. [Google Scholar] [CrossRef]
- Ciombor, K.K.; Goff, L.W. Current therapy and future directions in biliary tract malignancies. Curr. Treat. Opt. Oncol. 2013, 14, 337–349. [Google Scholar] [CrossRef] [PubMed]
- He, W.Z.; Guo, G.F.; Yin, C.X.; Jiang, C.; Wang, F.; Qiu, H.J.; Chen, X.X.; Rong, R.M.; Zhang, B.; Xia, L.P. Gamma-glutamyl transpeptidase level is a novel adverse prognostic indicator in human metastatic colorectal cancer. Colorectal Dis. 2013, 15, e443–e452. [Google Scholar] [CrossRef]
- Lehmann, J.M.; McKee, D.D.; Watson, M.A.; Willson, T.M.; Moore, J.T.; Kliewer, S.A. The human orphan nuclear receptor PXR is activated by compounds that regulate CYP3A4 gene expression and cause drug interactions. J. Clin. Invest. 1998, 102, 1016–1023. [Google Scholar] [CrossRef]
- Whitlock, J.P., Jr. Induction of cytochrome P4501A1. Annu. Rev. Pharm. Toxicol. 1999, 39, 103–125. [Google Scholar] [CrossRef]
- Nebert, D.W.; Dalton, T.P.; Okey, A.B.; Gonzalez, F.J. Role of aryl hydrocarbon receptor-mediated induction of the CYP1 enzymes in environmental toxicity and cancer. J. Biol. Chem. 2004, 279, 23847–23850. [Google Scholar] [CrossRef] [PubMed]
- Pimkaew, P.; Kublbeck, J.; Petsalo, A.; Jukka, J.; Suksamrarn, A.; Juvonen, R.; Auriola, S.; Piyachaturawat, P.; Honkakoski, P. Interactions of sesquiterpenes zederone and germacrone with the human cytochrome P450 system. Toxicol. In Vitro 2013, 27, 2005–2012. [Google Scholar] [CrossRef] [PubMed]
- Burk, O.; Arnold, K.A.; Nussler, A.K.; Schaeffeler, E.; Efimova, E.; Avery, B.A.; Avery, M.A.; Fromm, M.F.; Eichelbaum, M. Antimalarial artemisinin drugs induce cytochrome P450 and MDR1 expression by activation of xenosensors pregnane X receptor and constitutive androstane receptor. Mol. Pharm. 2005, 67, 1954–1965. [Google Scholar] [CrossRef] [PubMed]
- Burk, O.; Piedade, R.; Ghebreghiorghis, L.; Fait, J.T.; Nussler, A.K.; Gil, J.P.; Windshugel, B.; Schwab, M. Differential effects of clinically used derivatives and metabolites of artemisinin in the activation of constitutive androstane receptor isoforms. Br. J. Pharm. 2012, 167, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Therapeut. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Bousova, I.; Skalova, L.; Soucek, P.; Matouskova, P. The modulation of carbonyl reductase 1 by polyphenols. Drug Metab. Rev. 2015, 47, 520–533. [Google Scholar] [CrossRef] [PubMed]
- Penning, T.M.; Wangtrakuldee, P.; Auchus, R.J. Structural and Functional Biology of Aldo-Keto Reductase Steroid-Transforming Enzymes. Endocr. Rev. 2019, 40, 447–475. [Google Scholar] [CrossRef] [PubMed]
- Fischedick, J.T.; Standiford, M.; Johnson, D.A.; De Vos, R.C.; Todorovic, S.; Banjanac, T.; Verpoorte, R.; Johnson, J.A. Activation of antioxidant response element in mouse primary cortical cultures with sesquiterpene lactones isolated from Tanacetum parthenium. Planta. Med. 2012, 78, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yoshida, C.; Murakami, A.; Ohigashi, H.; Osawa, T.; Uchida, K. Zerumbone, a tropical ginger sesquiterpene, activates phase II drug metabolizing enzymes. Febs. Lett. 2004, 572, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Achour, B.; Barber, J.; Rostami-Hodjegan, A. Expression of hepatic drug-metabolizing cytochrome p450 enzymes and their intercorrelations: A meta-analysis. Drug Metab. Dispos. 2014, 42, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.; Sarsat, J.P.; de Waziers, I.; Housset, C.; Balladur, P.; Beaune, P.; Albaladejo, V.; Lerche-Langrand, C. Induction of cytochrome P450 2B6 and 3A4 expression by phenobarbital and cyclophosphamide in cultured human liver slices. Pharm. Res. 2003, 20, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Persson, K.P.; Ekehed, S.; Otter, C.; Lutz, E.S.; McPheat, J.; Masimirembwa, C.M.; Andersson, T.B. Evaluation of human liver slices and reporter gene assays as systems for predicting the cytochrome p450 induction potential of drugs in vivo in humans. Pharm. Res. 2006, 23, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.Z.; Lv, Q.L.; Wu, N.Y.; Cheng, L.; Chu, Y.C.; Chu, T.Y.; Hu, L.; Cheng, Y.; Zhang, X.; Zhou, H.H. Mechanism-based inhibition of Alantolactone on human cytochrome P450 3A4 in vitro and activity of hepatic cytochrome P450 in mice. J. Ethnopharmacol. 2015, 168, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Pelkonen, O.; Turpeinen, M.; Hakkola, J.; Honkakoski, P.; Hukkanen, J.; Raunio, H. Inhibition and induction of human cytochrome P450 enzymes: Current status. Arch. Toxicol. 2008, 82, 667–715. [Google Scholar] [CrossRef] [PubMed]
- Staines, A.G.; Sindelar, P.; Coughtrie, M.W.; Burchell, B. Farnesol is glucuronidated in human liver, kidney and intestine in vitro, and is a novel substrate for UGT2B7 and UGT1A1. Biochem. J. 2004, 384, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, S.; Xie, Z.; Pavlovicz, R.E.; Wu, J.; Chen, P.; Aimiuwu, J.; Pang, J.; Bhasin, D.; Neviani, P.; et al. Modulation of DNA methylation by a sesquiterpene lactone parthenolide. J. Pharm. Exp. 2009, 329, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Gopal, Y.N.; Arora, T.S.; Van Dyke, M.W. Parthenolide specifically depletes histone deacetylase 1 protein and induces cell death through ataxia telangiectasia mutated. Chem. Biol 2007, 14, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.Z.; Gao, W.; Yu, A.M. MicroRNAs regulate CYP3A4 expression via direct and indirect targeting. Drug Metab. Dispos. 2009, 37, 2112–2117. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.W.; Zhang, Y.F.; Li, Y.; Xu, Y.Z.; Li, Z.H.; Lu, H.; Zhu, Y.G.; Liu, Z.X.; Tian, Z.Q. Isoalantolactone Inhibits Esophageal Squamous Cell Carcinoma Growth Through Downregulation of MicroRNA-21 and Derepression of PDCD4. Dig. Dis. Sci. 2018, 63, 2285–2293. [Google Scholar] [CrossRef]
- Zuo, W.; Wang, Z.Z.; Xue, J. Artesunate induces apoptosis of bladder cancer cells by miR-16 regulation of COX-2 expression. Int. J. Mol. Sci. 2014, 15, 14298–14312. [Google Scholar] [CrossRef]
- Koe, X.F.; Lim, E.L.; Seah, T.C.; Amanah, A.; Wahab, H.A.; Adenan, M.I.; Sulaiman, S.F.; Tan, M.L. Evaluation of in vitro cytochrome P450 induction and inhibition activity of deoxyelephantopin, a sesquiterpene lactone from Elephantopus scaber L. Food Chem. Toxicol. 2013, 60, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Svecova, L.; Vrzal, R.; Burysek, L.; Anzenbacherova, E.; Cerveny, L.; Grim, J.; Trejtnar, F.; Kunes, J.; Pour, M.; Staud, F.; et al. Azole antimycotics differentially affect rifampicin-induced pregnane X receptor-mediated CYP3A4 gene expression. Drug Metab. Dispos. 2008, 36, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, Z.; Vrzal, R.; Henklova, P.; Jancova, P.; Anzenbacherova, E.; Maurel, P.; Svecova, L.; Pavek, P.; Ehrmann, J.; Havlik, R.; et al. JNK inhibitor SP600125 is a partial agonist of human aryl hydrocarbon receptor and induces CYP1A1 and CYP1A2 genes in primary human hepatocytes. Biochem. Pharm. 2008, 75, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Zarybnicky, T.; Matouskova, P.; Lancosova, B.; Subrt, Z.; Skalova, L.; Bousova, I. Inter-Individual Variability in Acute Toxicity of R-Pulegone and R-Menthofuran in Human Liver Slices and Their Influence on miRNA Expression Changes in Comparison to Acetaminophen. Int. J. Mol. Sci. 2018, 19, 1805. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liver Sample | Sex (Age) | Reason of Surgery | Comorbidities | Long-Term Pharmacotherapy |
|---|---|---|---|---|
| L5 | male (63) | Colorectal carcinoma | HTN, hyperuricemia, type 2 DM | Ramipril, atorvastatin, metformin, allopurinol |
| L6 | male (69) | Colorectal carcinoma | HTN | Hydrochlorothiazide |
| L7 | male (69) | Colorectal carcinoma | HTN, s/p CVA | Acetylsalicylic acid, nitrendipine |
| L9 | male (81) | Colorectal carcinoma | HTN, dyslipidemia | Betaxolol |
| L11 | female (57) | Colorectal carcinoma | none | none |
| L14 | female (45) | Benign focal nodular hyperplasia | none | none |
| L16 | female (59) | Colorectal carcinoma | HLD, ovarian cancer | none |
| L19 | female (65) | Colorectal carcinoma | HTN, HLD, impaired glucose tolerance | Amlodipine |
| L36 | female (78) | Cholangiocellular carcinoma | HTN, HLD, coronary artery disease, atrial fibrillation | Bisoprolol, furosemide, ramipril, simvastatin, enoxaparin, zolpidem |
| L38 | male (59) | Cholangiocellular carcinoma | none | none |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| CYP3A4 | CCCCTGAAATTAAGCTTAGGAGG | CTGGTGTTCTCAGGCACAGA |
| CYP2C | TTTGGGATGGGGAAGAGGAG | GGAGCACAGCCCAGGAT |
| CBR1 | TTGGTACCCGAGATGTGTGC | CTTGGGGTTTTATTAGAGGGAG |
| AKR1C | ATGAGGAGCAGGTTGGACTG | GCTTTGAAGTGTAGAATATGTCTTCT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šadibolová, M.; Zárybnický, T.; Smutný, T.; Pávek, P.; Šubrt, Z.; Matoušková, P.; Skálová, L.; Boušová, I. Sesquiterpenes Are Agonists of the Pregnane X Receptor but Do Not Induce the Expression of Phase I Drug-Metabolizing Enzymes in the Human Liver. Int. J. Mol. Sci. 2019, 20, 4562. https://doi.org/10.3390/ijms20184562
Šadibolová M, Zárybnický T, Smutný T, Pávek P, Šubrt Z, Matoušková P, Skálová L, Boušová I. Sesquiterpenes Are Agonists of the Pregnane X Receptor but Do Not Induce the Expression of Phase I Drug-Metabolizing Enzymes in the Human Liver. International Journal of Molecular Sciences. 2019; 20(18):4562. https://doi.org/10.3390/ijms20184562
Chicago/Turabian StyleŠadibolová, Michaela, Tomáš Zárybnický, Tomáš Smutný, Petr Pávek, Zdeněk Šubrt, Petra Matoušková, Lenka Skálová, and Iva Boušová. 2019. "Sesquiterpenes Are Agonists of the Pregnane X Receptor but Do Not Induce the Expression of Phase I Drug-Metabolizing Enzymes in the Human Liver" International Journal of Molecular Sciences 20, no. 18: 4562. https://doi.org/10.3390/ijms20184562