Molecular Cloning and Functional Identification of a Squalene Synthase Encoding Gene from Alfalfa (Medicago sativa L.)



Abstract
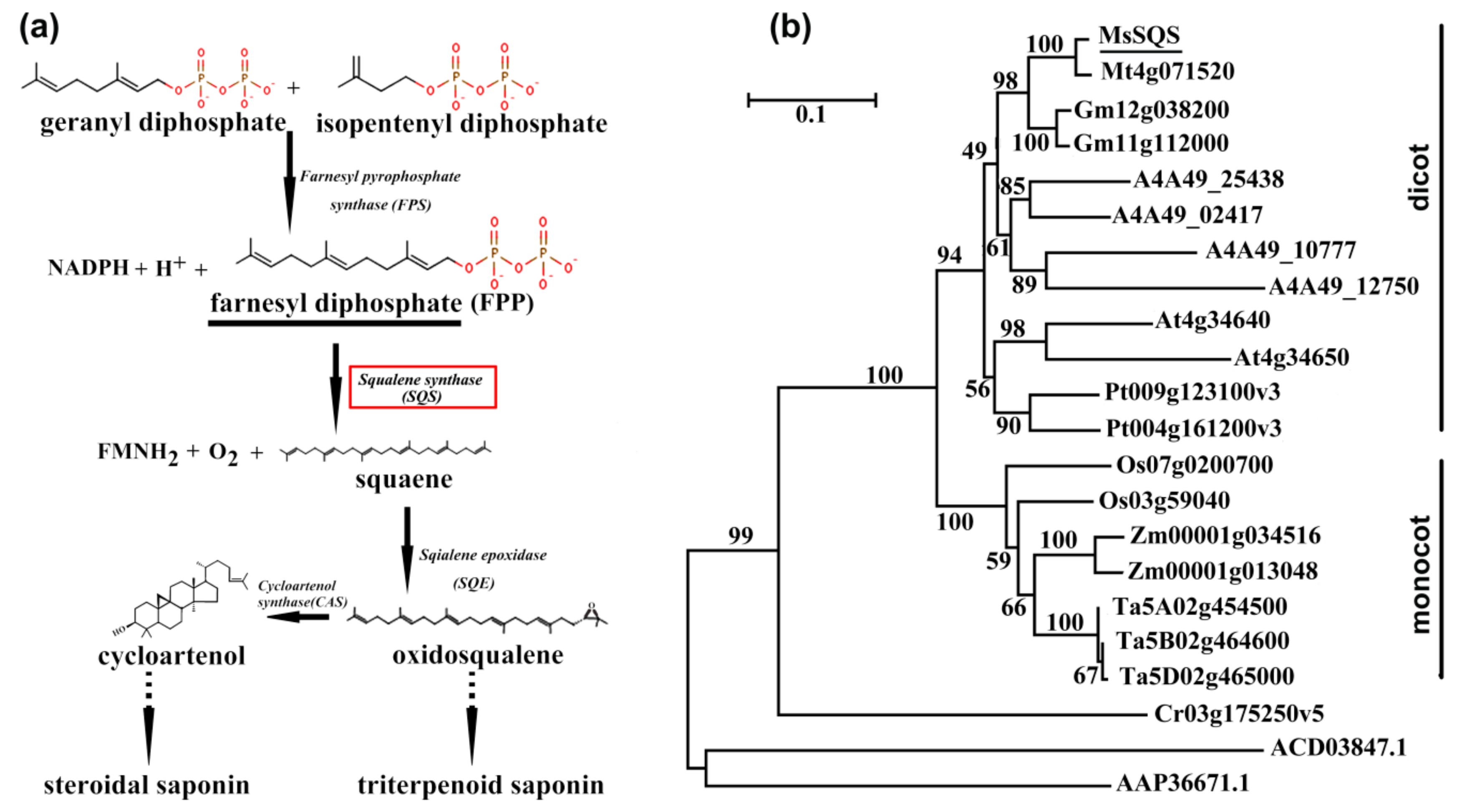
:1. Introduction
2. Results
2.1. MsSQS Encodes a Potential Squalene Synthase with High Sequence Identity to SQS Orthologs in Higher Plant
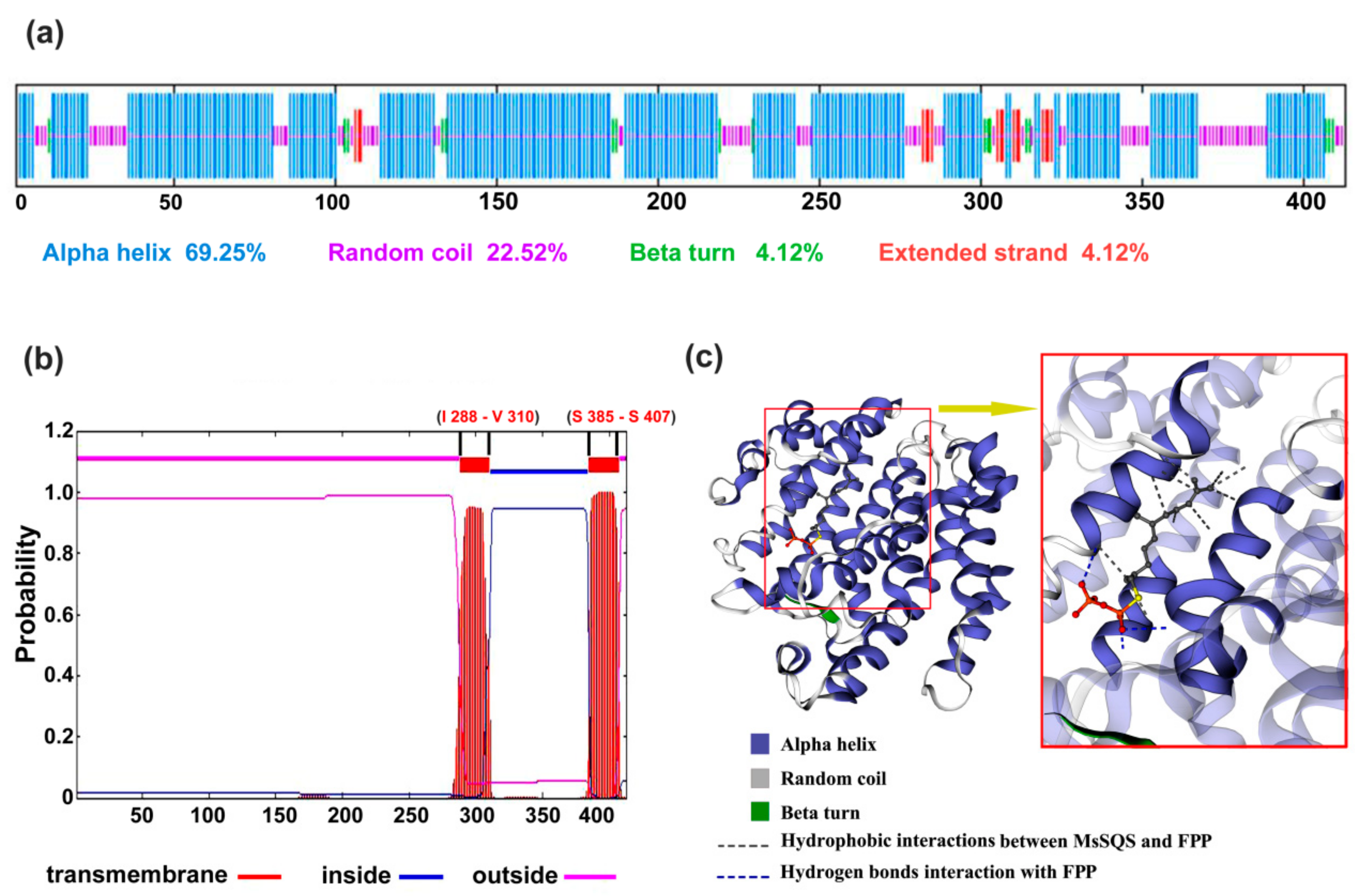
2.2. MsSQS Is a Canonical Squalene Synthase with the Common Features of SQS
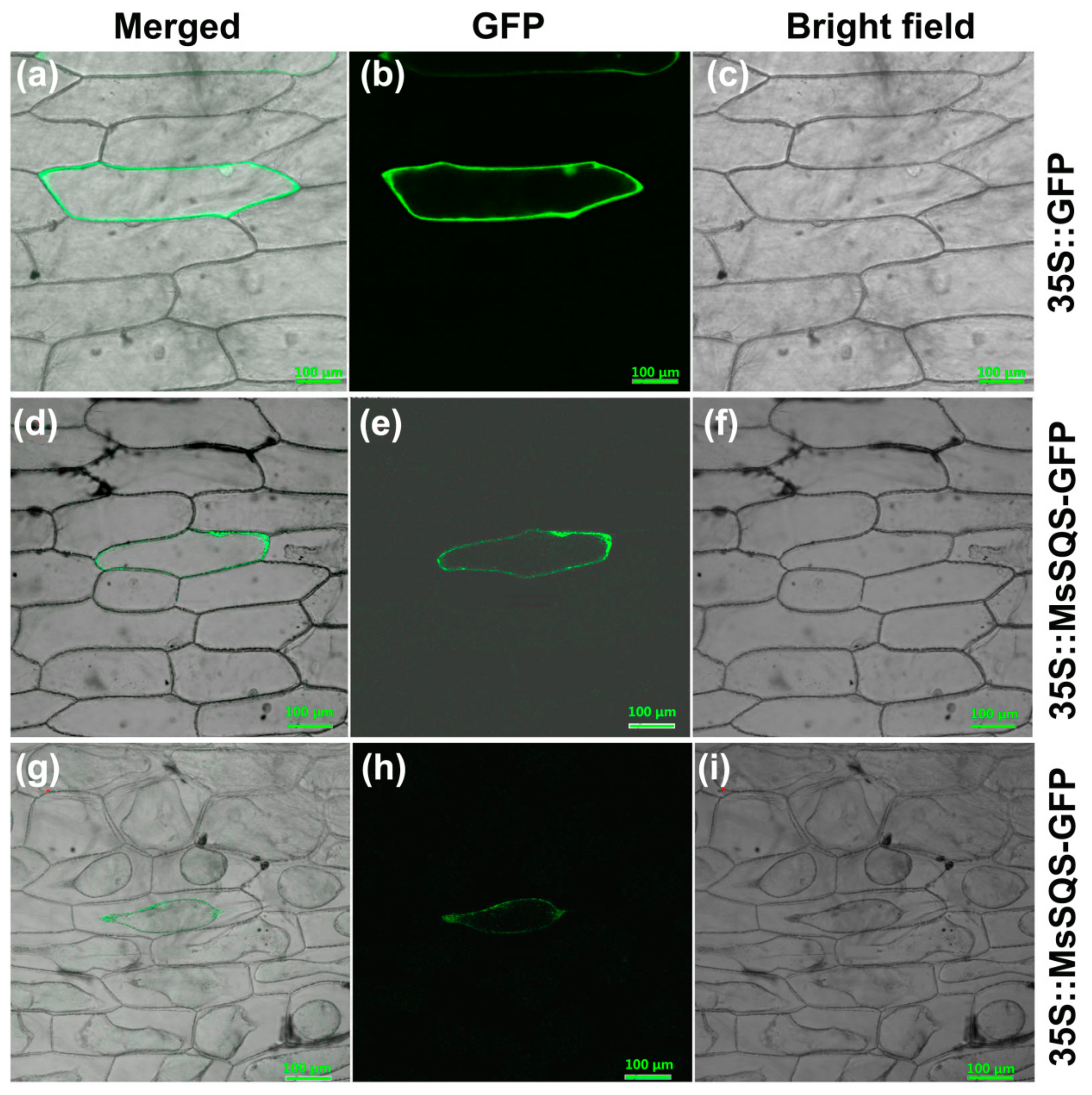
2.3. The MsSQS-GFP Recombinant Protein Resided Transiently on the Plasma Membrane of Onion Epidermal Cells
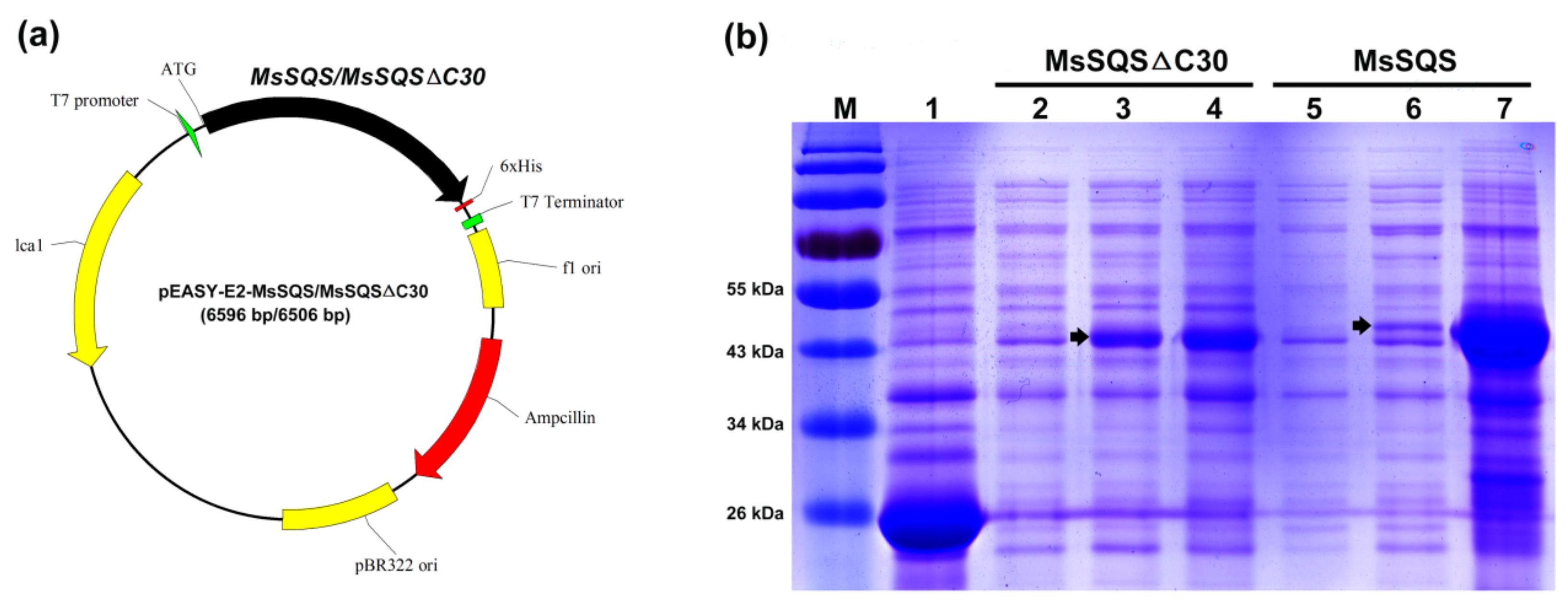
2.4. The Transmembrane Domain Affected the Solubility of MsSQS Expressed in Escherichia coli
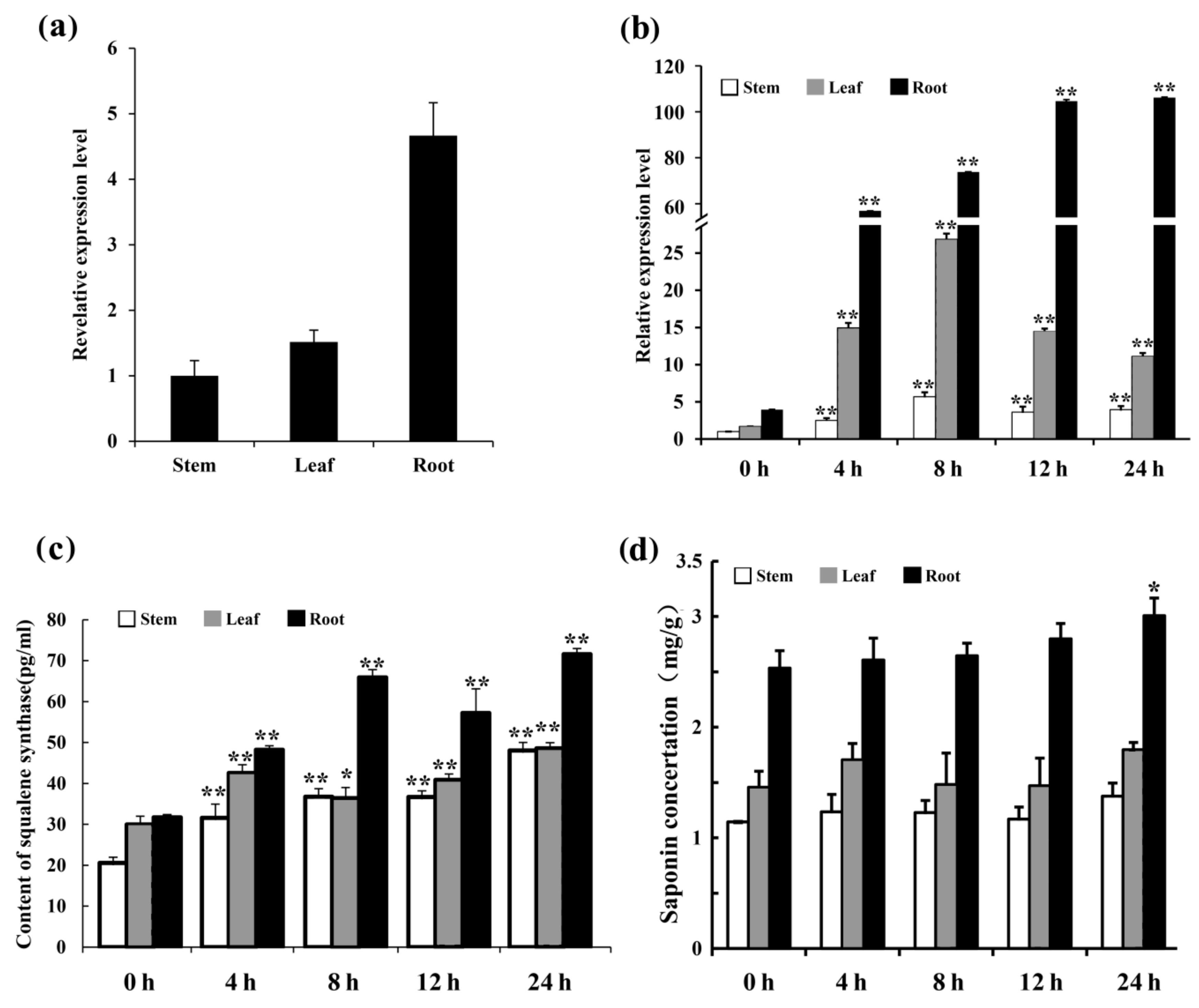
2.5. Expression Analysis of MsSQS in Alfalfa Tissues and under MeJA Treatment
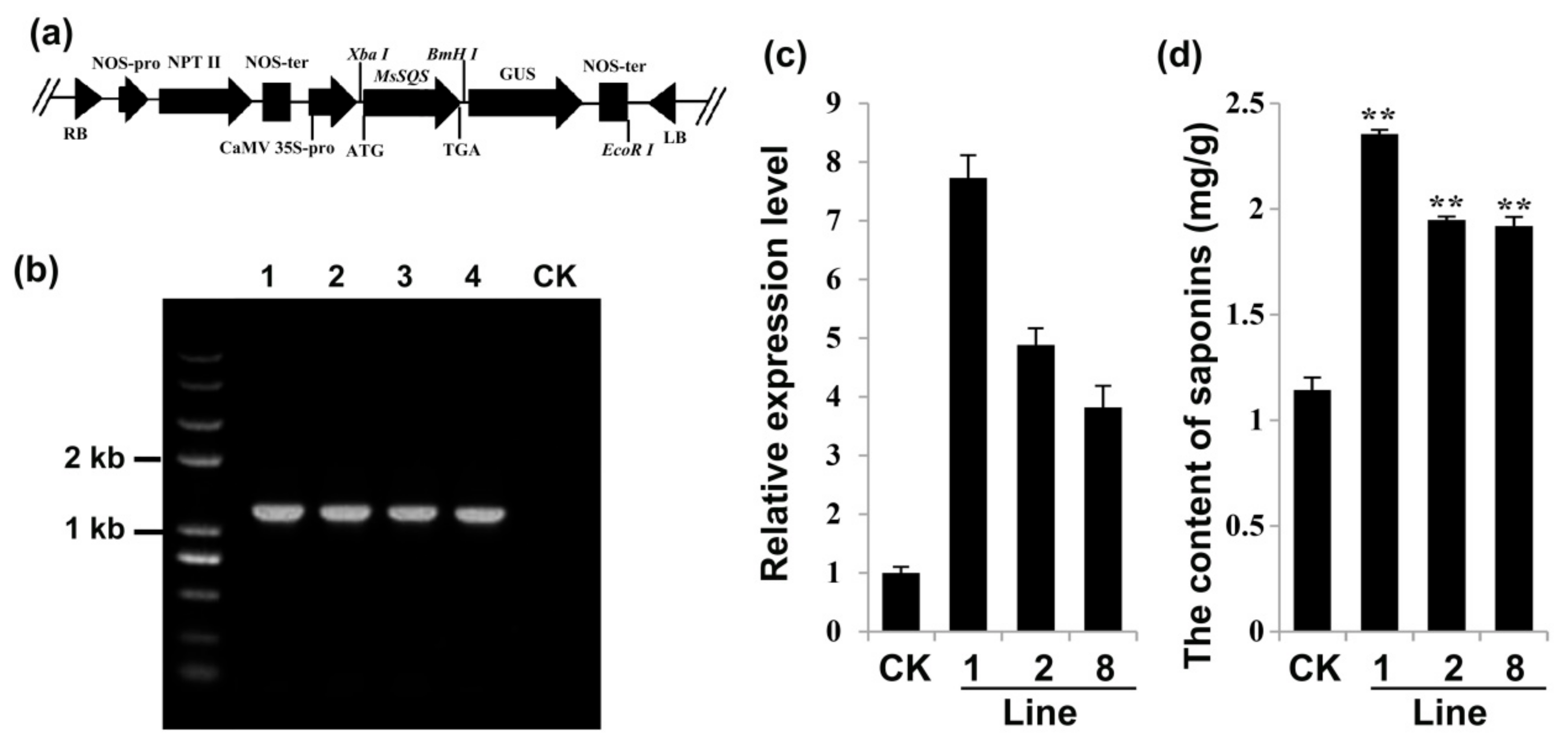
2.6. Overexpression of MsSQS Increased the Content of Saponins in Transgenic Alfalfa
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Plant Treatment
4.3. Cloning of MsSQS from Alfalfa and Expression Analysis by Quantitative Real-Time PCR
4.4. Bioinformatic Analysis of MsSQS
4.5. Constructions and Alfalfa Transformation
4.6. Protein Expression in Transient and Prokaryotic System
4.7. Measurement of the Content of Squalene Synthase Enzyme and Total Saponins
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Suzuki, H.; Achnine, L.; Xu, R.; Matsuda, S.P.; Dixon, R.A. A genomics approach to the early stages of triterpene saponin biosynthesis in medicago truncatula. Plant J. 2002, 32, 1033–1048. [Google Scholar] [CrossRef] [PubMed]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochem 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Aminfar, Z.; Tohidfar, M. In silico analysis of squalene synthase in fabaceae family using bioinformatics tools. J. Genet. Eng. Biotechnol. 2018, 16, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Linscott, K.B.; Niehaus, T.D.; Zhuang, X.; Bell, S.A.; Chappell, J. Mapping a kingdom-specific functional domain of squalene synthase. Biochim. Biophys. Acta 2016, 1861, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.W.; Tsay, Y.H.; Kienzle, B.K.; Smith-Monroy, C.A.; Bishop, R.W. Conservation between human and fungal squalene synthetases: Similarities in structure, function, and regulation. Mol. Cell Biol. 1993, 13, 2706–2717. [Google Scholar] [CrossRef]
- Akamine, S.; Nakamori, K.; Chechetka, S.A.; Banba, M.; Umehara, Y.; Kouchi, H.; Izui, K.; Hata, S. Cdna cloning, mrna expression, and mutational analysis of the squalene synthase gene of lotus japonicus. Biochim. Biophys. Acta 2003, 1626, 97–101. [Google Scholar] [CrossRef]
- Nakashima, T.; Inoue, T.; Oka, A.; Nishino, T.; Osumi, T.; Hata, S. Cloning, expression, and characterization of cdnas encoding arabidopsis thaliana squalene synthase. Proc. Natl. Acad. Sci. USA 1995, 92, 2328–2332. [Google Scholar] [CrossRef]
- Hata, S.; Sanmiya, K.; Kouchi, H.; Matsuoka, M.; Yamamoto, N.; Izui, K. Cdna cloning of squalene synthase genes from mono- and dicotyledonous plants, and expression of the gene in rice. Plant Cell Physiol. 1997, 38, 1409–1413. [Google Scholar] [CrossRef]
- Yousaf, A.; Qadir, A.; Anjum, T.; Ahmad, A. Transcriptional modulation of squalene synthase genes in barley treated with pgpr. Front. Plant Sci. 2015, 6, 672. [Google Scholar] [CrossRef]
- Ginzberg, I.; Thippeswamy, M.; Fogelman, E.; Demirel, U.; Mweetwa, A.M.; Tokuhisa, J.; Veilleux, R.E. Induction of potato steroidal glycoalkaloid biosynthetic pathway by overexpression of cdna encoding primary metabolism hmg-coa reductase and squalene synthase. Planta 2012, 235, 1341–1353. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Neelakadan, A.K.; Quach, T.N.; Valliyodan, B.; Kumar, R.; Zhang, Z.; Nguyen, H.T. Molecular characterization of glycine max squalene synthase genes in seed phytosterol biosynthesis. Plant Physiol. Biochem. 2013, 73, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Devarenne, T.P.; Shin, D.H.; Back, K.; Yin, S.; Chappell, J. Molecular characterization of tobacco squalene synthase and regulation in response to fungal elicitor. Arch. Biochem. Biophys. 1998, 349, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Zhao, C.L.; Chen, Z.J.; Wei, F.G.; Song, D.G.; Miao, K.R. Bioinformatics analysis of the squalene synthase gene and the amino acid sequence in ginseng species. Int. J. Clin. Exp. Med. 2015, 8, 12818–12825. [Google Scholar] [PubMed]
- Navarro Gallon, S.M.; Elejalde-Palmett, C.; Daudu, D.; Liesecke, F.; Jullien, F.; Papon, N.; Duge de Bernonville, T.; Courdavault, V.; Lanoue, A.; Oudin, A.; et al. Virus-induced gene silencing of the two squalene synthase isoforms of apple tree (malus x domestica l.) negatively impacts phytosterol biosynthesis, plastid pigmentation and leaf growth. Planta 2017, 246, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, S.; Yin, J.; Li, C.; Zhan, Y.; Xiao, J.; Liang, T.; Li, X. Molecular cloning and promoter analysis of squalene synthase and squalene epoxidase genes from betula platyphylla. Protoplasma 2016, 253, 1347–1363. [Google Scholar] [CrossRef]
- Grover, A.; Samuel, G.; Bisaria, V.S.; Sundar, D. Enhanced withanolide production by overexpression of squalene synthase in withania somnifera. J. Biosci. Bioeng. 2013, 115, 680–685. [Google Scholar] [CrossRef]
- Kim, Y.S.; Cho, J.H.; Park, S.; Han, J.Y.; Back, K.; Choi, Y.E. Gene regulation patterns in triterpene biosynthetic pathway driven by overexpression of squalene synthase and methyl jasmonate elicitation in bupleurum falcatum. Planta 2011, 233, 343–355. [Google Scholar] [CrossRef]
- Lee, M.H.; Jeong, J.H.; Seo, J.W.; Shin, C.G.; Kim, Y.S.; In, J.G.; Yang, D.C.; Yi, J.S.; Choi, Y.E. Enhanced triterpene and phytosterol biosynthesis in panax ginseng overexpressing squalene synthase gene. Plant Cell Physiol. 2004, 45, 976–984. [Google Scholar] [CrossRef]
- Seo, J.W.; Jeong, J.H.; Shin, C.G.; Lo, S.C.; Han, S.S.; Yu, K.W.; Harada, E.; Han, J.Y.; Choi, Y.E. Overexpression of squalene synthase in eleutherococcus senticosus increases phytosterol and triterpene accumulation. Phytochem 2005, 66, 869–877. [Google Scholar] [CrossRef]
- Uchida, H.; Yamashita, H.; Kajikawa, M.; Ohyama, K.; Nakayachi, O.; Sugiyama, R.; Yamato, K.T.; Muranaka, T.; Fukuzawa, H.; Takemura, M.; et al. Cloning and characterization of a squalene synthase gene from a petroleum plant, euphorbia tirucalli L. Planta 2009, 229, 1243–1252. [Google Scholar] [CrossRef]
- Jennings, S.M.; Tsay, Y.H.; Fisch, T.M.; Robinson, G.W. Molecular cloning and characterization of the yeast gene for squalene synthetase. Proc. Natl. Acad. Sci. USA 1991, 88, 6038–6042. [Google Scholar] [CrossRef] [PubMed]
- Busquets, A.; Keim, V.; Closa, M.; Del Arco, A.; Boronat, A.; Arro, M.; Ferrer, A. Arabidopsis thaliana contains a single gene encoding squalene synthase. Plant Mol. Biol. 2008, 67, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Wentzinger, L.F.; Bach, T.J.; Hartmann, M.A. Inhibition of squalene synthase and squalene epoxidase in tobacco cells triggers an up-regulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase. Plant Physiol. 2002, 130, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Oleszek, W. Alfalfa saponins: Structure, biological activity, and chemotaxonomy. Adv. Exp. Med. Biol. 1996, 405, 155–170. [Google Scholar] [PubMed]
- Liu, X.G.; Lv, M.C.; Huang, M.Y.; Sun, Y.Q.; Gao, P.Y.; Li, D.Q. A network pharmacology study on the triterpene saponins from medicago sativa l. For the treatment of neurodegenerative diseases. J. Food Biochem. 2019, 43, e12955. [Google Scholar] [CrossRef] [PubMed]
- Balestrazzi, A.; Carbonera, D.; Avato, P.; Tava, A. White poplar (populus alba l.) suspension cultures as a model system to study apoptosis induced by alfalfa saponins. Anticancer Agents Med. Chem. 2014, 14, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Cabral de Oliveira, A.C.; Perez, A.C.; Merino, G.; Prieto, J.G.; Alvarez, A.I. Protective effects of panax ginseng on muscle injury and inflammation after eccentric exercise. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2001, 130, 369–377. [Google Scholar] [CrossRef]
- Yoshiki, Y.; Okubo, K. Active oxygen scavenging activity of ddmp (2,3-dihydro-2,5-dihydroxy-6-methyl-4h-pyran-4-one) saponin in soybean seed. Biosci. Biotech. Biochem. 1995, 59, 1556–1557. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Abbruscato, P.; Tosi, S.; Crispino, L.; Biazzi, E.; Menin, B.; Picco, A.M.; Pecetti, L.; Avato, P.; Tava, A. Triterpenoid glycosides from medicago sativa as antifungal agents against pyricularia oryzae. J. Agric. Food Chem. 2014, 62, 11030–11036. [Google Scholar] [CrossRef]
- Mazahery-Laghab, H.; Yazdi-Samadi, B.; Bagheri, M.; Bagheri, A.R. Alfalfa (medicago sativa L.) shoot saponins: Identification and bio-activity by the assessment of aphid feeding. Br. J. Nutr. 2011, 105, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Gu, P.; Ishii, Y.; Spencer, T.A.; Shechter, I. Function-structure studies and identification of three enzyme domains involved in the catalytic activity in rat hepatic squalene synthase. J. Biol. Chem. 1998, 273, 12515–12525. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Long, R.; Zhang, T.; Wang, Z.; Zhang, F.; Yang, Q.; Kang, J.; Sun, Y. Molecular cloning and functional analysis of the drought tolerance gene mshsp70 from alfalfa (medicago sativa L.). J. Plant Res. 2017, 130, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Liu, Y.; Ma, C.; Chao, N.; Chen, Q.; Zhang, Y.; Luo, Y.; Cai, D.; Wu, Y. Positive selection of squalene synthase in cucurbitaceae plants. Int. J. Genomics 2019, 2019, 5913491. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, Y.; Chen, M.; Feng, J.; Ma, Z.; Zhang, X.; Zhu, C. Cloning, expression analysis and functional characterization of squalene synthase (sqs) from tripterygium wilfordii. Molecules 2018, 23, 269. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Fu, C.; Wang, Z.Y. A unified agrobacterium-mediated transformation protocol for alfalfa (medicago sativa L.) and medicago truncatula. Methods Mol. Biol. 2019, 1864, 153–163. [Google Scholar] [PubMed]
- Sparg, S.G.; Light, M.E.; Van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef]
- Ohana, P.; Delmer, D.P.; Carlson, R.W.; Glushka, J.; Azadi, P.; Bacic, T.; Benziman, M. Identification of a novel triterpenoid saponin from pisum sativum as a specific inhibitor of the diguanylate cyclase of acetobacter xylinum. Plant Cell Physiol. 1998, 39, 144–152. [Google Scholar] [CrossRef]
- Ellington, A.A.; Berhow, M.; Singletary, K.W. Induction of macroautophagy in human colon cancer cells by soybean b-group triterpenoid saponins. Carcinogenesis 2005, 26, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Fu, J. Squalene synthase cloning and functional identification in wintersweet plant (chimonanthus zhejiangensis). Bot. Stud. 2018, 59, 30. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Sharma, A. In silico study of binding motifs in squalene synthase enzyme of secondary metabolic pathway of solanaceae family. Mol. Biol. Rep. 2014, 41, 7201–7208. [Google Scholar]
- Su, H.; Liu, Y.; Xiao, Y.; Tan, Y.; Gu, Y.; Liang, B.; Huang, H.; Wu, Y. Molecular and biochemical characterization of squalene synthase from siraitia grosvenorii. Biotechnol. Lett. 2017, 39, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, L.N.; Pan, Y.T.; Kong, J.Q. Cdna isolation and functional characterization of squalene synthase gene from ornithogalum caudatum. Protein Expr. Purif. 2017, 130, 63–72. [Google Scholar] [CrossRef]
- Yin, X.M.; Bai, Z.C.; Niu, Y.Y.; Luo, H.M.; Chen, S.L. Cloning and analysis of squalene synthase (hssqs1) gene in huperzia serrata. Yao xue xue bao = Acta pharmaceutica Sinica 2012, 47, 1079–1084. [Google Scholar] [PubMed]
- Bhat, W.W.; Lattoo, S.K.; Razdan, S.; Dhar, N.; Rana, S.; Dhar, R.S.; Khan, S.; Vishwakarma, R.A. Molecular cloning, bacterial expression and promoter analysis of squalene synthase from withania somnifera (L.) dunal. Gene 2012, 499, 25–36. [Google Scholar] [CrossRef]
- Jiang, D.; Rong, Q.; Chen, Y.; Yuan, Q.; Shen, Y.; Guo, J.; Yang, Y.; Zha, L.; Wu, H.; Huang, L.; et al. Molecular cloning and functional analysis of squalene synthase (ss) in panax notoginseng. Int. J. Biol. Macromol. 2017, 95, 658–666. [Google Scholar] [CrossRef]
- Gupta, N.; Sharma, P.; Santosh Kumar, R.J.; Vishwakarma, R.K.; Khan, B.M. Functional characterization and differential expression studies of squalene synthase from withania somnifera. Mol. Biol. Rep. 2012, 39, 8803–8812. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Zhang, T.J.; Yang, Q.C.; Sun, Y.; Long, R.C.; Kang, J.M. Determination of the content of total saponins in different tissues of medicago sativa L. Chin. J. Grassland 2017, 39, 7–12. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Exon 1-0 a | Exon 1 | Exon 2 | Exon 3 | Exon 4 | Exon 5 | Exon 6 | Exon 7 | Exon 8 | Exon 9 | Exon 10 | Exon 11 | Exon 12 | Exon 13 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MsSQS | 216 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 245 | |
| Mt4g071520 | 572 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 766 | |
| At4g34640 | 495 | 43 | 90 | 76 | 70 | 150 b | 105 | 147 | 76 | 89 | 92 | 43 | 426 | |
| At3g34650 | 330 | 43 | 90 | 76 | 70 | 150 b | 105 | 147 | 76 | 89 | 92 | 43 | 345 | |
| Gm12g038200 | 421 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 452 | |
| Gm11g112000 | 453 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 467 | |
| A4A49_02417 | 407 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 610 | |
| A4A49_25438 | 305 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 70 | |
| A4A49_10777 | 257 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 86 | 28 | 70 | |
| A4A49_12750 | 243 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 43 | 67 | |
| Os03g0805100 | 231 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 40 | 107 | |
| Os07g0200700 | 310 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 40 | 251 | |
| Zm00001d013048 | 320 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 40 | 106 | |
| Zm00001d034516 | 336 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 40 | 109 | |
| Pt009g123100v3 | 408 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 923 | |
| Pt004g161200v3 | 37 | 347 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 46 | 431 |
| Ta5A02g454500 | 375 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 43 | 104 | |
| Ta5B02g464600 | 594 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 43 | 109 | |
| Ta5D02g465000 | 713 | 43 | 90 | 76 | 70 | 144 | 105 | 147 | 76 | 89 | 92 | 43 | 113 | |
| √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||||
| Cr03g175250v5 | 151 | 95 | 162 | 163 | 79 | 101 | 132 | 148 | 110 | 96 | 1208 | |||
| AAP36671.1 | 439 | 173 | 98 | 184 | 129 | 192 | 177 | 153 | 922 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.; Zhang, Q.; Jiang, X.; Zhang, T.; Long, R.; Yang, Q.; Wang, Z. Molecular Cloning and Functional Identification of a Squalene Synthase Encoding Gene from Alfalfa (Medicago sativa L.). Int. J. Mol. Sci. 2019, 20, 4499. https://doi.org/10.3390/ijms20184499
Kang J, Zhang Q, Jiang X, Zhang T, Long R, Yang Q, Wang Z. Molecular Cloning and Functional Identification of a Squalene Synthase Encoding Gene from Alfalfa (Medicago sativa L.). International Journal of Molecular Sciences. 2019; 20(18):4499. https://doi.org/10.3390/ijms20184499
Chicago/Turabian StyleKang, Junmei, Qiaoyan Zhang, Xu Jiang, Tiejun Zhang, Ruicai Long, Qingchuan Yang, and Zhen Wang. 2019. "Molecular Cloning and Functional Identification of a Squalene Synthase Encoding Gene from Alfalfa (Medicago sativa L.)" International Journal of Molecular Sciences 20, no. 18: 4499. https://doi.org/10.3390/ijms20184499
APA StyleKang, J., Zhang, Q., Jiang, X., Zhang, T., Long, R., Yang, Q., & Wang, Z. (2019). Molecular Cloning and Functional Identification of a Squalene Synthase Encoding Gene from Alfalfa (Medicago sativa L.). International Journal of Molecular Sciences, 20(18), 4499. https://doi.org/10.3390/ijms20184499