MicroRNA Networks Modulate Oxidative Stress in Cancer

Liver Research Center, Chang Gung Memorial Hospital, Linkou, Taoyuan 333, Taiwan
Int. J. Mol. Sci. 2019, 20(18), 4497; https://doi.org/10.3390/ijms20184497
Submission received: 20 August 2019
/
Revised: 6 September 2019
/
Accepted: 9 September 2019
/
Published: 11 September 2019
(This article belongs to the Special Issue Crosstalk between MicroRNA and Oxidative Stress in Physiology and Pathology)
Abstract
:Imbalanced regulation of reactive oxygen species (ROS) and antioxidant factors in cells is known as “oxidative stress (OS)”. OS regulates key cellular physiological responses through signal transduction, transcription factors and noncoding RNAs (ncRNAs). Increasing evidence indicates that continued OS can cause chronic inflammation, which in turn contributes to cardiovascular and neurological diseases and cancer development. MicroRNAs (miRNAs) are small ncRNAs that produce functional 18-25-nucleotide RNA molecules that play critical roles in the regulation of target gene expression by binding to complementary regions of the mRNA and regulating mRNA degradation or inhibiting translation. Furthermore, miRNAs function as either tumor suppressors or oncogenes in cancer. Dysregulated miRNAs reportedly modulate cancer hallmarks such as metastasis, angiogenesis, apoptosis and tumor growth. Notably, miRNAs are involved in ROS production or ROS-mediated function. Accordingly, investigating the interaction between ROS and miRNAs has become an important endeavor that is expected to aid in the development of effective treatment/prevention strategies for cancer. This review provides a summary of the essential properties and functional roles of known miRNAs associated with OS in cancers.
1. Introduction
Imbalanced regulation of reactive oxygen species (ROS) and antioxidant factors in cells is known as “oxidative stress (OS)” (Figure 1). OS drives key cellular physiological regulatory responses through signal transduction, transcription factors (TFs) and noncoding RNAs (ncRNAs) [1]. ROS are oxygen-containing products and are formed during cellular oxidative metabolism. ROS, including superoxide anion (O2−), hydroxyl radical (OH−), hydrogen peroxide (H2O2), nitric oxide (NO) and singlet oxygen (1O2), play important roles in cell differentiation, cell death, cell growth, signal transduction, cell apoptosis and chemoresistance [2,3]. Dual roles have been proposed for ROS in biological phenotypes according to their cellular level [4]. High levels of ROS promote cell apoptosis, while low levels of ROS act as a signal transducer to induce cell survival (Figure 1). Recently, excessive ROS production was identified in several cancers where they were significantly correlated with tumorigenesis. However, the underlying mechanism of ROS regulation in cancer development remains unclear.
MicroRNAs (miRNAs) are small ncRNA comprising 18-25-nucleotide functional RNA molecules that play critical roles in the regulation of target gene expression by binding to complementary regions of mRNA and regulating mRNA degradation or inhibiting translation (Figure 2). Previous studies have demonstrated that miRNAs are significantly associated with tumor growth, metastasis and cancer progression [5,6]. Based on these findings, dysregulated miRNA expression is a hallmark of cancer.
Cross-talk between ROS and miRNAs has been implicated in cancer development, and it is important to identify the nature of this connection. Interestingly, some specific miRNAs, called ROS-miRs or redoximiRs, are regulated by OS and modulate target gene expression in response to ROS [7,8]. Mesenguer et al. [9] demonstrated that the OS/NFκB axis induced miR-9/9* expression and inhibited expression of its target genes, GTPBP3, MTO1 and TRMU, in MELAS cells. On the other hand, a previous study indicated that miR-21 regulated ROS homeostasis and suppressed the antioxidant response in human umbilical vein endothelial cells (HUVECs) [10]. These findings suggest that ROS could be upstream regulators or downstream effectors of miRNAs. In this review, we focus on how ROS affect biological phenotypes through miRNA and how miRNAs regulate ROS-mediated function in cancer.
2. Regulation of ROS Homeostasis in Cells
OS promotes both nuclear and mitochondrial DNA damage and initiates DNA repair pathways [11]. Furthermore, cellular ROS levels can be produced by different mechanisms, such as ionizing radiation, UV radiation, inflammatory cells and chemotherapy. ROS are primarily generated in cells through the byproducts of leaked electrons from the mitochondrial electron transport chain (ETC). Mutations or aberrantly expressed nuclear or mitochondrial genes encoding the ETC components can influence the electron transfer reaction that leads to electron leakage. The electrons are captured by O2, producing O2−, which is usually converted to H2O2 by manganese (Mn)-containing mitochondrial superoxide dismutase (MnSOD or SOD2), Cu/Zn-containing cytosolic SOD1 or SOD3 [12]. Subsequently, H2O2 can attack chromosomal DNA and subsequently induce DNA damage. On the other hand, O2− can be generated through a reaction catalyzed by some enzymes, including the membrane-located NAD(P)H oxidase complex (NOX), which consists of NOX1-4, endoplasmic reticulum-associated xanthine oxidase (XO), cytochrome c oxidase and cyclooxygenase in some cancer cells [13]. In fact, H2O2 plays an important role in carcinogenesis because it is capable of diffusing throughout the cell components and producing cellular injury. The injurious effects of ROS in mammalian cells are mediated by the hydroxyl radical (·OH). The generation of OH in vivo is produced in the presence of reduced transition metals, including Co, Cu, Fe, or Ni, mainly through the Fenton reaction [14]. Notably, the ·OH-induced DNA damage includes the generation of 8-hydroxyguanosine (8-OHG), in which the hydrolysis product is 8-hydroxydeoxyguanosine (8-OHdG). 8-OHdG is the most widely used marker of radical attack on DNA. Notably, 8-OHdG is strongly correlated with cancer progression, including that of breast cancer, colorectal cancer, ovarian cancer and hepatocellular carcinoma (HCC) [15,16,17]. For example, hepatic 8-OHdG levels are useful biomarkers for identifying hepatitis C virus (HCV) infection in patients [18]. Alternatively, cells maintain ROS homeostasis by reducing ROS production and triggering specific antioxidant mechanisms to neutralize ROS or mitigate OS [19]. In fact, antioxidant enzymes include SODs, catalase, peroxiredoxins (PRDXs), thioredoxins, glutathione peroxidase and heme oxygenase. First, SOD converts O2− to O2 or H2O2. Then, catalase and glutathione peroxidase subsequently convert H2O2 to H2O and O2.
3. MiRNAs and Their Roles in Oxidative Stress
Previous studies indicated that ROS can induce or suppress miRNA expression and contribute to downstream biological function through regulation of target genes [20]. Increasing evidence has shown cross-talk between miRNAs and components of redox signaling [21,22]. The transcription, biogenesis, translocation and function of miRNAs are highly correlated with ROS, and miRNAs may regulate the expression of redox sensors and other ROS modulators, such as the key components of cellular antioxidant machinery. Redox sensors have been identified and they include transcription factors (e.g., p53, NFκB, c-Myc and nuclear factor erythroid 2 related factor 2 (NRF2)) and kinases (e.g., Akt and IKK), which trigger cellular redox signaling. Here, we summarize how miRNAs are regulated by ROS at the posttranscriptional and transcriptional levels and how the miRNA/ROS axis controls tumorigenesis.
3.1. MiRNA Processing is Regulated by ROS
Recently, it was reported that miRNAs can be transcribed by RNA polymerase II/III as longer primary transcripts called primary miRNAs (pri-miRNAs). The mature form of miRNA is generated by the two-step processing of pri-miRNA and is subsequently associated with the effector RNA-induced silencing complex (RISC). The biogenesis and function of miRNAs regulated by ROS are described. Two key genes (Dicer and Drosha) are mediated by the miRNA processing pathway (Figure 2). A report showed that the expression of Dicer was downregulated by aging-related OS in cerebromicrovascular endothelial cells (CMVECs) [23]. Downregulating Dicer dramatically reduced miRNA expression under H2O2 treatment compared with the expression of the control. Notably, knocking down Dicer suppressed ROS production in human microvascular endothelial cells (HMECs) [24]. These findings indicated that Dicer expression is part of a feedback loop that modulates ROS production and maintains cellular homeostasis. Upon OS, the expression of pre-miRNA and miRNA in myoblasts is decreased through DGCR8/heme oxygenase-1 (HMOX1) regulation [25]. Heme is required for DGCR8 activity, and the heme-binding domain of DGCR8 plays a crucial role in pri-miRNA recognition for miRNA processing by DROSHA.
3.2. ROS Regulate miRNA Expression through the Modulation of Transcription Factors
Accumulating studies have investigated the miRNAs regulated by ROS/TFs such as c-myc, p53, c-Jun, HIF and NFκB [20,26]. This section summarizes how miRNAs are regulated by ROS/TF at the transcriptional level.
ROS exposure has been shown to be correlated with oncogenic signals such as those transduced by c-Myc and Ras [27,28]. c-Myc, a well-known oncogene, is involved in tumor growth, migration, invasion, metabolism and metastasis through the regulation of gene expression. c-Myc activation induces DNA damage in normal human fibroblasts. This effect has been correlated with ROS generation. Expression levels of miR-15a/16, miR-23a, miR-29 and miR-34 family members were downregulated by c-Myc [29]. Overexpression of miR-15a/16 suppressed cell proliferation, angiogenesis, migration and invasion through inhibition of FGF2 in vitro and in vivo [30]. Furthermore, hypoxia-induced suppression of miR-15/16 expression was directly regulated by c-Myc. By contrast, miR-17-92 and miR-221/222 expression is stimulated by c-Myc [29]. The expression levels of miR-17-92 were remarkably inhibited by triptolide in a c-Myc-dependent manner, which resulted in the induction of target genes, including PTEN, BIM and p21, in HCC cells [31]. Moreover, this suppressive effect contributed to enhanced triptolide-induced cell apoptosis.
P53, a tumor suppressor gene, regulates the cell cycle, apoptosis, growth and metabolism through modulation of target genes. P53 is involved in regulating the drosha-dedicated pri-miRNA processing pathway [32]. In addition, p53 modulates miRNA transcription, such as miR-17-92, miR-34a and miR-200c. Interestingly, stress-regulated miRNAs, namely, miR-34 and miR-200, are upregulated in a p53-dependent manner [33,34]. MiR-34 has been implicated as a tumor suppressor because it suppresses the epithelial-mesenchymal transition (EMT), which promotes cancer cell metastasis. Its expression level is positively associated with p53. Importantly, p53 suppresses Snail expression by interacting with miR-34. A study indicates that miR-200c is upregulated upon H2O2 treatment in endothelial cells and that it contributes to cell apoptosis and senescence through inhibition of the target gene ZEB1 [35]. Moreover, knockdown of p53 can reverse H2O2-induced miR-200c expression [34].
As mentioned above, exposure to ROS induces chronic inflammation. NFκB acts as a master mediator of the inflammatory response to regulate innate and adaptive immune functions. MiRNA (miR-9, miR-21, miR-30b, miR-146a, miR-155 and miR-17-92 cluster) expression was identified and found to be directly transcriptionally regulated by NFκB [36,37]. Bazzoni et al. [38] indicated that miR-9-1 was induced by lipopolysaccharide (LPS) in a MyD88- and NF-κB-dependent manner. DNA damage activated miR-21 expression through recruitment of NF-κB and signal transducer and activator of transcription 3 (STAT3) to its promoter region and contributed to promoting cell invasion in breast cancer [39]. Another study reported that miR-21 expression and function were mediated by ROS in highly metastatic breast cancer cell lines [40]. In addition, miR-21 induced by ROS via NF-κB activity was involved in arsenic-induced cell transformation [41]. NF-κB bound to the promoter region of the miR-17-92 cluster was identified using chromatin immunoprecipitation (ChIP) assay and was further confirmed by luciferase reporter assay [42]. On the other hand, multiple miRNAs have been identified and have been found to modulate NF-κB activity. MiR-126a was shown to target IκBα, an NFκB inhibitor, and promoted the NFκB signaling pathway [43]. MiR-506 inhibited the expression of the NFκB p65 subunit and led to the production of ROS and p53-dependent apoptosis in lung cancer cells [44]. Notably, miR-506 was regulated by p53. These findings indicated that miR-506 was involved in the p53/NFκB signaling pathway.
NRF2 is a member of the Cap’n’Collar (CNC) family of basic leucine zipper (bZIP) transcription factors [45]. Previously, the actin-binding protein kelch-like ECH-associated protein 1 (KEAP1) was identified as a repressor of NRF2 via proteasomal degradation [46]. NRF2 is involved in antioxidant metabolism, protein degradation, inflammation and radioresistance [47]. Notably, miRNAs can be both indirectly and directly regulated by NRF2 [47,48]. Singh et al. [49] group demonstrated that NRF2 repressed miR-1 and miR-206 expression and led to reprogram glucose metabolism in cancer cells. Furthermore, miR-29 and miR-125b were identified as direct target genes of NRF2 [50,51]. Upregulation of miR-125b by NRF2 resulted in the repression of aryl hydrocarbon receptor repressor and protection of cancer cells from drug-induced toxicity [51]. On the other hand, NRF2 gene was regulated by miRNAs such as miR-28, miR-34a, miR-93 and miR-200a [52,53,54,55]. MiR-28 has been shown to interact with NRF2 3’UTR and represses NRF2 expression in breast cancer cells [52]. Overexpression of miR-34a suppressed NRF2 and NRF2 target genes expressions [53]. Functionally, miR-34a was involved in NRF2-dependent antioxidant pathway in liver. These findings suggested that NRF2 and miRNAs formed a regulatory network and regulated cellular functions.
3.3. ROS Regulate miRNA Expression via Epigenetic Regulation
Recently, epigenetic modifications/regulations of the genome have been explored and associated with cancer progression [56]. Changes in the structure or conformation at the nuclear or mitochondrial DNA (nDNA and mtDNA) or RNA level, but not the DNA/RNA sequence, are called epigenetic marks. The main epigenetic alterations in humans are DNA methylation and histone modification, which includes methylation, acetylation and phosphorylation. Aberrant miRNA expression in cancers was discovered and found to be controlled by epigenetic regulation. Promoter regions of miR-125b and miR-199a are hypermethylated through DNMT1 during H2O2 treatment, as determined using methylation-specific PCR and bisulfate sequencing [57]. Moreover, these two miRNAs are downregulated by ROS in ovarian cancer cells. The level of histone acetylation has an important role in activating gene expression through chromatin remodeling. In contrast, the gene is silenced by histone deacetylases (HDACs), which promote the deacetylation of lysine residues. MiR-466h-5p acts in a proapoptotic role by directly targeting antiapoptotic genes such as BCL2L2 [58]. ROS induce miR-466h-5p expression through inhibition of HDAC2 and result in increased apoptosis.
4. Interplay between Oxidative Stress, miRNA and Cancer Development
OS has been reported to contribute to neurological disorders, hypertension, diabetes and cancers. This section focuses on the associations of OS and hypoxia, angiogenesis, metastasis, metabolism, cancer stem cell and senescence, which are all involved in cancer progression.
4.1. Association between OS, miRNA and Hypoxia
Hypoxia, known as reduced oxygen availability, mostly occurs in the center of tumors due to the high proliferation ability of cancer cells and abnormal vasculature [59]. Hypoxia-inducible factor 1α (HIF1α) is the master regulator in hypoxia. The activation of HIF1α promotes the expression of several genes, including protein-encoding genes and ncRNAs, and facilitates stem cell renewal, cancer cell survival, metabolism and chemoresistance. The HIF1 transcription factor consists of three hypoxia-induced α subunits (HIF1α/2α/3α) and one β subunit (HIF1β). HIF1α is stabilized and activates a downstream signaling pathway mediated by ROS [60]. Some evidence suggests that telomerase activity is associated with ROS in HCC [61]. Moreover, ROS-mediated telomerase activity is dependent on HIF1α [62]. Expression levels of the human telomerase reverse transcriptase gene (hTERT) are upregulated by HIF1α. Specific binding sites for HIF1α in the hTERT promoter regions were identified by luciferase and ChIP assays. In addition, cancer stem cell (CSC) markers, OCT4 and Notch, are induced by HIF1α and promote stem cell renewal. Expression levels of SOX2 and KLF4 are positively regulated by ROS in glioblastoma cells [63]. HIF1α and ROS activation are responsible for regulating glucose transporter 1 (GLUT1), hexokinase II (HKII) and glutaminase expression and the reprogramming of cancer cell metabolism [64]. Moreover, miR-210 acts in an oncogenic role in cancer development and is induced under hypoxic conditions [65]. HIF1α directly binds to the hypoxia response element (HRE) of the miR-210 promoter. Therefore, miR-210 plays an important role in regulating cellular adaption to hypoxia, suggesting that targeting miR-210 may be a novel approach for the prevention and/or treatment of cancer.
4.2. Association between OS, miRNA and Angiogenesis
Angiogenesis is the process of generating new blood vessels from preexisting vasculature and is required for many functions, such as tissue repair, organ regeneration, cancer development and metastasis [66]. The angiogenesis process is regulated by several cytokines and growth factors, such as vascular endothelial growth factor (VEGF), transforming growth factor β (TGFβ), angiopoietin 1 (Ang-1) and placental growth factor, platelet-derived growth factor (PDGF) β [67,68,69,70]. VEGF acts as an effector to control endothelial cell proliferation and new vessel formation. The HIF1α/ROS axis activates tissue-specific angiogenesis through the upregulation of VEGF and its receptors VEGFR1 and VEGFR2. By contrast, VEGF induces ROS production by promoting NADPH oxidase in endothelial cells. ROS can also modulate VEGFR activation, phosphorylation and polymerization. A report indicated that genotoxic stress-induced miR-494 expression suppressed DNA repair and angiogenesis through regulation of MRE11a/RAD50/NBN (MRN) complex in endothelial cells [71]. Moreover, VEGF signaling is regulated by MRN complex in vitro and in vivo. Alternatively, ROS stimulate the MAPK pathway and promote the expression of VEGF. A previous study demonstrated that oxidized phospholipids interact with VEGFR2 and induce angiogenesis through the Src signaling pathway [72]. Other mechanisms of ROS-mediated angiogenesis are the ataxia telangiectasia mutated gene (ATM)/p38α pathway and Sirtuin 1 (SIRT1). Previous studies have indicated that ATM functions in the cell cycle regulation, DNA damage repair and oxidative defense [73]. ATM promotes endothelial cell proliferation and facilitates angiogenesis [74]. Previously, the subtype of histone H2A, called H2AX, can be phosphorylated (γH2AX) and is involved in DNA damage response. Economopoulou et al. [75] group indicated that H2AX is required for endothelial cells to sustain their growth under hypoxia and is important for hypoxia-driven neovascularization. Wilson et al. [76] have shown that miR-103 suppresses developmental and pathological angiogenesis through inhibition of three prime exonucleases 1 in endothelial cells. On the other hand, Yang and co-workers demonstrated that overexpression of miR-328-3p suppressed cell proliferation and promoted radiosensitivity of osteosarcoma cells through suppression of H2AX in vitro and in vivo [77]. Recently, Marampon et al. group demonstrated that NRF2/antioxidant enzymes/H2AX/miRNAs (miR-22, miR-34a, miR-126, miR-146a, miR-210 and miR-375) axis act as potential candidates in radiosensitizing therapeutic strategy for rhabdomyosarcoma clinical treatment [78]. SIRT1, also known as NAD-dependent deacetylase sirtuin-1, has been demonstrated to regulate cellular functions including oxidative stress, apoptosis and aging via deacetylation of a variety of substrates [79]. A report indicates that inhibition of SIRT1 with either an inhibitor or siRNA leads to increased ROS levels, suggesting an association between SIRT1 and ROS. MiR-138, miR-181 and miR-199 have been shown to directly target and inhibit SIRT1 expression in various cell lines [80,81,82]. MiR-181 is induced by treatment with a high-fat diet and results in repressed SIRT1 expression and insulin sensitivity in the liver [81]. In addition, HIF1α and SIRT1 are upregulated in miR-199a-depleted cells during normoxic conditions [83]. Moreover, SIRT1 is actually a direct target gene of miR-199a and is responsible for suppressing prolyl hydroxylase 2.
4.3. Association between OS, miRNA and Metastasis
Metastasis is a complicated process that includes invasion, intravasation into blood, extravasation to distant organs and growth [84]. Due to these multiple steps, few metastasizing tumor cells can survive and form micrometastases. A typical phenotype that leads to metastasis is EMT, which is a biological event by which epithelial cells undergo alterations that induce the development of a more aggressive mesenchymal phenotype [84]. Increasing evidence suggests that cancer cells during the metastasis process are killed by OS [85,86]. In addition, cancer cells are more sensitive to ROS than normal cells. Reducing ROS levels by treating with antioxidant inhibits tumor promotion of tumor progression in mouse models. ROS-mediated EMT regulation through TGFβ/Smad, E-cadherin, Snail, integrin, β-catenin, matrix metalloproteinases (MMPs) and miRNA has been documented [87,88,89]. Among these interactions, activation of TGFβ induces ROS production and leads to the promotion of SMAD and ERK1/2 phosphorylation. Moreover, the ROS/TGFβ axis regulates EMT through the interaction of NFκB, HIF1α and cyclooxygenase-2 (COX-2). A previous study indicated that MMP-3, MMP-10 and MMP-13 were directly upregulated by oxidative treatment and promoted cell invasion ability in NMuMG cells [90]. In addition, the activity of MMP-2 and MMP-9 were posttranscriptionally regulated by oxidant treatment [91,92]. These studies suggest that MMP expression or activity is modulated by OS, which is related to chronic inflammation, malignant transformation and the invasive potential of cells. Yoon et al. [93] demonstrated that sustained treatment with H2O2 enhances MMP2 activity via the PDGF, VEGF, phosphatidylinositol 3-kinase and NF-κB pathways in HT1080 cell lines. Song and coworkers reported that the expression of miR-509 is significantly more downregulated in breast cancer than it is in normal tissues [94]. Overexpressed miR-509 abrogated cell growth, migration and invasion through inhibition of the target gene SOD2, which is a crucial effector in the production of ROS.
4.4. Association between OS, miRNA and Metabolism
Tumor progression is characterized by the occurrence of metabolic alterations, including those in glycolysis, fatty acid oxidation (FAO) and oxidative phosphorylation [95,96]. The connection and reciprocal regulation between the metabolism and the redox balance of tumor cells have been shown. For this reason, it is important to determine the major metabolic pathways that are the main controllers of the ROS homeostasis of cancer cells. Glucose is converted to glucose-6-phosphate by hexokinase enzyme and triggers a series of downstream enzyme-catalyzed reactions. It is an essential pathway for providing nutrients, metabolites and energy to cells. In 1924, Otto Warburg proposed a theory suggesting that tumor cells tend to exhibit glycolysis regardless of the presence of oxygen [97]. Accumulating evidence has shown that metabolites produced by glucose metabolism are major regulators of the redox homeostasis of tumor cells [98]. Cancer cells demonstrate increased sensitivity to glucose-deprivation-induced cytotoxicity compared with that in normal cells by restricting the burden of ROS. Moreover, inhibition of lactate dehydrogenase-A by a specific inhibitor, FX11, reduced intracellular ATP and promoted OS, which suppressed tumor progression in lymphoma and pancreatic cancer. Sala et al. [10] have shown that miR-21 is upregulated by glucose treatment and inhibits ROS homeostatic genes such as NRF2, SOD2 and KRIT1. Furthermore, other metabolic enzymes, such as TIGAR and ALDH4, decrease ROS production by either inhibiting glycolysis and inducing NAPDH production or enhancing mitochondrial function [99,100]. FAO consists of multiple processes by which fatty acids are broken down by cells to produce ATP and generate biosynthetic pathways. In general, the β-oxidation reaction takes place in mitochondria. FAO causes ROS formation and contributes to the enhanced development of nonalcoholic fatty liver disease (NAFLD) [101]. In hypoxia, HIF-1 suppression of medium-chain acyl-CoA dehydrogenase (MCAD) and light chain acyl-CoA dehydrogenase (LCAD) expression inhibits FAO and ROS production while promoting cell growth of liver cancer cells [102]. The expression levels of LCAD in HCC specimens were analyzed and found to be negatively correlated with survival. These findings indicate the relevance of FAO suppression in the progression of cancer. Previous studies have identified miR-33a/b as an intronic miRNA located with the sterol regulatory element binding factor (SREBP) 1 and 2 genes [103]. These two miRNAs cotranscribe with their host gene and regulate high density lipoprotein (HDL) biosynthesis.
4.5. Association between OS, miRNA and Cancer Stem Cells
Cancer cells are believed to be derived from a small subset of tumor cells that have a high capacity for self-renewal and differentiation—namely, cancer stem cells (CSCs) or tumor-initiating cells [104]. Increasing evidence indicates that miRNAs function as regulators of CSCs and are associated with ROS production during tumor progression and cancer development. Some miRNAs, such as let-7a, miR-21, miR-34a, miR-200 and miR-210, are potentially involved in the modulation of ROS production in CSCs [105,106,107,108,109]. A previous study showed that let-7 acts as a negative regulator of CSC-mediated function by targeting PTEN and LIN28b in prostate and pancreatic cancer. Recently, OS reduced let-7 expression in a p53-dependent manner in various cancer cells. Some experimental studies revealed that the expression of miR-21 is remarkably increased in CSC subpopulations compared to the expression in the hypobromite non-CSC counterparts in vitro and in vivo. Notably, knocking down miR-21 suppressed cell migration, invasion and EMT phenotype in breast cancer CSCs. Moreover, OS induced miR-21 expression and promoted cell migration and self-renewal in prostate and pancreatic CSCs. Another report indicates that miR-21 enhances ROS production via the MAPK pathway and suppresses SOD2, SOD3 and sprouty homolog 2 (SPRY-2) expression [110]. Additionally, a number of studies have revealed that miR-34a suppresses CSC-related genes, such as CD44, and EMT makers and subsequently attenuates cell invasion, metastasis and self-renewal capacity [111]. The interplay between ROS and miR-34 has been documented. The expression of miR-34 is induced by OS in stromal and tumor cells. The first evidence miR-200 was associated with stem cell phenotype, reported in 2009 [112]. Moreover, all five members of the miR-200 family were downregulated in human breast CSCs as well as in normal human and murine mammary stem/progenitor cells [112]. Mechanistically, miR-200 suppresses the expression of B lymphoma Mo-MLV insertion region 1 homolog (Bmil-1), Suz12, and Notch homolog 1 (Notch1), which are known regulators of CSC and EMT phenotypes, and inhibits the CSC self-renewal capacity. MiR-210 expression is enriched in MCF-7 spheroid cells and CD44+/CD24− MCF7 cells compared with MCF-7 parental cells [113]. Overexpression of miR-210 enhances proliferation, self-renewal capacity, migration and invasion through inhibition of E-cadherin in vitro and in vivo. Thus, these observations indicate that the miRNA/ROS axis plays important roles in multiple events related to CSCs.
4.6. Association between OS, miRNA and Senescence
Cellular senescence is characterized by the expression of senescence-associated β-galactosidase (SA-β-gal), overexpression of the cyclin-dependent kinase (CDK) inhibitor, senescence-associated secretory phenotype (SASP), telomere shortening and persistent DNA damage response (DDR) [114]. ROS cause cell senescence by stimulating the DDR pathway to stabilize p53 and promote CDK inhibitor gene expression. In fact, p53 acts as a master regulator in the cellular response to OS. Mechanically, p53 can decrease ROS levels and repair DNA damage in cells. In contrast, it can also enhance ROS production and promote cell apoptosis or senescence [115]. Several reports indicate that p53 reduces intracellular ROS levels by promoting antioxidant reactions. Several miRNAs, including miR-21, miR-22, miR-29, miR-34a, miR-106b, miR-125b, miR-126, miR-146a, the miR-17-92 cluster, the miR-200 family and miR-210, have been identified to be differentially expressed in senescent cells and to be involved in cellular senescence [116,117,118,119,120,121,122]. Notably, miR-34a was found to promote cellular senescence by inhibiting SIRT1 expression in a variety of tissues. Another group indicated that miR-34a and miR-335 promote premature cellular senescence by targeting antioxidative enzymes. Furthermore, miR-217 induces a premature senescence-like phenotype and represses angiogenesis by inhibiting the expression of target gene SIRT1 in endothelial cells [123]. In addition, miR-92a was found to exacerbate endothelial dysfunction under OS exposure by directly targeting SIRT1, Krüppel-like factor 2 (KLF2) and KLF4 genes [124]. Additionally, Liu et al. group demonstrated that knockdown of miR-92a promoted cell growth, decreased caspase 3 activity and ROS through regulation of NRF2-KEAP1/ARE signal pathway [125].
5. ROS-Mediated Therapeutic Strategies in Cancer
OS clearly plays a role in the development of cancer, metastasis and chemotherapeutic resistance. In a strategy to modulate ROS-mediated effects, these biochemical characteristics of tumors are directly impaired. In light of recent studies, the strategy of inhibiting metabolic pathways, targeting NADPH oxidase and ROS scavenging mechanisms represent promising therapeutic options for treatments [126]. The other strategy is to target tumor cells with oxidation-promoting agents that either enhance ROS production or inhibit cellular antioxidants. NADPH oxidase plays an important role in regulating ROS production. Several inhibitors have been demonstrated to reduce NADPH function. In general, diphenylene iodonium (DPI) and apocynin are NADPH inhibitors [127,128,129]. DPI can inhibit XOD and the proteins of mitochondrial ETC and block flavoprotein. In another strategy, ROS scavenging enzymes are enhanced and used for anticancer therapy [130]. GSH, GST, SOD, GPX, and catalase are able to suppress tumor formation. There are several analogs of GSH drugs, such as N-acetylcysteine (NAC), YM737 and Telcyta, used for cancer treatment [131]. NOV-002, an agent containing oxidized GSH, improved the efficacy of cyclophosphamide to treat colon cancer by controlling the ratio of GSH to GSSG and promoting S-glutathionylation [131]. Yang and coworkers indicated that lithocholic acid treatment and bile duct ligation model promoted c-Myc/miR-27/prohibitin 1 axis, with the consequence of repressing NRF2 expression and ARE binding, resulting in decreased suppressed GSH synthesis and antioxidant ability in chronic cholestatic liver injury [132]. Another study reported that the rate-limiting GSH biosynthetic heterodimeric enzyme γ-glutamyl-cysteine ligase (GCL) was regulated by miR-433 [133]. Ectopic of miR-433 in HUVEC inhibited GCL expression in an NRF2-independent manner. Moreover, inhibition of miR-433 prevented TGFβ-mediated GCL downregulation and fibrogenesis in hepatic cells. Recently, Cheng et al. [134] demonstrated that miR-30e expression was suppressed in an atherosclerosis (AS) model. MiR-30e regulates Snai1/TGF-β/Nox4 expression to modulate ROS. These findings provide novel insights on miRNAs in the anti-ROS pathway, in which miRNA-30e may represent a novel target for AS.
6. Conclusions
Overall, many studies have been conducted to elucidate the molecular mechanisms underlying the ROS/miRNA axis and its role in tumorigenesis (Figure 3). Moreover, miRNAs networks that modulate OS in cancer are comprehensively listed in Table 1. Indeed, ROS and miRNAs exhibit overlapping characteristics in tumorigenesis. ROS, as upstream regulators, modulate miRNA expression through transcriptional, posttranscriptional and epigenetic regulation, respectively. On the other hand, miRNAs disrupt ROS production (downstream mediator) and are involved in ROS-mediated functions. MiRNAs and ROS can act either synergistically or antagonistically to regulate cancer progression. However, many details of their interaction remain unclear and need to be further investigated. MiRNAs/ROS-mediated phenotypes depend on the net result of the downstream molecules and multiple signaling pathways in the specific context. There are still many limitations to treatment because ROS play dual roles in cancer progression. As discussed in this review, the functional roles of miRNA in cellular adaptation to ROS are different in cells based on tissue and cell-type specific effects. These observations raise the possibilities to apply specific miRNAs as therapeutic targets in different contexts. Advantages of using miRNA-target therapy include the conservation of miRNA across multiple species with known sequences and the ability to target multiple genes within defined pathways. Notably, several miRNA-based therapies are being developed. For example, the locked nucleic acid (LNA)-modified anti-miR-122 is the first miRNA-targeted therapy to treat HCV in clinical trials. The association between ROS-mediated function and miRNA regulation provides opportunities for developing novel anticancer strategies.
Funding
This work was supported by grants from Chang Gung Memorial Hospital, Taoyuan, Taiwan (CMRPG3H0721, CMRPG3H0722, NZRPG3G0171, NZRPG3G0172, NZRPG3G0173, NMRPG3H0561, NRRPG3J0141 to YHL) and the Ministry of Science and Technology of the Republic of China (MOST 106-2321-B-182A-004-MY3, MOST 107-2320-B-182A-028-, MOST 108-2320-B-182A-004- to YHL).
Conflicts of Interest
The author has no conflict of interest to disclose.
References
- Dong, Y.; Xu, W.; Liu, C.; Liu, P.; Li, P.; Wang, K. Reactive oxygen species related noncoding RNAs as regulators of cardiovascular diseases. Int. J. Biol. Sci. 2019, 15, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Engedal, N.; Zerovnik, E.; Rudov, A.; Galli, F.; Olivieri, F.; Procopio, A.D.; Rippo, M.R.; Monsurro, V.; Betti, M.; Albertini, M.C. From oxidative stress damage to pathways, networks, and autophagy via microRNAs. Oxid. Med. Cell. Longev. 2018, 2018, 4968321. [Google Scholar] [CrossRef] [PubMed]
- Castaneda-Arriaga, R.; Perez-Gonzalez, A.; Reina, M.; Alvarez-Idaboy, J.R.; Galano, A. Comprehensive investigation of the antioxidant and pro-oxidant effects of phenolic compounds: A double-edged sword in the context of oxidative stress? J. Phys. Chem. B 2018, 122, 6198–6214. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. (Lausanne) 2018, 9, 402. [Google Scholar] [CrossRef]
- Tan, W.; Liu, B.; Qu, S.; Liang, G.; Luo, W.; Gong, C. MicroRNAs and cancer: Key paradigms in molecular therapy. Oncol. Lett. 2018, 15, 2735–2742. [Google Scholar] [CrossRef]
- Liu, Y.; Qiang, W.; Xu, X.; Dong, R.; Karst, A.M.; Liu, Z.; Kong, B.; Drapkin, R.I.; Wei, J.J. Role of miR-182 in response to oxidative stress in the cell fate of human fallopian tube epithelial cells. Oncotarget 2015, 6, 38983–38998. [Google Scholar] [CrossRef]
- Fierro-Fernandez, M.; Miguel, V.; Lamas, S. Role of redoximiRs in fibrogenesis. Redox Biol. 2016, 7, 58–67. [Google Scholar] [CrossRef]
- Meseguer, S.; Martinez-Zamora, A.; Garcia-Arumi, E.; Andreu, A.L.; Armengod, M.E. The ROS-sensitive microRNA-9/9* controls the expression of mitochondrial tRNA-modifying enzymes and is involved in the molecular mechanism of MELAS syndrome. Hum. Mol. Genet. 2015, 24, 167–184. [Google Scholar] [CrossRef]
- La Sala, L.; Mrakic-Sposta, S.; Micheloni, S.; Prattichizzo, F.; Ceriello, A. Glucose-sensing microRNA-21 disrupts ROS homeostasis and impairs antioxidant responses in cellular glucose variability. Cardiovasc. Diabetol. 2018, 17, 105. [Google Scholar] [CrossRef]
- Saki, M.; Prakash, A. DNA damage related crosstalk between the nucleus and mitochondria. Free Radic. Biol. Med. 2017, 107, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Case, A.J. On the origin of superoxide dismutase: An evolutionary perspective of superoxide-mediated redox signaling. Antioxidants (Basel) 2017, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Meitzler, J.L.; Antony, S.; Wu, Y.; Juhasz, A.; Liu, H.; Jiang, G.; Lu, J.; Roy, K.; Doroshow, J.H. Nadph oxidases: A perspective on reactive oxygen species production in tumor biology. Antioxid. Redox Signal. 2014, 20, 2873–2889. [Google Scholar] [CrossRef] [PubMed]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [PubMed]
- Pylvas, M.; Puistola, U.; Laatio, L.; Kauppila, S.; Karihtala, P. Elevated serum 8-OHdG is associated with poor prognosis in epithelial ovarian cancer. Anticancer Res. 2011, 31, 1411–1415. [Google Scholar] [PubMed]
- Plachetka, A.; Adamek, B.; Strzelczyk, J.K.; Krakowczyk, L.; Migula, P.; Nowak, P.; Wiczkowski, A. 8-hydroxy-2’-deoxyguanosine in colorectal adenocarcinoma--is it a result of oxidative stress? Med. Sci. Monit. 2013, 19, 690–695. [Google Scholar] [PubMed]
- Ma-On, C.; Sanpavat, A.; Whongsiri, P.; Suwannasin, S.; Hirankarn, N.; Tangkijvanich, P.; Boonla, C. Oxidative stress indicated by elevated expression of NRF2 and 8-OHdG promotes hepatocellular carcinoma progression. Med. Oncol. 2017, 34, 57. [Google Scholar] [CrossRef]
- Tanaka, H.; Fujita, N.; Sugimoto, R.; Urawa, N.; Horiike, S.; Kobayashi, Y.; Iwasa, M.; Ma, N.; Kawanishi, S.; Watanabe, S.; et al. Hepatic oxidative DNA damage is associated with increased risk for hepatocellular carcinoma in chronic hepatitis c. Br. J. Cancer 2008, 98, 580–586. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Ahamed, M.; Alhadlaq, H.A.; Alshamsan, A. Mechanism of ROS scavenging and antioxidant signalling by redox metallic and fullerene nanomaterials: Potential implications in ROS associated degenerative disorders. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 802–813. [Google Scholar] [CrossRef]
- He, J.; Jiang, B.H. Interplay between reactive oxygen species and microRNAs in cancer. Curr. Pharmacol. Rep. 2016, 2, 82–90. [Google Scholar] [CrossRef]
- Gong, Y.Y.; Luo, J.Y.; Wang, L.; Huang, Y. MicroRNAs regulating reactive oxygen species in cardiovascular diseases. Antioxid. Redox Signal. 2018, 29, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Huang, Z.; Han, J.; Shao, J.; Huang, C. Redox regulation of microRNAs in cancer. Cancer Lett. 2018, 418, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tucsek, Z.; Sosnowska, D.; Toth, P.; Gautam, T.; Podlutsky, A.; Csiszar, A.; Losonczy, G.; Valcarcel-Ares, M.N.; Sonntag, W.E.; et al. Aging-induced dysregulation of dicer1-dependent microRNA expression impairs angiogenic capacity of rat cerebromicrovascular endothelial cells. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Shilo, S.; Roy, S.; Khanna, S.; Sen, C.K. Evidence for the involvement of miRNA in redox regulated angiogenic response of human microvascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Kozakowska, M.; Ciesla, M.; Stefanska, A.; Skrzypek, K.; Was, H.; Jazwa, A.; Grochot-Przeczek, A.; Kotlinowski, J.; Szymula, A.; Bartelik, A.; et al. Heme oxygenase-1 inhibits myoblast differentiation by targeting myomirs. Antioxid. Redox Signal. 2012, 16, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Markopoulos, G.S.; Roupakia, E.; Tokamani, M.; Alabasi, G.; Sandaltzopoulos, R.; Marcu, K.B.; Kolettas, E. Roles of NF-kB signaling in the regulation of miRNAs impacting on inflammation in cancer. Biomedicines 2018, 6, 40. [Google Scholar] [CrossRef]
- Graves, J.A.; Metukuri, M.; Scott, D.; Rothermund, K.; Prochownik, E.V. Regulation of reactive oxygen species homeostasis by peroxiredoxins and c-myc. J. Biol. Chem. 2009, 284, 6520–6529. [Google Scholar] [CrossRef]
- Ferro, E.; Goitre, L.; Retta, S.F.; Trabalzini, L. The interplay between ROS and Ras GTPases: Physiological and pathological implications. J. Signal. Transduct. 2012, 2012, 365769. [Google Scholar] [CrossRef]
- Jackstadt, R.; Hermeking, H. MicroRNAs as regulators and mediators of c-Myc function. Biochim. Biophys. Acta 2015, 1849, 544–553. [Google Scholar] [CrossRef]
- Xue, G.; Yan, H.L.; Zhang, Y.; Hao, L.Q.; Zhu, X.T.; Mei, Q.; Sun, S.H. c-MYC-mediated repression of miR-15-16 in hypoxia is induced by increased HIF-2alpha and promotes tumor angiogenesis and metastasis by upregulating FGF2. Oncogene 2015, 34, 1393–1406. [Google Scholar] [CrossRef]
- Li, S.G.; Shi, Q.W.; Yuan, L.Y.; Qin, L.P.; Wang, Y.; Miao, Y.Q.; Chen, Z.; Ling, C.Q.; Qin, W.X. c-Myc-dependent repression of two oncogenic miRNA clusters contributes to triptolide-induced cell death in hepatocellular carcinoma cells. J. Exp. Clin. Cancer Res. 2018, 37, 51. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, A.; Falcone, E.; Garibaldi, F.; Piaggio, G. Dysregulation of microRNA biogenesis in cancer: The impact of mutant p53 on drosha complex activity. J. Exp. Clin. Cancer Res. 2016, 35, 45. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.R.; Vuppusetty, C.; Colley, T.; Papaioannou, A.I.; Fenwick, P.; Donnelly, L.; Ito, K.; Barnes, P.J. Oxidative stress dependent microRNA-34a activation via pi3kalpha reduces the expression of sirtuin-1 and sirtuin-6 in epithelial cells. Sci. Rep. 2016, 6, 35871. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Yan, W.; Lu, L.; Wang, Y.; Lu, W.; Cao, Y.; Cai, W. P38/p53/miR-200a-3p feedback loop promotes oxidative stress-mediated liver cell death. Cell Cycle 2015, 14, 1548–1558. [Google Scholar] [CrossRef] [PubMed]
- Magenta, A.; Cencioni, C.; Fasanaro, P.; Zaccagnini, G.; Greco, S.; Sarra-Ferraris, G.; Antonini, A.; Martelli, F.; Capogrossi, M.C. MiR-200c is upregulated by oxidative stress and induces endothelial cell apoptosis and senescence via zeb1 inhibition. Cell Death Differ. 2011, 18, 1628–1639. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ding, J.; Yang, J.; Guo, X.; Zheng, Y. MicroRNA roles in the nuclear factor kappa b signaling pathway in cancer. Front. Immunol. 2018, 9, 546. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Becker Buscaglia, L.E.; Barker, J.R.; Li, Y. MicroRNAs in NF-kappaB signaling. J. Mol. Cell Biol. 2011, 3, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Bazzoni, F.; Rossato, M.; Fabbri, M.; Gaudiosi, D.; Mirolo, M.; Mori, L.; Tamassia, N.; Mantovani, A.; Cassatella, M.A.; Locati, M. Induction and regulatory function of miR-9 in human monocytes and neutrophils exposed to proinflammatory signals. Proc. Natl. Acad. Sci. USA 2009, 106, 5282–5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, J.; Shi, Y.; Tan, G.; Yang, C.H.; Fan, M.; Pfeffer, L.M.; Wu, Z.H. DNA damage induces NF-kappaB-dependent microRNA-21 up-regulation and promotes breast cancer cell invasion. J. Biol. Chem. 2012, 287, 21783–21795. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Guo, Y.; Li, P.; Chao, L. Role of kallistatin treatment in aging and cancer by modulating miR-34a and miR-21 expression. Oxid. Med. Cell. Longev. 2017, 2017, 5025610. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Son, Y.O.; Divya, S.P.; Wang, L.; Zhang, Z.; Shi, X. Oncogenic transformation of human lung bronchial epithelial cells induced by arsenic involves ROS-dependent activation of STAT3-miR-21-PDCD4 mechanism. Sci. Rep. 2016, 6, 37227. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Hu, G.; Gong, A.Y.; Chen, X.M. Binding of NF-kappaB p65 subunit to the promoter elements is involved in LPS-induced transactivation of miRNA genes in human biliary epithelial cells. Nucleic Acids Res. 2010, 38, 3222–3232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Tan, W.; Cheng, S.; Wang, H.; Ye, S.; Yu, C.; He, Y.; Zeng, J.; Cen, J.; Hu, J.; et al. Upregulation of microRNA-126 in hepatic stellate cells may affect pathogenesis of liver fibrosis through the NF-kappaB pathway. DNA Cell Biol. 2015, 34, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Ren, X.; Zhang, X.; Luo, Y.; Wang, G.; Huang, K.; Feng, S.; Bao, X.; Huang, K.; He, X.; et al. Selective killing of lung cancer cells by miRNA-506 molecule through inhibiting NF-kappaB p65 to evoke reactive oxygen species generation and p53 activation. Oncogene 2015, 34, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Sykiotis, G.P.; Bohmann, D. Stress-activated cap’n’collar transcription factors in aging and human disease. Sci. Signal. 2010, 3, re3. [Google Scholar] [CrossRef] [PubMed]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Zhang, C.; Shu, L.; Kong, A.N. MicroRNAs: New players in cancer prevention targeting Nrf2, oxidative stress and inflammatory pathways. Curr. Pharmacol. Rep. 2015, 1, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.H.; Ku, C.H.; Siow, R.C.M. Regulation of the Nrf2 antioxidant pathway by microRNAs: New players in micromanaging redox homeostasis. Free Radic. Biol. Med. 2013, 64, 4–11. [Google Scholar] [CrossRef]
- Singh, A.; Happel, C.; Manna, S.K.; Acquaah-Mensah, G.; Carrerero, J.; Kumar, S.; Nasipuri, P.; Krausz, K.W.; Wakabayashi, N.; Dewi, R.; et al. Transcription factor Nrf2 regulates miR-1 and miR-206 to drive tumorigenesis. J. Clin. Investig. 2013, 123, 2921–2934. [Google Scholar] [CrossRef]
- Kurinna, S.; Schafer, M.; Ostano, P.; Karouzakis, E.; Chiorino, G.; Bloch, W.; Bachmann, A.; Gay, S.; Garrod, D.; Lefort, K.; et al. A novel Nrf2-miR-29-desmocollin-2 axis regulates desmosome function in keratinocytes. Nat. Commun. 2014, 5, 5099. [Google Scholar] [CrossRef] [Green Version]
- Joo, M.S.; Lee, C.G.; Koo, J.H.; Kim, S.G. Mir-125b transcriptionally increased by nrf2 inhibits AHR repressor, which protects kidney from cisplatin-induced injury. Cell Death Dis. 2013, 4, e899. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yao, Y.; Eades, G.; Zhang, Y.; Zhou, Q. Mir-28 regulates Nrf2 expression through a Keap1-independent mechanism. Breast Cancer Res. Treat. 2011, 129, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, Y.; Qin, J.; Lu, S. The role of mir-34a in the hepatoprotective effect of hydrogen sulfide on ischemia/reperfusion injury in young and old rats. PLoS ONE 2014, 9, e113305. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Ronghe, A.M.; Chatterjee, A.; Bhat, N.K.; Bhat, H.K. MicroRNA-93 regulates Nrf2 expression and is associated with breast carcinogenesis. Carcinogenesis 2013, 34, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Eades, G.; Yang, M.; Yao, Y.; Zhang, Y.; Zhou, Q. Mir-200a regulates Nrf2 activation by targeting Keap1 mRNA in breast cancer cells. J. Biol. Chem. 2011, 286, 40725–40733. [Google Scholar] [CrossRef] [PubMed]
- Nebbioso, A.; Tambaro, F.P.; Dell’Aversana, C.; Altucci, L. Cancer epigenetics: Moving forward. PLoS Genet. 2018, 14, e1007362. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Xu, Q.; Jing, Y.; Agani, F.; Qian, X.; Carpenter, R.; Li, Q.; Wang, X.R.; Peiper, S.S.; Lu, Z.; et al. Reactive oxygen species regulate ERBB2 and ERBB3 expression via miR-199a/125b and DNA methylation. EMBO Rep. 2012, 13, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Druz, A.; Betenbaugh, M.; Shiloach, J. Glucose depletion activates mmu-miR-466h-5p expression through oxidative stress and inhibition of histone deacetylation. Nucleic Acids Res. 2012, 40, 7291–7302. [Google Scholar] [CrossRef] [Green Version]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The role of hypoxia in cancer progression, angiogenesis, metastasis, and resistance to therapy. Hypoxia 2015, 3, 83–92. [Google Scholar] [CrossRef]
- Azimi, I.; Petersen, R.M.; Thompson, E.W.; Roberts-Thomson, S.J.; Monteith, G.R. Hypoxia-induced reactive oxygen species mediate n-cadherin and serpine1 expression, EGFR signalling and motility in mda-mb-468 breast cancer cells. Sci. Rep. 2017, 7, 15140. [Google Scholar] [CrossRef]
- Cardin, R.; Piciocchi, M.; Sinigaglia, A.; Lavezzo, E.; Bortolami, M.; Kotsafti, A.; Cillo, U.; Zanus, G.; Mescoli, C.; Rugge, M.; et al. Oxidative DNA damage correlates with cell immortalization and miR-92 expression in hepatocellular carcinoma. BMC Cancer 2012, 12, 177. [Google Scholar] [CrossRef]
- Yatabe, N.; Kyo, S.; Maida, Y.; Nishi, H.; Nakamura, M.; Kanaya, T.; Tanaka, M.; Isaka, K.; Ogawa, S.; Inoue, M. HIF-1-mediated activation of telomerase in cervical cancer cells. Oncogene 2004, 23, 3708–3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, N.; Kwon, H.J.; Jung, H.J. Downregulation of mitochondrial UQCRB inhibits cancer stem cell-like properties in glioblastoma. Int. J. Oncol. 2018, 52, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, S.E.; O’Neill, L.A. HIF1alpha and metabolic reprogramming in inflammation. J. Clin. Investig. 2016, 126, 3699–3707. [Google Scholar] [CrossRef] [PubMed]
- Bavelloni, A.; Ramazzotti, G.; Poli, A.; Piazzi, M.; Focaccia, E.; Blalock, W.; Faenza, I. Mirna-210: A current overview. Anticancer Res. 2017, 37, 6511–6521. [Google Scholar]
- Fallah, A.; Sadeghinia, A.; Kahroba, H.; Samadi, A.; Heidari, H.R.; Bradaran, B.; Zeinali, S.; Molavi, O. Therapeutic targeting of angiogenesis molecular pathways in angiogenesis-dependent diseases. Biomed. Pharmacother. 2019, 110, 775–785. [Google Scholar] [CrossRef]
- Hoeben, A.; Landuyt, B.; Highley, M.S.; Wildiers, H.; Van Oosterom, A.T.; De Bruijn, E.A. Vascular endothelial growth factor and angiogenesis. Pharmacol. Rev. 2004, 56, 549–580. [Google Scholar] [CrossRef]
- Ferrari, G.; Cook, B.D.; Terushkin, V.; Pintucci, G.; Mignatti, P. Transforming growth factor-beta 1 (TGF-beta1) induces angiogenesis through vascular endothelial growth factor (VEGF)-mediated apoptosis. J. Cell. Physiol. 2009, 219, 449–458. [Google Scholar] [CrossRef]
- Fagiani, E.; Christofori, G. Angiopoietins in angiogenesis. Cancer Lett. 2013, 328, 18–26. [Google Scholar] [CrossRef]
- Raica, M.; Cimpean, A.M. Platelet-derived growth factor (PDGF)/PDGFreceptors (PDGFR) axis as target for antitumor and antiangiogenic therapy. Pharmaceuticals 2010, 3, 572–599. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Wilson, R.; Chatterjee, N.; Hudson, C.; Ruhl, R.; Hipfinger, C.; Helms, E.; Khan, O.F.; Anderson, D.G.; Anand, S. MicroRNA regulation of the MRN complex impacts DNA damage, cellular senescence, and angiogenic signaling. Cell Death Dis. 2018, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.Y.; Ramakrishnan, D.P.; Silverstein, R.L. Thrombospondin-1 modulates VEGF signaling via CD36 by recruiting SHP-1 to VEGFR2 complex in microvascular endothelial cells. Blood 2013, 122, 1822–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Kozlov, S.; Lavin, M.F.; Person, M.D.; Paull, T.T. Atm activation by oxidative stress. Science 2010, 330, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Okuno, Y.; Nakamura-Ishizu, A.; Otsu, K.; Suda, T.; Kubota, Y. Pathological neoangiogenesis depends on oxidative stress regulation by atm. Nat. Med. 2012, 18, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Economopoulou, M.; Langer, H.F.; Celeste, A.; Orlova, V.V.; Choi, E.Y.; Ma, M.; Vassilopoulos, A.; Callen, E.; Deng, C.; Bassing, C.H.; et al. Histone h2ax is integral to hypoxia-driven neovascularization. Nat. Med. 2009, 15, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Espinosa-Diez, C.; Kanner, N.; Chatterjee, N.; Ruhl, R.; Hipfinger, C.; Advani, S.J.; Li, J.; Khan, O.F.; Franovic, A.; et al. MicroRNA regulation of endothelial TREX1 reprograms the tumour microenvironment. Nat. Commun. 2016, 7, 13597. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wa, Q.D.; Lu, C.; Pan, W.; Lu, Z.; Ao, J. Mir3283p enhances the radiosensitivity of osteosarcoma and regulates apoptosis and cell viability via H2AX. Oncol. Rep. 2018, 39, 545–553. [Google Scholar] [PubMed]
- Marampon, F.; Codenotti, S.; Megiorni, F.; Del Fattore, A.; Camero, S.; Gravina, G.L.; Festuccia, C.; Musio, D.; De Felice, F.; Nardone, V.; et al. NRF2 orchestrates the redox regulation induced by radiation therapy, sustaining embryonal and alveolar rhabdomyosarcoma cells radioresistance. J. Cancer Res. Clin. Oncol. 2019, 145, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Crosstalk between oxidative stress and SIRT1: Impact on the aging process. Int. J. Mol. Sci. 2013, 14, 3834–3859. [Google Scholar] [CrossRef]
- Luo, J.; Chen, P.; Xie, W.; Wu, F. MicroRNA-138 inhibits cell proliferation in hepatocellular carcinoma by targeting SIRT1. Oncol. Rep. 2017, 38, 1067–1074. [Google Scholar] [CrossRef]
- Zhou, B.; Li, C.; Qi, W.; Zhang, Y.; Zhang, F.; Wu, J.X.; Hu, Y.N.; Wu, D.M.; Liu, Y.; Yan, T.T.; et al. Downregulation of miR-181a upregulates sirtuin-1 (SIRT1) and improves hepatic insulin sensitivity. Diabetologia 2012, 55, 2032–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamakuchi, M. MicroRNA regulation of SIRT1. Front. Physiol. 2012, 3, 68. [Google Scholar] [CrossRef] [PubMed]
- Rane, S.; He, M.; Sayed, D.; Vashistha, H.; Malhotra, A.; Sadoshima, J.; Vatner, D.E.; Vatner, S.F.; Abdellatif, M. Downregulation of miR-199a derepresses hypoxia-inducible factor-1alpha and Sirtuin 1 and recapitulates hypoxia preconditioning in cardiac myocytes. Circ. Res. 2009, 104, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging biological principles of metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed]
- Peiris-Pages, M.; Martinez-Outschoorn, U.E.; Sotgia, F.; Lisanti, M.P. Metastasis and oxidative stress: Are antioxidants a metabolic driver of progression? Cell Metab. 2015, 22, 956–958. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.G.; Piskounova, E.; Morrison, S.J. Cancer, oxidative stress, and metastasis. Cold Spring Harb. Symp. Quant. Biol. 2016, 81, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.L.; Liu, C.M.; Chen, H.A.; Yang, S.T.; Shigemura, K.; Kitagawa, K.; Yamamichi, F.; Fujisawa, M.; Liu, Y.R.; Lee, W.H.; et al. Reactive oxygen species-mediated switching expression of MMP-3 in stromal fibroblasts and cancer cells during prostate cancer progression. Sci. Rep. 2017, 7, 9065. [Google Scholar] [CrossRef] [PubMed]
- Cichon, M.A.; Radisky, D.C. ROS-induced epithelial-mesenchymal transition in mammary epithelial cells is mediated by NF-kB-dependent activation of snail. Oncotarget 2014, 5, 2827–2838. [Google Scholar] [CrossRef]
- Zeller, K.S.; Riaz, A.; Sarve, H.; Li, J.; Tengholm, A.; Johansson, S. The role of mechanical force and ROS in integrin-dependent signals. PLoS ONE 2013, 8, e64897. [Google Scholar] [CrossRef]
- Mori, K.; Shibanuma, M.; Nose, K. Invasive potential induced under long-term oxidative stress in mammary epithelial cells. Cancer Res. 2004, 64, 7464–7472. [Google Scholar] [CrossRef]
- Sawicki, G. Intracellular regulation of matrix metalloproteinase-2 activity: New strategies in treatment and protection of heart subjected to oxidative stress. Scientifica 2013, 2013, 130451. [Google Scholar] [CrossRef] [PubMed]
- Dezerega, A.; Madrid, S.; Mundi, V.; Valenzuela, M.A.; Garrido, M.; Paredes, R.; Garcia-Sesnich, J.; Ortega, A.V.; Gamonal, J.; Hernandez, M. Pro-oxidant status and matrix metalloproteinases in apical lesions and gingival crevicular fluid as potential biomarkers for asymptomatic apical periodontitis and endodontic treatment response. J. Inflamm. 2012, 9, 8. [Google Scholar] [CrossRef]
- Yoon, S.O.; Park, S.J.; Yoon, S.Y.; Yun, C.H.; Chung, A.S. Sustained production of H(2)O(2) activates pro-matrix metalloproteinase-2 through receptor tyrosine kinases/phosphatidylinositol 3-kinase/NF-kappa B pathway. J. Biol. Chem. 2002, 277, 30271–30282. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Wang, J.; Nie, G.; Chen, Y.J.; Li, X.; Jiang, X.; Cao, W.H. MicroRNA-509-5p functions as an anti-oncogene in breast cancer via targeting sod2. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 3617–3625. [Google Scholar] [PubMed]
- Yu, L.; Chen, X.; Sun, X.; Wang, L.; Chen, S. The glycolytic switch in tumors: How many players are involved? J. Cancer 2017, 8, 3430–3440. [Google Scholar] [CrossRef] [PubMed]
- Pacella, I.; Procaccini, C.; Focaccetti, C.; Miacci, S.; Timperi, E.; Faicchia, D.; Severa, M.; Rizzo, F.; Coccia, E.M.; Bonacina, F.; et al. Fatty acid metabolism complements glycolysis in the selective regulatory t cell expansion during tumor growth. Proc. Natl. Acad. Sci. USA 2018, 115, E6546–E6555. [Google Scholar] [CrossRef]
- Schwartz, L.; Supuran, C.T.; Alfarouk, K.O. The warburg effect and the hallmarks of cancer. Anticancer Agents Med. Chem. 2017, 17, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, J.; Bae, J.S. ROS homeostasis and metabolism: A critical liaison for cancer therapy. Exp. Mol. Med. 2016, 48, e269. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, C.; Wu, R.; Sun, Y.; Levine, A.; Feng, Z. Glutaminase 2, a novel p53 target gene regulating energy metabolism and antioxidant function. Proc. Natl. Acad. Sci. USA 2010, 107, 7455–7460. [Google Scholar] [CrossRef] [Green Version]
- Budanov, A.V. The role of tumor suppressor p53 in the antioxidant defense and metabolism. Subcell. Biochem. 2014, 85, 337–358. [Google Scholar]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mello, T.; Simeone, I.; Galli, A. Mito-nuclear communication in hepatocellular carcinoma metabolic rewiring. Cells 2019, 8, 417. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Nishino, T.; Baba, O.; Kuwabara, Y.; Nakao, T.; Nishiga, M.; Usami, S.; Izuhara, M.; Sowa, N.; Yahagi, N.; et al. MicroRNA-33 regulates sterol regulatory element-binding protein 1 expression in mice. Nat. Commun. 2013, 4, 2883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayob, A.Z.; Ramasamy, T.S. Cancer stem cells as key drivers of tumour progression. J. Biomed. Sci. 2018, 25, 20. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.D.; Savage, J.E.; Cao, L.; Soule, B.P.; Ly, D.; DeGraff, W.; Harris, C.C.; Mitchell, J.B.; Simone, N.L. Cellular stress induced alterations in microRNA let-7a and let-7b expression are dependent on p53. PLoS ONE 2011, 6, e24429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ng, W.L.; Wang, P.; Tian, L.; Werner, E.; Wang, H.; Doetsch, P.; Wang, Y. MicroRNA-21 modulates the levels of reactive oxygen species by targeting sod3 and tnfalpha. Cancer Res. 2012, 72, 4707–4713. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Ren, Z.J.; Tang, J.H. MicroRNA-34a: A potential therapeutic target in human cancer. Cell Death Dis. 2014, 5, e1327. [Google Scholar] [CrossRef]
- Balzano, F.; Cruciani, S.; Basoli, V.; Santaniello, S.; Facchin, F.; Ventura, C.; Maioli, M. Mir200 and miR302: Two big families influencing stem cell behavior. Molecules 2018, 23, 282. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, S.G.; Song, S.Y.; Kim, J.K.; Sung, J.H. Reactive oxygen species-responsive miR-210 regulates proliferation and migration of adipose-derived stem cells via ptpn2. Cell Death Dis. 2013, 4, e588. [Google Scholar] [CrossRef]
- Mei, Y.; Bian, C.; Li, J.; Du, Z.; Zhou, H.; Yang, Z.; Zhao, R.C. Mir-21 modulates the erk-mapk signaling pathway by regulating spry2 expression during human mesenchymal stem cell differentiation. J. Cell. Biochem. 2013, 114, 1374–1384. [Google Scholar] [CrossRef]
- Yu, G.; Yao, W.; Xiao, W.; Li, H.; Xu, H.; Lang, B. MicroRNA-34a functions as an anti-metastatic microRNA and suppresses angiogenesis in bladder cancer by directly targeting cd44. J. Exp. Clin. Cancer Res. 2014, 33, 779. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E. Let-7 and miR-200 microRNAs: Guardians against pluripotency and cancer progression. Cell Cycle 2009, 8, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Yang, Z.; Zhu, Q.; Wu, Y.; Sun, K.; Alahdal, M.; Zhang, Y.; Xing, Y.; Shen, Y.; Xia, T.; et al. Up-regulation of miR-210 induced by a hypoxic microenvironment promotes breast cancer stem cells metastasis, proliferation, and self-renewal by targeting e-cadherin. FASEB J. 2018, 32, fj201801013R. [Google Scholar] [CrossRef] [PubMed]
- Maciel-Baron, L.A.; Moreno-Blas, D.; Morales-Rosales, S.L.; Gonzalez-Puertos, V.Y.; Lopez-Diazguerrero, N.E.; Torres, C.; Castro-Obregon, S.; Konigsberg, M. Cellular senescence, neurological function, and redox state. Antioxid. Redox Signal. 2018, 28, 1704–1723. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Xu, Y. P53, oxidative stress, and aging. Antioxid. Redox Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Dellago, H.; Preschitz-Kammerhofer, B.; Terlecki-Zaniewicz, L.; Schreiner, C.; Fortschegger, K.; Chang, M.W.; Hackl, M.; Monteforte, R.; Kuhnel, H.; Schosserer, M.; et al. High levels of oncomiR-21 contribute to the senescence-induced growth arrest in normal human cells and its knock-down increases the replicative lifespan. Aging Cell 2013, 12, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Takeshita, F.; Hino, Y.; Fukunaga, S.; Kudo, Y.; Tamaki, A.; Matsunaga, J.; Takahashi, R.U.; Takata, T.; Shimamoto, A.; et al. Mir-22 represses cancer progression by inducing cellular senescence. J. Cell Biol. 2011, 193, 409–424. [Google Scholar] [CrossRef]
- Hu, Z.; Klein, J.D.; Mitch, W.E.; Zhang, L.; Martinez, I.; Wang, X.H. MicroRNA-29 induces cellular senescence in aging muscle through multiple signaling pathways. Aging 2014, 6, 160–175. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Yang, A.; McDonald, D.G.; Riemer, E.C.; Vanek, K.N.; Schulte, B.A.; Wang, G.Y. Mir-34a modulates ionizing radiation-induced senescence in lung cancer cells. Oncotarget 2017, 8, 69797–69807. [Google Scholar] [CrossRef]
- Hong, L.; Lai, M.; Chen, M.; Xie, C.; Liao, R.; Kang, Y.J.; Xiao, C.; Hu, W.Y.; Han, J.; Sun, P. The miR-17-92 cluster of microRNAs confers tumorigenicity by inhibiting oncogene-induced senescence. Cancer Res. 2010, 70, 8547–8557. [Google Scholar] [CrossRef]
- Nyholm, A.M.; Lerche, C.M.; Manfe, V.; Biskup, E.; Johansen, P.; Morling, N.; Thomsen, B.M.; Glud, M.; Gniadecki, R. Mir-125b induces cellular senescence in malignant melanoma. BMC Dermatol. 2014, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Lazzarini, R.; Recchioni, R.; Marcheselli, F.; Rippo, M.R.; Di Nuzzo, S.; Albertini, M.C.; Graciotti, L.; Babini, L.; Mariotti, S.; et al. Mir-146a as marker of senescence-associated pro-inflammatory status in cells involved in vascular remodelling. Age 2013, 35, 1157–1172. [Google Scholar] [CrossRef] [PubMed]
- Menghini, R.; Casagrande, V.; Cardellini, M.; Martelli, E.; Terrinoni, A.; Amati, F.; Vasa-Nicotera, M.; Ippoliti, A.; Novelli, G.; Melino, G.; et al. MicroRNA 217 modulates endothelial cell senescence via silent information regulator 1. Circulation 2009, 120, 1524–1532. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Wang, S.C.; Hsu, C.Y.; Miao, Y.; Martin, M.; Yin, Y.; Wu, C.C.; Wang, Y.T.; Wu, G.; Chien, S.; et al. MicroRNA-92a mediates endothelial dysfunction in ckd. J. Am. Soc. Nephrol. 2017, 28, 3251–3261. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, H.Y.; Wang, W.Y.; Zhao, Z.L.; Liu, X.Y.; Wang, L.Y. Regulation of miR-92a on vascular endothelial aging via mediating nrf2-keap1-are signal pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2734–2742. [Google Scholar] [PubMed]
- Sanchez-Sanchez, B.; Gutierrez-Herrero, S.; Lopez-Ruano, G.; Prieto-Bermejo, R.; Romo-Gonzalez, M.; Llanillo, M.; Pandiella, A.; Guerrero, C.; Miguel, J.F.; Sanchez-Guijo, F.; et al. Nadph oxidases as therapeutic targets in chronic myelogenous leukemia. Clin. Cancer Res. 2014, 20, 4014–4025. [Google Scholar] [CrossRef]
- Altenhofer, S.; Radermacher, K.A.; Kleikers, P.W.; Wingler, K.; Schmidt, H.H. Evolution of nadph oxidase inhibitors: Selectivity and mechanisms for target engagement. Antioxid. Redox Signal. 2015, 23, 406–427. [Google Scholar] [CrossRef]
- Suzuki, S.; Pitchakarn, P.; Sato, S.; Shirai, T.; Takahashi, S. Apocynin, an nadph oxidase inhibitor, suppresses progression of prostate cancer via rac1 dephosphorylation. Exp. Toxicol. Pathol. 2013, 65, 1035–1041. [Google Scholar] [CrossRef]
- Fuji, S.; Suzuki, S.; Naiki-Ito, A.; Kato, H.; Hayakawa, M.; Yamashita, Y.; Kuno, T.; Takahashi, S. The nadph oxidase inhibitor apocynin suppresses preneoplastic liver foci of rats. Toxicol. Pathol. 2017, 45, 544–550. [Google Scholar] [CrossRef]
- Kumari, S.; Badana, A.K.; G, M.M.; G, S.; Malla, R. Reactive oxygen species: A key constituent in cancer survival. Biomark. Insights 2018, 13, 1177271918755391. [Google Scholar] [CrossRef]
- Traverso, N.; Ricciarelli, R.; Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Role of glutathione in cancer progression and chemoresistance. Oxid. Med. Cell. Longev. 2013, 2013, 972913. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, T.W.; Zhou, Y.; Peng, H.; Liu, T.; Zandi, E.; Martinez-Chantar, M.L.; Mato, J.M.; Lu, S.C. Activation of a novel c-Myc-miR27-prohibitin 1 circuitry in cholestatic liver injury inhibits glutathione synthesis in mice. Antioxid. Redox Signal. 2015, 22, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Diez, C.; Fierro-Fernandez, M.; Sanchez-Gomez, F.; Rodriguez-Pascual, F.; Alique, M.; Ruiz-Ortega, M.; Beraza, N.; Martinez-Chantar, M.L.; Fernandez-Hernando, C.; Lamas, S. Targeting of gamma-glutamyl-cysteine ligase by miR-433 reduces glutathione biosynthesis and promotes tgf-beta-dependent fibrogenesis. Antioxid. Redox Signal. 2015, 23, 1092–1105. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhou, M.; Zhou, W. MicroRNA-30e regulates tgf-beta-mediated nadph oxidase 4-dependent oxidative stress by snai1 in atherosclerosis. Int. J. Mol. Med. 2019, 43, 1806–1816. [Google Scholar] [PubMed]
- Cho, K.J.; Song, J.; Oh, Y.; Lee, J.E. MicroRNA-let-7a regulates the function of microglia in inflammation. Mol. Cell. Neurosci. 2015, 68, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Fulciniti, M.; Amodio, N.; Bandi, R.L.; Cagnetta, A.; Samur, M.K.; Acharya, C.; Prabhala, R.; D’Aquila, P.; Bellizzi, D.; Passarino, G.; et al. Mir-23b/sp1/c-myc forms a feed-forward loop supporting multiple myeloma cell growth. Blood Cancer J. 2016, 6, e380. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, F.; Wang, Y.; Liu, Z.; Sun, A.; Zen, K.; Zhang, C.Y.; Zhang, Q. The transcription factor c-myc suppresses miR-23b and miR-27b transcription during fetal distress and increases the sensitivity of neurons to hypoxia-induced apoptosis. PLoS ONE 2015, 10, e0120217. [Google Scholar] [CrossRef]
- Gao, P.; Tchernyshyov, I.; Chang, T.C.; Lee, Y.S.; Kita, K.; Ochi, T.; Zeller, K.I.; De Marzo, A.M.; Van Eyk, J.E.; Mendell, J.T.; et al. C-myc suppression of miR-23a/b enhances mitochondrial glutaminase expression and glutamine metabolism. Nature 2009, 458, 762–765. [Google Scholar] [CrossRef]
- Mott, J.L.; Kurita, S.; Cazanave, S.C.; Bronk, S.F.; Werneburg, N.W.; Fernandez-Zapico, M.E. Transcriptional suppression of miR-29b-1/miR-29a promoter by c-myc, hedgehog, and nf-kappab. J. Cell. Biochem. 2010, 110, 1155–1164. [Google Scholar] [CrossRef]
- Hou, M.; Zuo, X.; Li, C.; Zhang, Y.; Teng, Y. Mir-29b regulates oxidative stress by targeting sirt1 in ovarian cancer cells. Cell. Physiol. Biochem. 2017, 43, 1767–1776. [Google Scholar] [CrossRef]
- Mazzoccoli, L.; Robaina, M.C.; Apa, A.G.; Bonamino, M.; Pinto, L.W.; Queiroga, E.; Bacchi, C.E.; Klumb, C.E. Mir-29 silencing modulates the expression of target genes related to proliferation, apoptosis and methylation in burkitt lymphoma cells. J. Cancer Res. Clin. Oncol. 2018, 144, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.W.; Hsu, N.Y.; Wang, Y.C.; Lee, M.C.; Cheng, Y.W.; Chen, C.Y.; Lee, H. C-myc suppresses microRNA-29b to promote tumor aggressiveness and poor outcomes in non-small cell lung cancer by targeting fhit. Oncogene 2015, 34, 2072–2082. [Google Scholar] [CrossRef] [PubMed]
- Okada, N.; Lin, C.P.; Ribeiro, M.C.; Biton, A.; Lai, G.; He, X.; Bu, P.; Vogel, H.; Jablons, D.M.; Keller, A.C.; et al. A positive feedback between p53 and miR-34 miRNAs mediates tumor suppression. Genes Dev. 2014, 28, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Mihailovich, M.; Bremang, M.; Spadotto, V.; Musiani, D.; Vitale, E.; Varano, G.; Zambelli, F.; Mancuso, F.M.; Cairns, D.A.; Pavesi, G.; et al. Mir-17-92 fine-tunes myc expression and function to ensure optimal b cell lymphoma growth. Nat. Commun. 2015, 6, 8725. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.L.; Xue, G.; Mei, Q.; Wang, Y.Z.; Ding, F.X.; Liu, M.F.; Lu, M.H.; Tang, Y.; Yu, H.Y.; Sun, S.H. Repression of the miR-17-92 cluster by p53 has an important function in hypoxia-induced apoptosis. EMBO J. 2009, 28, 2719–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borkowski, R.; Du, L.; Zhao, Z.; McMillan, E.; Kosti, A.; Yang, C.R.; Suraokar, M.; Wistuba, I.I.; Gazdar, A.F.; Minna, J.D.; et al. Genetic mutation of p53 and suppression of the miR-17 approximately 92 cluster are synthetic lethal in non-small cell lung cancer due to upregulation of vitamin d signaling. Cancer Res. 2015, 75, 666–675. [Google Scholar] [CrossRef] [PubMed]
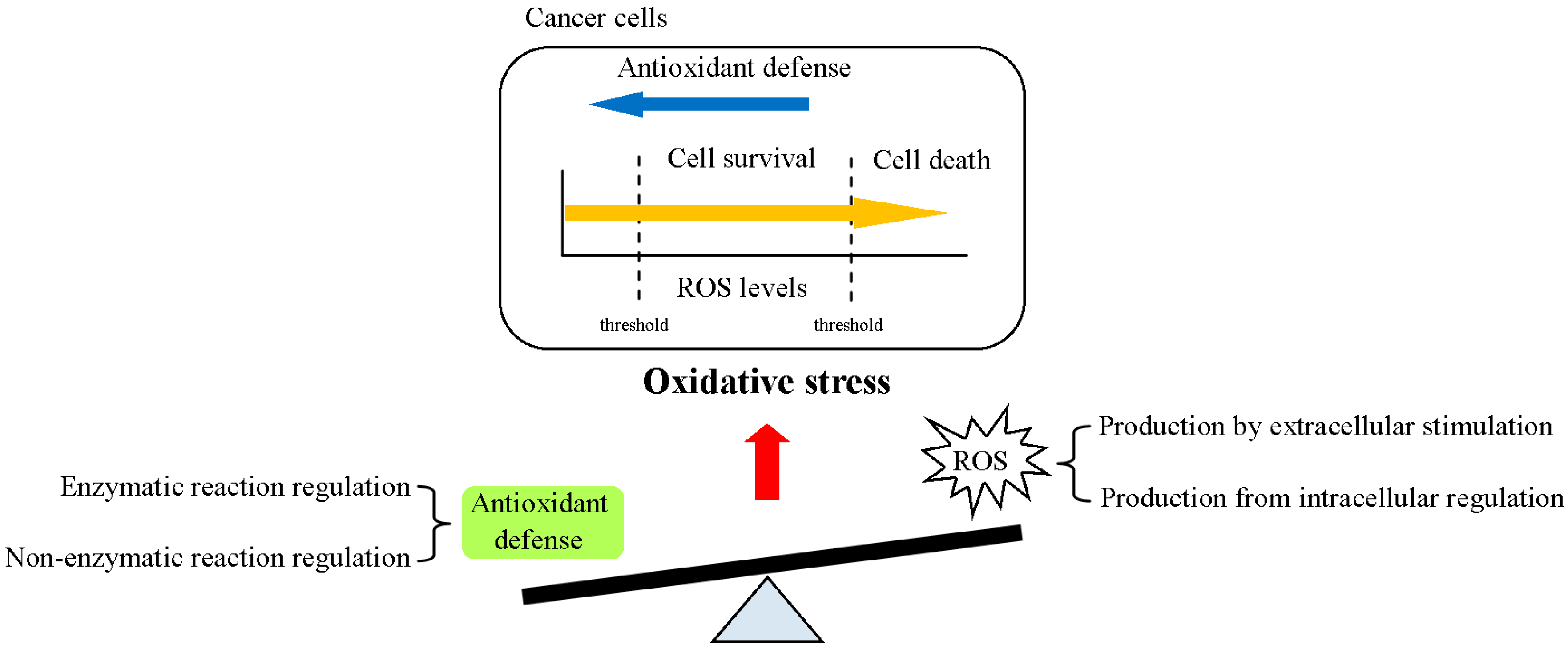
Figure 1.
Reactive oxygen species (ROS) production and antioxidant defense in the control of redox homeostasis in cancer cells. Disruption of redox homeostasis by ROS (intra- or extracellular signals) and antioxidant defense (enzymatic or non-enzymatic reactions) induces oxidative stress (OS) and results in various cell functions. The physiological function of ROS is dependent on its concentration. Elevated ROS production and accumulation lead to cell apoptosis. On the other hand, medium levels of ROS promote cell survival and progression.
Figure 1.
Reactive oxygen species (ROS) production and antioxidant defense in the control of redox homeostasis in cancer cells. Disruption of redox homeostasis by ROS (intra- or extracellular signals) and antioxidant defense (enzymatic or non-enzymatic reactions) induces oxidative stress (OS) and results in various cell functions. The physiological function of ROS is dependent on its concentration. Elevated ROS production and accumulation lead to cell apoptosis. On the other hand, medium levels of ROS promote cell survival and progression.

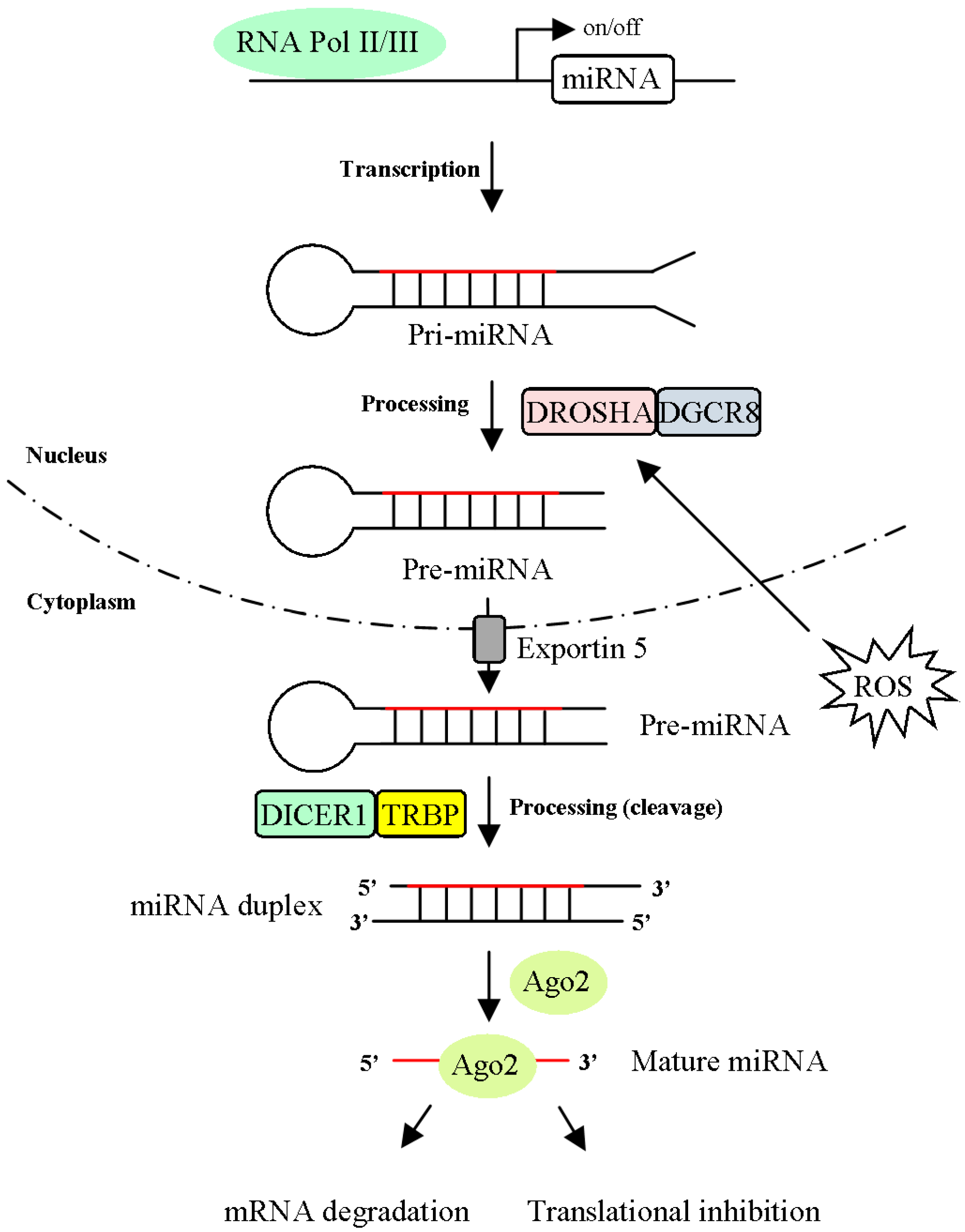
Figure 2.
The biogenesis and regulation mechanisms of microRNAs (miRNAs). MiRNAs are transcribed by RNA polymerase II/III and generated the primary miRNA transcript (pri-miRNA). The pri-miRNAs are cleaved into precursor miRNA transcript (pre-miRNA) by the microprocessor complex, a combination of DROSHA and DGCR8. Pre-miRNA is exported to cytoplasm via exportin 5 and further processed by the RNase III enzyme Dicer with the cofactor protein TRBP to generate an approximately 18-25-nt duplex. Either 5p or 3p strand of the mature miRNA (red line) interacts with Argonaute (Ago) protein and forms a miRNA-induced silencing complex (miRISC). There are two models (mRNA degradation and translational repression) of miRNA-mediated gene silencing.
Figure 2.
The biogenesis and regulation mechanisms of microRNAs (miRNAs). MiRNAs are transcribed by RNA polymerase II/III and generated the primary miRNA transcript (pri-miRNA). The pri-miRNAs are cleaved into precursor miRNA transcript (pre-miRNA) by the microprocessor complex, a combination of DROSHA and DGCR8. Pre-miRNA is exported to cytoplasm via exportin 5 and further processed by the RNase III enzyme Dicer with the cofactor protein TRBP to generate an approximately 18-25-nt duplex. Either 5p or 3p strand of the mature miRNA (red line) interacts with Argonaute (Ago) protein and forms a miRNA-induced silencing complex (miRISC). There are two models (mRNA degradation and translational repression) of miRNA-mediated gene silencing.

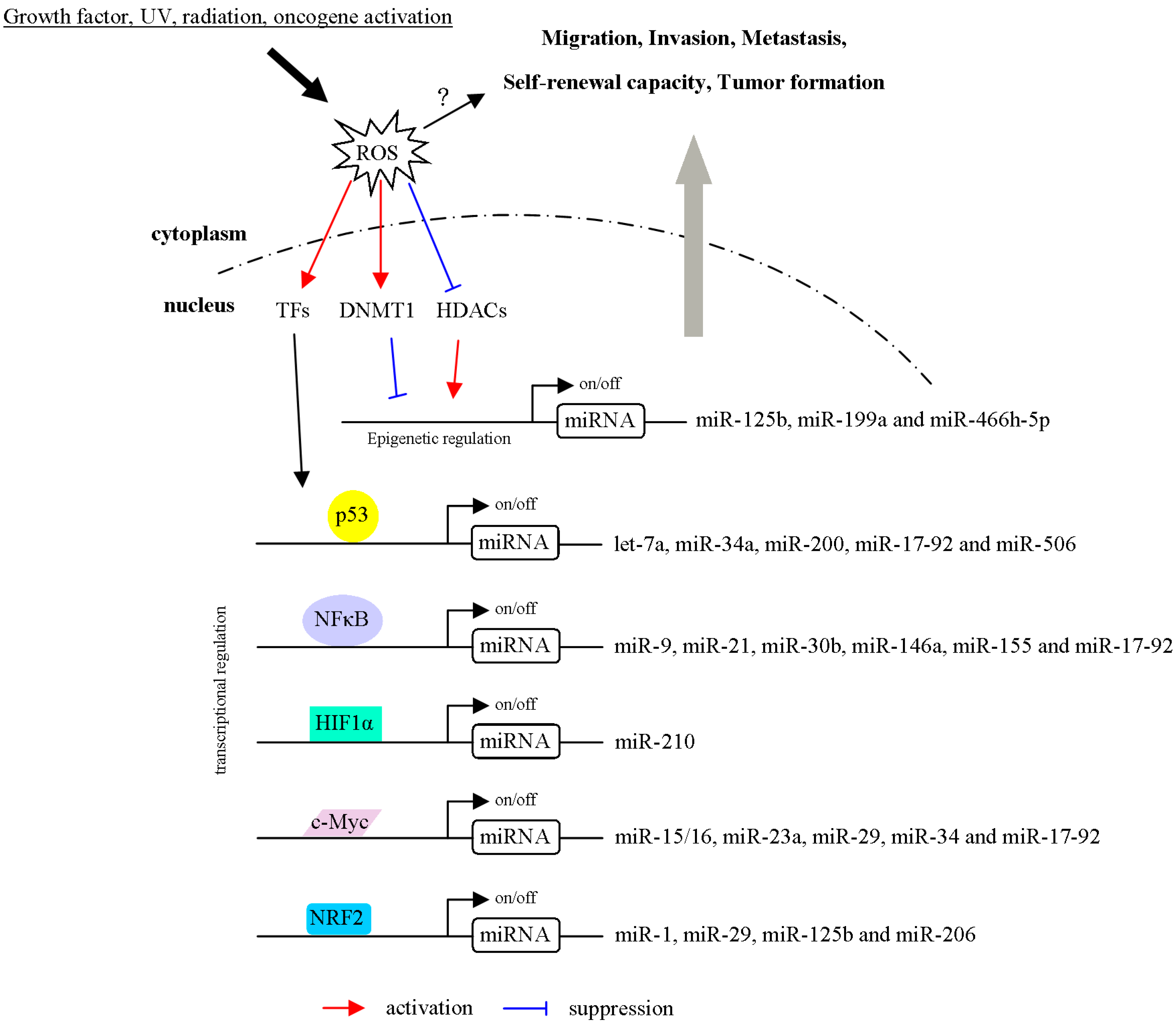
Figure 3.
Schematic model showing mechanisms in which ROS regulates the biogenesis and transcription of miRNAs. ROS activate or inhibit epigenetic, transcriptional regulations of miRNA expression. For example, miRNAs are regulated by ROS through modulation of chromatin remodeling factors (DNMT1 and HDACs). In addition, ROS induces or represses transcriptional factor (p53, NFκB, HIF1α, c-Myc and NRF2) to regulate miRNA expressions. Furthermore, ROS/TF/miRNA axis controls cell migration, invasion, metastasis, self-renewal capacity and tumor formation.
Figure 3.
Schematic model showing mechanisms in which ROS regulates the biogenesis and transcription of miRNAs. ROS activate or inhibit epigenetic, transcriptional regulations of miRNA expression. For example, miRNAs are regulated by ROS through modulation of chromatin remodeling factors (DNMT1 and HDACs). In addition, ROS induces or represses transcriptional factor (p53, NFκB, HIF1α, c-Myc and NRF2) to regulate miRNA expressions. Furthermore, ROS/TF/miRNA axis controls cell migration, invasion, metastasis, self-renewal capacity and tumor formation.


{kind=link}
{kind=link}
{kind=link}
Table 1.
ROS-related miRNAs and their potential mechanisms in cancers.
| miRNA | Regulation Mechanism a | ROS Production b | Expression in Cancer c | Cell/Cancer Types | Molecules, Cellular Processes and Signaling Pathways Involved d | References |
|---|---|---|---|---|---|---|
| Let-7a | OS, p53 | ✓ | Down | CSC, prostate cancer, pancreatic cancer | PTEN, LIN28b | [105,135] |
| miR-1 | NRF2, HDAC4 | ✓ | Down | Non-small cell lung cancer | NRF2, KEAP1, glucose metabolism, tumor growth | [49] |
| miR-15/16 | c-Myc | ✓ | Down | Skin, colon cancer | FGF2, HIF-2α, senescence-like phenotype, angiogenesis, metastasis | [30] |
| miR-21 | Glucose, NFκB, STAT3 | ✓ | Up | CSCs, lung cancer, liver cancer, colorectal cancer | MAPK pathway, cell migration, invasion and EMT phenotype, self-renewal ability | [41,110] |
| miR-23a | c-Myc | ✓ | - | Cardiac disease, myeloma | Glutaminase, MnSOD, apoptosis, cell growth | [136,137,138] |
| miR-29 | c-Myc, H2O2, NRF2 | ✓ | Dual role | Ovarian cancer, lung cancer, lymphoma | SIRT1, senescence, proliferation, apoptosis | [50,139,140,141,142] |
| miR-33a/b | - | ✓ | Down | Liver | HDL biosynthesis, apoptosis, OS resistance | [103] |
| miR-34 | OS, c-Myc, p53 | ✓ | Down | Stromal cells, CSC, bladder cancer, lung cancer | CD44, EMT markers, SIRT1, senescence, metastasis | [33,111,143] |
| miR-17-92 | c-Myc, p53, NFκB | ✓ | Up | Lung cancer, | Vitamin D, Senescence, apoptosis | [120,144,145,146] |
| miR-92a | - | ✓ | Up | Endothelial cells | SIRT1, KLF2, KLF4 | [124,125] |
| miR-125b | DNMT1, H2O2, NRF2 | ✓ | Dual role | Ovarian cancer, liver | Epigenetic regulation | [51,57] |
| miR-181 | - | ✓ | Up | Macrophagy, HCC | SIRT1, insulin sensitivity, NFκB activity, apoptosis | [80] |
| miR-199a | DNMT1, H2O2 | ✓ | Down, (hypermethylation) | Ovarian cancer | HIF1α, SIRT1, Epigenetic regulation | [57,83] |
| miR-200 | P53, H2O2 | ✓ | Down | CSC, breast cancer, liver cancer | Bmil-1, Suz12, Notch-1, self-renewal capacity, EMT markers, senescence | [34,35] |
| miR-210 | Hypoxia | ✓ | Up | CSCs | E-cadherin, Hypoxia, proliferation, self-renewal capacity, migration and invasion, senescence | [65,109] [113] |
| miR-217 | - | - | Dual role | Endothelial cells | SIRT1, Angiogenesis, premature senescence-like phenotype | [123] |
| miR-466h-5p | ROS, HDAC2 | - | - | Mouse ovarian epithelial | BCL2L2, apoptosis | [58] |
| MiR-506 | P53 | ✓ | Down | Lung cancer | NFκB signaling pathway | [44] |
| miR-509 | - | ✓ | Down | Breast cancer | SOD2, Cell growth, migration and invasion | [94] |
a: MiRNAs are regulated by upstream transcriptional factor, ROS or hypoxia, as indicated. -: Information is unavailable. b: ✓: MiRNAs are responsible for producing ROS. -: Information is unavailable. c: Expression level of miRNAs in cancer. Up: upregulated in cancer, Down: downregulated in cancer, Dual role: up- or downregulated in cancer. d: Downstream molecules, signaling pathways and phenotypes involved in miRNA-mediated functions.
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, Y.-H. MicroRNA Networks Modulate Oxidative Stress in Cancer. Int. J. Mol. Sci. 2019, 20, 4497. https://doi.org/10.3390/ijms20184497
AMA Style
Lin Y-H. MicroRNA Networks Modulate Oxidative Stress in Cancer. International Journal of Molecular Sciences. 2019; 20(18):4497. https://doi.org/10.3390/ijms20184497
Chicago/Turabian StyleLin, Yang-Hsiang. 2019. "MicroRNA Networks Modulate Oxidative Stress in Cancer" International Journal of Molecular Sciences 20, no. 18: 4497. https://doi.org/10.3390/ijms20184497
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.