Jasmonic Acid Methyl Ester Induces Xylogenesis and Modulates Auxin-Induced Xylary Cell Identity with NO Involvement

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
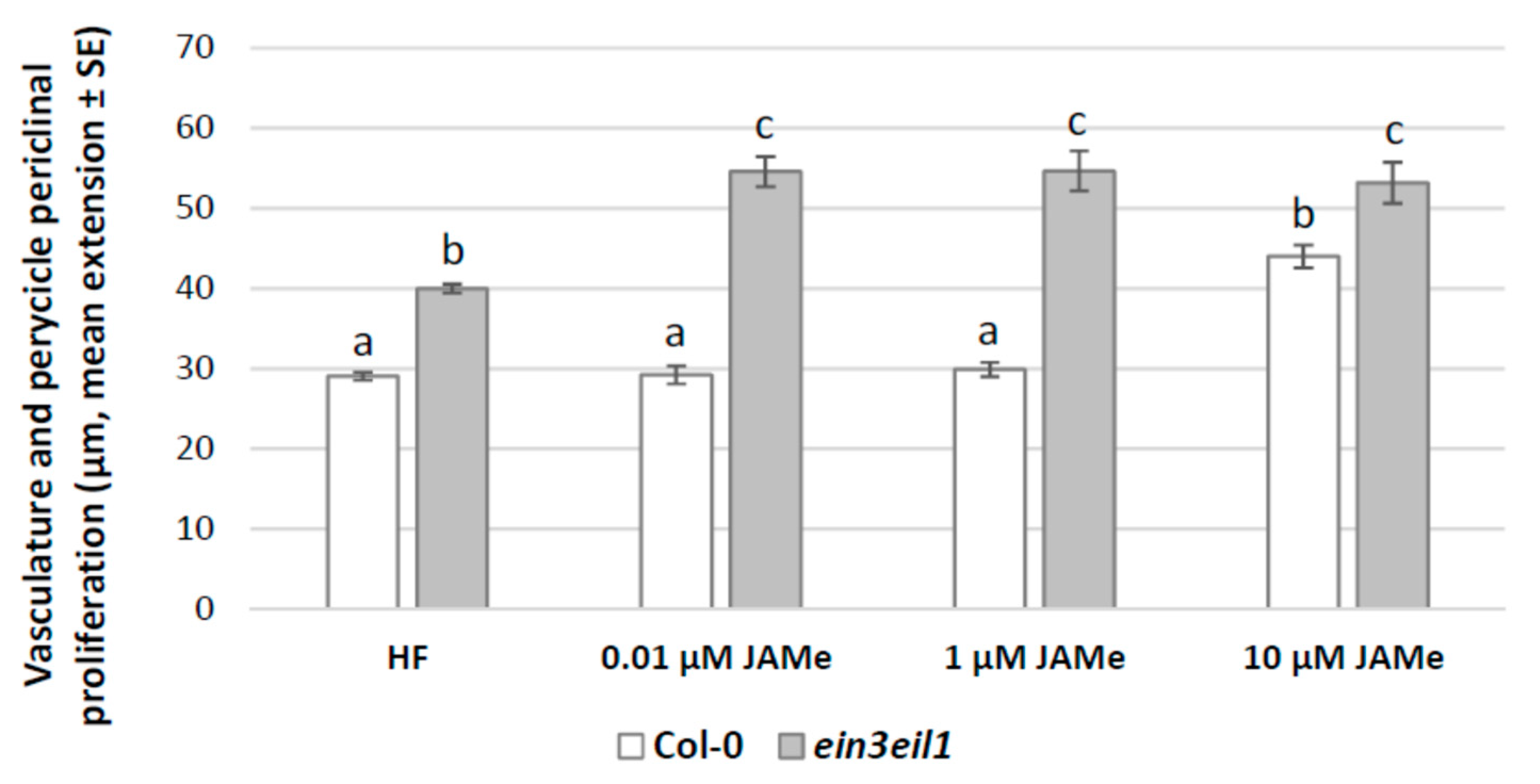
2.1. A Specific Concentration of Exogenous JAMe Enhances Ectopic Protoxylem Formation but its Combination with ACC Reduces it, without Affecting Xylary Identity
2.2. JAMe Changes the Xylary Cell Identity Determined by Exogenous IBA or IAA and Promotes Adventitious Rooting when Applied with IAA/IBA
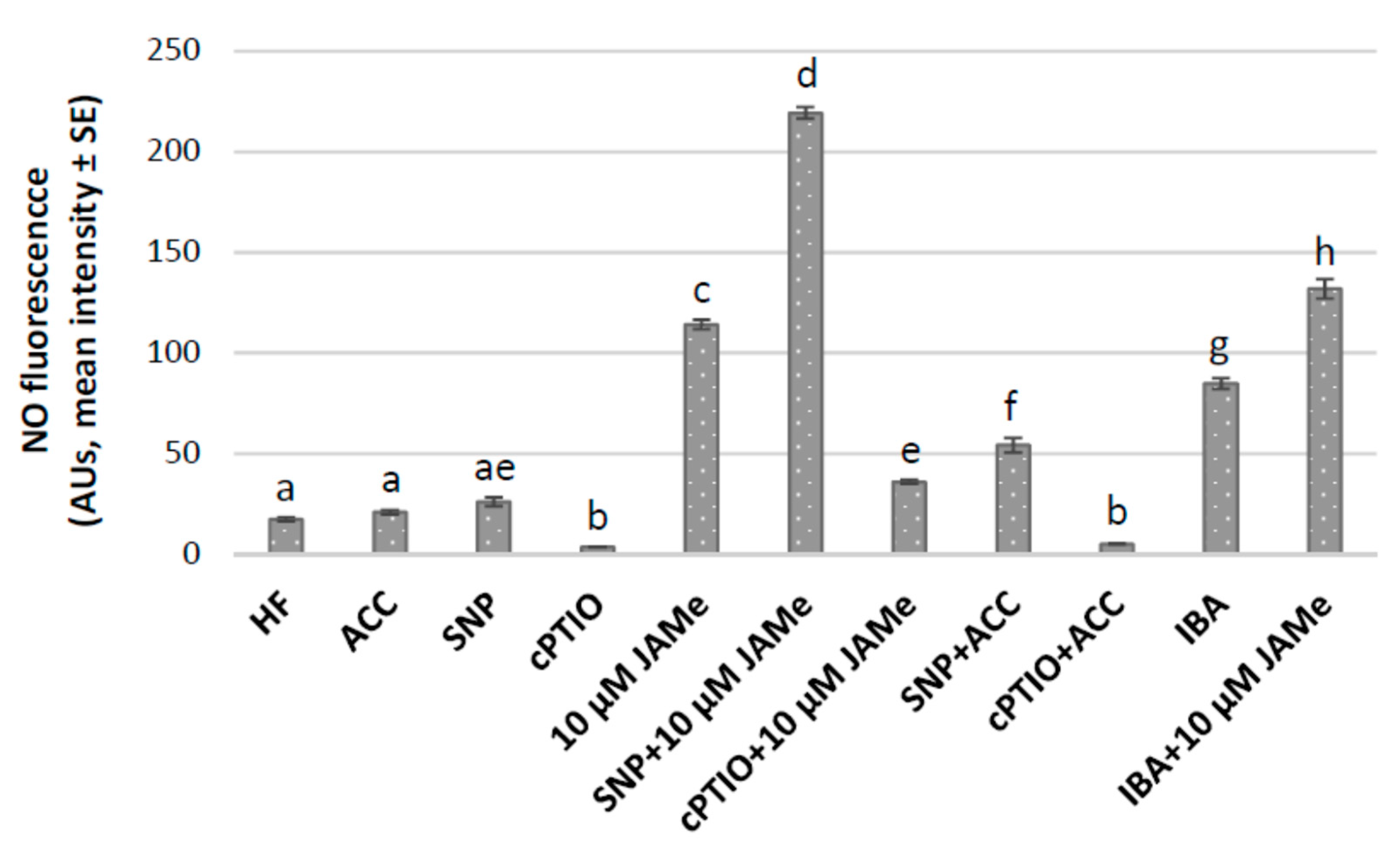
2.3. Nitric Oxide is an Early Marker of Cell Reactivation in the Hypocotyl Basal Pericycle Involved in Either Xylogenesis or Adventitious Rooting, and its Signal is Enhanced by JAMe
3. Discussion
3.1. The Action of JAMe on Xylogenesis is Negatively Affected by ET Signalling by EIN3EIL1
3.2. Nitric Oxide is a Common Marker of JAMe-induced Xylogenesis and Auxin-Induced Adventitious Rooting Acting Downstream to Pericycle Cell Determination to either Program
3.3. The Auxin-Determined Xylary Cell Identity is Modulated by JAMe
4. Materials and Methods
4.1. Plant Growth
4.2. Histological Analysis
4.3. Nitric Oxide Detection
4.4. Measurement Procedures and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | 1-Aminocyclopropane-1-carboxylic acid |
| ANOVA | Analysis of variance |
| AR | Adventitious root |
| ARP | Adventitious root primordium |
| AU | Arbitrary unit |
| cPTIO | 2-4-carboxyphenyl-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide |
| DAF-FMDA | Diaminofluorescein-FM diacetate |
| DAS | Days after stratification |
| ET | Ethylene |
| HF | Hormone-free |
| IAA | Indole-3-acetic acid |
| IBA | Indole-3-butyric acid |
| JA | Jasmonic acid |
| JAMe | Jasmonic acid methyl ester |
| SE | Standard error |
| SNP | Sodium nitroprusside |
| TF | Transcription factor |
| XE | Xylary element |
References
- Yoshida, S.; Iwamoto, K.; Demura, T.; Fukuda, H. Comprehensive analysis of the regulatory roles of auxin in early transdifferentiation into xylem cells. Plant Mol. Biol. 2009, 70, 457–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyashima, S.; Sebastian, J.; Lee, J.Y.; Helariutta, Y. Stem cell function during plant vascular development. EMBO J. 2013, 32, 178–193. [Google Scholar] [CrossRef]
- Fukuda, H. Tracheary element differentiation. Plant Cell 1997, 9, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Falasca, G.; Zaghi, D.; Possenti, M.; Altamura, M.M. Adventitious root formation in Arabidopsis thaliana thin cell layers. Plant Cell Rep. 2004, 23, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Falasca, G.; Altamura, M.M. Histological analysis of adventitious rooting in Arabidopsis thaliana (L.) Heynh seedlings. Plant Biosystems 2003, 137, 265–273. [Google Scholar] [CrossRef]
- Fukuda, H. Signals that control plant vascular cell differentiation. Nat. Rev. Mol. Cell Biol. 2004, 5, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Della Rovere, F.; Fattorini, L.; D’Angeli, S.; Veloccia, A.; Del Duca, S.; Cai, G.; Falasca, G.; Altamura, M.M. Arabidopsis SHR and SCR transcription factors and AUX1 auxin influx carrier control the switch between adventitious rooting and xylogenesis in planta and in in vitro cultured thin cell layers. Ann. Bot. 2015, 115, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, B.; Uggla, C.; Tuominen, H. Cambial growth and auxin gradients. In Cell and Molecular Biology of Wood Formation; Savidge, R.A., Barnett, J.R., Napier, R., Eds.; BIOS Scientific Publishers: Oxford, UK, 2000; pp. 169–188. [Google Scholar]
- McCann, M.C.; Domingo, C.; Stacey, N.J.; Milioni, D.; Roberts, K. Tracheary element formation in an in vitro system. In Cell and Molecular Biology of Wood Formation; Savidge, R.A., Barnett, J.R., Napier, R., Eds.; BIOS Scientific Publishers: Oxford, UK, 2000; pp. 457–470. [Google Scholar]
- Charton, L.; Plett, A.; Linka, N. Plant peroxisomal solute transporter proteins. J. Integrative Plant Biol. 2019, 61, 817–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattorini, L.; Della Rovere, F.; Andreini, E.; Ronzan, M.; Falasca, G.; Altamura, M.M. Indole-3-butyric acid induces ectopic formation of metaxylem in the hypocotyl of Arabidopsis thaliana without conversion into indole-3-acetic acid and with a positive interaction with ethylene. Int. J. Mol. Sci. 2017, 18, 2474. [Google Scholar] [CrossRef]
- Miller, A.R.; Pengelly, W.L.; Roberts, L.W. Introduction of xylem differentiation in Lactuca by ethylene. Plant Physiol. 1984, 75, 1165–1166. [Google Scholar] [CrossRef] [PubMed]
- Pesquet, E.; Tuominen, H. Ethylene stimulates tracheary element differentiation in Zinnia elegans cell cultures. New Phytol. 2011, 190, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Pesquet, E.; Zhang, B.; Gorzsás, A.; Puhakainen, T.; Serk, H.; Escamez, S.; Barbier, O.; Gerber, L.; Courtois-Moreau, C.; Alatalo, E.; et al. Non-cell-autonomous postmortem lignification of tracheary elements in Zinnia elegans. Plant Cell 2013, 25, 1314–1328. [Google Scholar] [CrossRef] [PubMed]
- Biondi, S.; Scaramagli, S.; Capitani, F.; Marino, G.; Altamura, M.M.; Torrigiani, P. Ethylene involvement in vegetative bud formation in tobacco thin layers. Protoplasma 1998, 202, 134–144. [Google Scholar] [CrossRef]
- Eklund, L.; Tiltu, A. Cambial activity in ‘normal’ spruce Picea abies Karst (L.) and snake spruce Picea abies (L.) Karst f. virgata (Jacq.) Rehd. in response to ethylene. J. Exp. Bot. 1999, 50, 1489–1493. [Google Scholar] [CrossRef]
- Love, J.; Björklund, S.; Vahala, J.; Hertzberg, M.; Kangasjärvi, J.; Sundberg, B. Ethylene is an endogenous stimulator of cell division in the cambial meristem of Populus. Proc. Natl. Acad. Sci. USA 2009, 106, 5984–5989. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yu, J.; Wen, C.K. An alternate route of ethylene receptor signaling. Front. Plant Sci. 2014, 5, 648. [Google Scholar] [CrossRef] [PubMed]
- Veloccia, A.; Fattorini, L.; Della Rovere, F.; Sofo, A.; D’Angeli, S.; Betti, C.; Falasca, G.; Altamura, M.M. Ethylene and auxin interaction in the control of adventitious rooting in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6445–6458. [Google Scholar] [CrossRef] [PubMed]
- Růžička, K.; Ljung, K.; Vanneste, S.; Podhorsk, R.; Beeckman, T.; Friml, J.; Benková, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell. 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z. Molecular basis for jasmonate and ethylene signal interactions in Arabidopsis. J. Exp. Bot. 2014, 65, 5743–5748. [Google Scholar] [CrossRef] [PubMed]
- Sehr, E.M.; Agusti, J.; Lehner, R.; Farmer, E.E.; Schwarz, M.; Greb, T. Analysis of secondary growth in the Arabidopsis shoot reveals a positive role of jasmonate signalling in cambium formation. Plant J. 2010, 63, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, L.; Falasca, G.; Kevers, C.; Rocca, L.M.; Zadra, C.; Altamura, M.M. Adventitious rooting is enhanced by methyl jasmonate in tobacco thin cell layers. Planta 2009, 231, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Chang, S.H.; Um, T.Y.; Lee, S.; Kim, J.K.; Choi, Y.D. Antagonistic interaction between jasmonic acid and cytokinin in xylem development. Sci. Rep. 2017, 7, 10212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, G.; Lee, S.; Chang, S.H.; Kim, J.-K.; Choi, Y.D. Jasmonic acid modulates xylem development by controlling polar auxin transport in vascular tissues. Plant Biotechnol. Rep. 2018, 12, 265–271. [Google Scholar] [CrossRef]
- Fattorini, L.; Hause, B.; Gutierrez, L.; Veloccia, A.; Della Rovere, F.; Piacentini, D.; Falasca, G.; Altamura, M.M. Jasmonate promotes auxin-induced adventitious rooting in dark-grown Arabidopsis thaliana seedlings and stem thin cell layers by a cross-talk with ethylene signalling and a modulation of xylogenesis. BMC Plant Biol. 2018, 18, 182. [Google Scholar] [CrossRef] [PubMed]
- Freschi, L. Nitric oxide and phytohormone interactions: Current status and perspectives. Front Plant Sci. 2013, 4, 398. [Google Scholar] [CrossRef] [PubMed]
- Neill, S. NO way to die – nitric oxide, programmed cell death and xylogenesis. New Phytol. 2005, 165, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Gabaldón, C.; Gómez, R.L.V.; Pedreño, M.A.; Ros, B.A. Nitric oxide production by the differentiating xylem of Zinnia elegans. New Phytol. 2005, 165, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Feng, F.; Liu, J.; Zhao, Q. Nitric oxide affects rice root growth by regulating auxin transport under nitrate supply. Front. Plant Sci. 2018, 9, 659. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.H.; Guo, F.Q. Nitric oxide regulates dark-induced leaf senescence through EIN2 in Arabidopsis. J. Integr. Plant Biol. 2012, 54, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.M.; Stepanova, A.N. The ethylene signalling pathway. Science 2004, 306, 1513–1515. [Google Scholar] [CrossRef]
- Iakimova, E.T.; Woltering, E.J. Xylogenesis in zinnia (Zinnia elegans) cell cultures: Unravelling the regulatory steps in a complex developmental programmed cell death event. Planta 2017, 245, 681–705. [Google Scholar] [CrossRef] [PubMed]
- Bagniewska-Zadworna, A.; Arasimowicz-Jelonek, M.; Smoliński, D.J.; Stelmasik, A. New insights into pioneer root xylem development: Evidence obtained from Populus trichocarpa plants grown under field conditions. Ann. Bot. 2014, 113, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, L.; Veloccia, A.; Della Rovere, F.; D’Angeli, S.; Falasca, G.; Altamura, M.M. Indole-3-butyric acid promotes adventitious rooting in Arabidopsis thaliana thin cell layers by conversion into indole-3-acetic acid and stimulation of anthranilate synthase activity. BMC Plant Biol. 2017, 17, 121. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Barroso, J.B. Peroxisomal plant metabolism – an update on nitric oxide, Ca2+ and the NADPH recycling network. J. Cell Sci. 2018, 131, jcs202978. [Google Scholar] [CrossRef]
- Chini, A.; Monte, I.; Zamarreño, A.M.; Hamberg, M.; Lassueur, S.; Reymond, P.; Weiss, S.; Stintzi, A.; Schaller, A.; Porzel, A.; et al. An OPR3-independent pathway uses 4,5-didehydrojasmonate for jasmonate synthesis. Nat. Chem. Biol. 2018, 14, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.W.; Wu, J.Y. Nitric oxide is involved in methyl jasmonate-induced defense responses and secondary metabolism activities of Taxus cells. Plant Cell Physiol. 2005, 46, 923–930. [Google Scholar] [CrossRef]
- Zhu, Z.; An, F.; Feng, Y.; Li, P.; Xue, L.; Angelo, A.M.; Jiang, Z.; Kim, J.M.; To, T.K.; Li, W.; et al. Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 12539–12544. [Google Scholar] [CrossRef]
- Della Rovere, F.; Fattorini, L.; D’Angeli, S.; Veloccia, A.; Falasca, G.; Altamura, M.M. Auxin and cytokinin control formation of the quiescent centre in the adventitious root apex of Arabidopsis. Ann. Bot. 2013, 112, 1395–1407. [Google Scholar] [CrossRef]
- Betti, C.; Della Rovere, F.; Ronzan, M.; Fattorini, L. EIN2 and COI1 control the antagonism between ethylene and jasmonate in adventitious rooting of Arabidopsis thaliana thin cell layers. PCTOC 2019, 138, 41–51. [Google Scholar] [CrossRef]
- Tiwari, S.B.; Hagen, G.; Guilfoyle, T. The roles of auxin response factor domains in auxin-responsive transcription. Plant Cell 2003, 15, 533–543. [Google Scholar] [CrossRef]
- Song, S.; Huang, H.; Gao, H.; Wang, J.; Wu, D.; Liu, X.; Yang, S.; Zhai, Q.; Li, C.; Qi, T.; et al. Interaction between MYC2 and ETHYLENE INSENSITIVE3 modulates antagonism between jasmonate and ethylene signaling in Arabidopsis. Plant Cell 2014, 26, 263–279. [Google Scholar] [CrossRef]
- Yemets, A.I.; Krasylenko, Y.A.; Sheremet, Y.A.; Blume, Y.B. Microtubule reorganization as a response to implementation of NO signals in plant cells. Cytol. Genet. 2009, 43, 73–79. [Google Scholar] [CrossRef]
- Jovanović, A.M.; Durst, S.; Nick, P. Plant cell division is specifically affected by nitrotyrosine. J. Exp. Bot. 2010, 61, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Nakamura, Y.; Mori, I.C.; Murata, Y. Nitric oxide functions in both methyl jasmonate signalling and abscisic acid signalling in Arabidopsis guard cells. Plant Signal. Behav. 2009, 4, 119–120. [Google Scholar] [CrossRef]
- Zhou, J.; Jia, F.; Shao, S.; Zhang, H.; Li, G.; Xia, X.; Zhou, Y.; Yu, J.; Shi, K. Involvement of nitric oxide in the jasmonate-dependent basal defense against root-knot nematode in tomato plants. Front. Plant Sci. 2015, 6, 193. [Google Scholar] [CrossRef] [PubMed]
- Ötvös, K.; Pasternak, T.P.; Miskolczi, P.; Domoki, M.; Dorjgotov, D.; Szűcs, A.; Bottka, S.; Dudits, D.; Fehér, A. Nitric oxide is required for, and promotes auxin-mediated activation of cell division and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell cultures. Plant J. 2005, 43, 849–860. [Google Scholar] [CrossRef]
- Sun, J.; Xu, Y.; Ye, S.; Jiang, H.; Chen, Q.; Liu, F.; Zhou, W.; Chen, R.; Li, X.; Tietz, O.; et al. Arabidopsis ASA1 is important for jasmonate-mediated regulation of auxin biosynthesis and transport during lateral root formation. Plant Cell 2009, 21, 1495–1511. [Google Scholar] [CrossRef] [PubMed]
- Bishopp, A.; Help, H.; El-Showk, S.; Weijers, D.; Scheres, B.; Friml, J.; Benková, E.; Mähönen, A.P.; Helariutta, Y. A mutually inhibitory interaction between auxin and cytokinin specifies vascular pattern in roots. Curr. Biol. 2011, 21, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H. Signaling, transcriptional regulation, and asynchronous pattern formation governing plant xylem development. Proc Jpn. Acad. Ser. B Phys. Biol. Sci. 2016, 92, 98–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, M.; Udagawa, M.; Nishikubo, N.; Horiguchi, G.; Yamaguchi, M.; Ito, J.; Mimura, T.; Fukuda, H.; Demura, T. Transcription switches for protoxylem and metaxylem vessel formation. Gene Dev. 2005, 19, 1855–1860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Kubo, M.; Fukuda, H.; Demura, T. VASCULAR-RELATED NAC-DOMAIN7 is involved in the differentiation of all types of xylem vessels in Arabidopsis roots and shoots. Plant J. 2008, 55, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Moubayidin, L.; Di Mambro, R.; Sozzani, R.; Pacifici, E.; Salvi, E.; Terpstra, I.; Bao, D.; van Dijken, A.; Dello Ioio, R.; Perilli, S.; et al. Spatial coordination between stem cell activity and cell differentiation in the root meristem. Dev. Cell 2013, 26, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Carlsbecker, A.; Lee, J.Y.; Roberts, C.J.; Dettmer, J.; Lehesranta, S.; Zhou, J.; Lindgren, O.; Moreno-Risueno, M.A.; Vatén, A.; Thitamadee, S.; et al. Cell signalling by microRNA165/6 directs gene dose-dependent root cell fate. Nature 2010, 465, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etchells, J.P.; Smit, M.E.; Gaudinier, A.; Williams, C.J.; Brady, S.M. A brief history of the TDIF-PXY signalling module: Balancing meristem identity and differentiation during vascular development. New Phytol. 2016, 209, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hirayama, T.; Fujishige, N.; Kunii, T.; Nishimura, N.; Iuchi, S.; Shinozaki, K. A novel ethanol-hypersensitive mutant of Arabidopsis. Plant Cell Physiol. 2004, 45, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.W.; Yang, J.L.; Qin, C.; Jin, C.W.; Mo, J.H.; Ye, T.; Zheng, S.J. Nitric oxide acts downstream of auxin to trigger root ferric-chelate reductase activity in response to iron deficiency in Arabidopsis. Plant Physiol. 2010, 154, 810–819. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Col-0 Ectopic XEs Mean Number (± SE) | ein3eil1 Ectopic XEs Mean Number (± SE) | Col-0 Ectopic Protoxylem % | Col-0 Ectopic Metaxylem % | ein3eil1 Ectopic Protoxylem % | ein3eil1 Ectopic Metaxylem % |
|---|---|---|---|---|---|---|
| HF (control) | 0.5 ± 0.2 a | 0.6 ± 0.3 a | 100 | 0 | 100 | 0 |
| 0.01 µM JAMe | 1.3 ± 0.3 a | 2.3 ± 0.7 b | 100 | 0 | 100 | 0 |
| 1 µM JAMe | 1.0 ± 0.3 a | 2.4 ± 0.7 b | 100 | 0 | 100 | 0 |
| 10 µM JAMe | 2.8 ± 0.3 b | 2.5 ± 0.4 b | 100 | 0 | 100 | 0 |
| 0.1 µM ACC | 1.1 ± 0.3 a | 0.6 ± 0.2 a | 45.5 | 54.5 | 100 | 0 |
| 10 µM JAMe + ACC | 0.8 ± 0.2 a | 2.4 ± 0.3 b | 100 | 0 | 100 | 0 |
| 10 µM IBA | 4.4 ± 0.6 b | 2.0 ± 0.1 b | 0 | 100 | 100 | 0 |
| IBA + 0.01 µM JAMe | 3.5 ± 0.4 b | 2.4 ± 0.3 b | 17.6 | 82.4 | 20.7 | 79.3 |
| IBA + 1 µM JAMe | 3.7 ± 0.8 b | 2.3 ± 0.4 b | 22.7 | 77.3 | 22.2 | 77.8 |
| IBA + 10 µM JAMe | 4.0 ± 0.6 b | 3.6 ± 0.2 b | 61.9 | 38.1 | 50 | 50 |
| 10 µM IAA | 2.8 ± 0.5 b | 2.0 ± 0.4 b | 100 | 0 | 100 | 0 |
| IAA + 0.01 µM JAMe | 2.5 ± 0.4 b | 1.8 ± 0.4 b | 100 | 0 | 35 | 65 |
| IAA + 1 µM JAMe | 2.5 ± 0.2 b | 2.2 ± 0.3 b | 33.3 | 66.7 | 25 | 75 |
| IAA + 10 µM JAMe | 2.6 ± 0.5 b | 3.8 ± 0.8 b | 7.7 | 92.3 | 9 | 91 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Rovere, F.; Fattorini, L.; Ronzan, M.; Falasca, G.; Altamura, M.M.; Betti, C. Jasmonic Acid Methyl Ester Induces Xylogenesis and Modulates Auxin-Induced Xylary Cell Identity with NO Involvement. Int. J. Mol. Sci. 2019, 20, 4469. https://doi.org/10.3390/ijms20184469
Della Rovere F, Fattorini L, Ronzan M, Falasca G, Altamura MM, Betti C. Jasmonic Acid Methyl Ester Induces Xylogenesis and Modulates Auxin-Induced Xylary Cell Identity with NO Involvement. International Journal of Molecular Sciences. 2019; 20(18):4469. https://doi.org/10.3390/ijms20184469
Chicago/Turabian StyleDella Rovere, Federica, Laura Fattorini, Marilena Ronzan, Giuseppina Falasca, Maria Maddalena Altamura, and Camilla Betti. 2019. "Jasmonic Acid Methyl Ester Induces Xylogenesis and Modulates Auxin-Induced Xylary Cell Identity with NO Involvement" International Journal of Molecular Sciences 20, no. 18: 4469. https://doi.org/10.3390/ijms20184469