Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System

Abstract
1. Introduction
2. Results
2.1. Vector Construction, Antigen Expression in Sf9 Cells, and Western Blot Identification
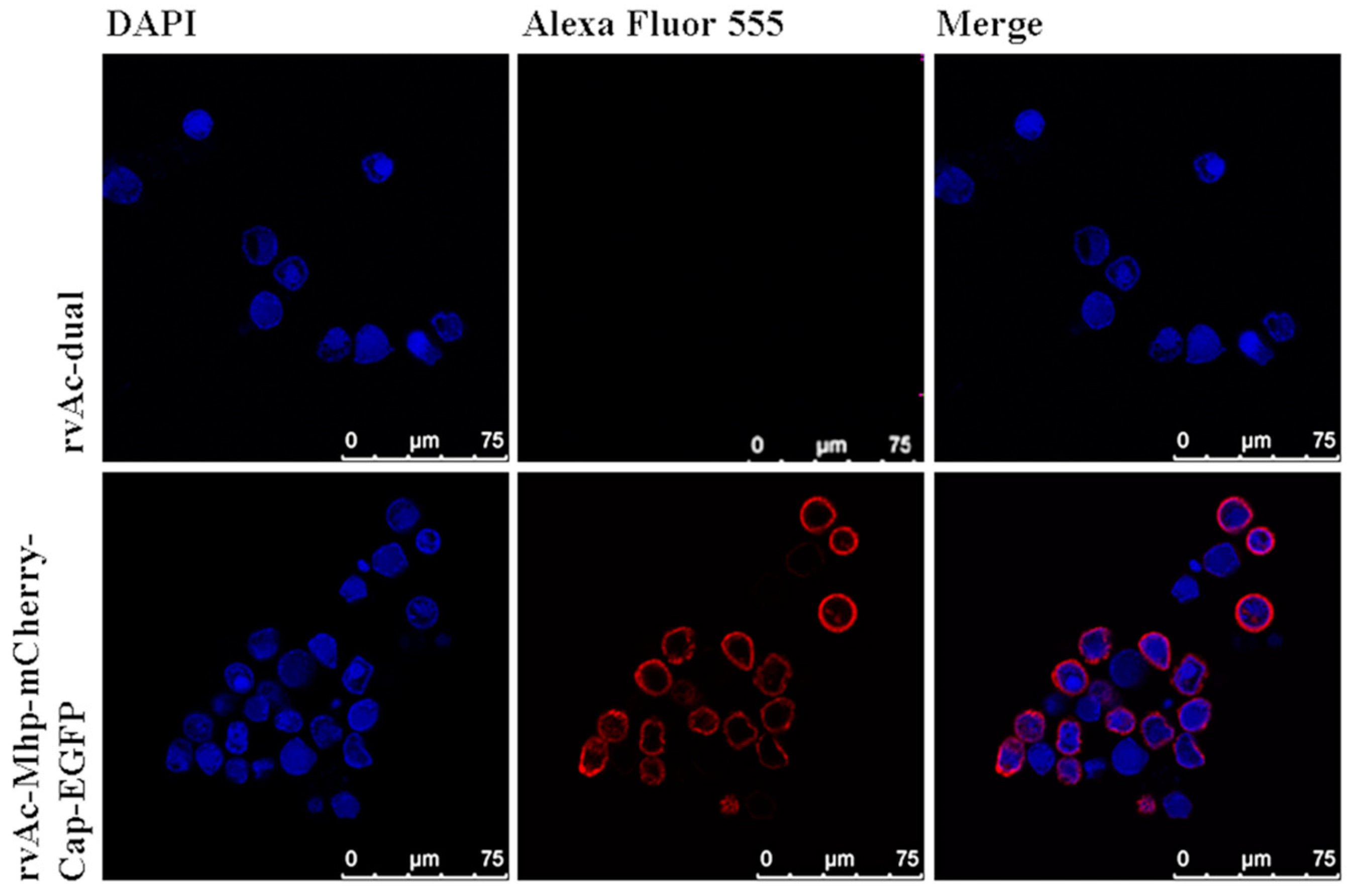
2.2. Localization of Antigen Expression in Cells and Recombinant Baculovirus rvAc-P97R1P46P42-mCherry-Cap-EGFP
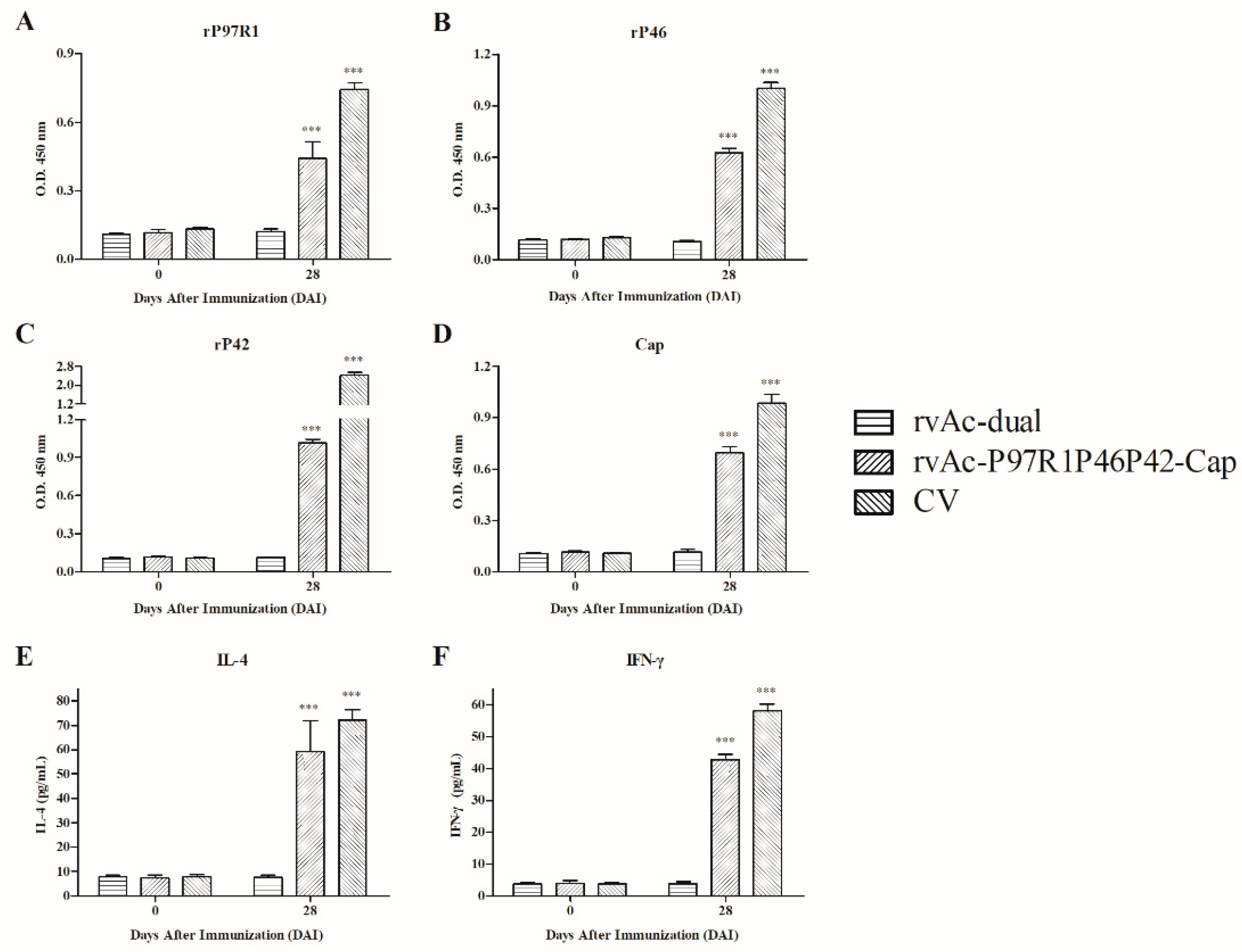
2.3. Systemic Humoral and Cellular Immune Response of rvAc-P97R1P46P42-Cap Induction in Mice
2.4. Immunogenicity in Piglets
3. Discussion
4. Materials and Methods
4.1. Selection of Coding Sequences and Gene Design and Construction of the Baculovirus Expression Vector
4.2. Culture of Spodoptera Frugiperda Cells (Sf9) and Preparation of Pathogens (PCV2 and Mhp)
4.3. Preparation of Recombinant Bacmid DNA and Proliferation of the Recombinant Baculovirus
4.4. Indirect Immunofluorescence and Immunogold Electron Microscopy Assays
4.5. Transformation of the Vector and Baculovirus for Animal Immune Experiments
4.6. Mouse Vaccination
4.7. IgG Antibody Analysis
4.8. Lymphocyte Proliferation Assay
4.9. Reactivity with Native Proteins
4.10. Piglet Vaccination
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chaves, L.C.S.; Ribeiro, B.M.; Blissard, G.W. Production of GP64-free virus-like particles from baculovirus-infected insect cells. J. Gen. Virol. 2018, 99, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Zhang, T.; Zhang, Y.; Wang, H.; Deng, F. Zika virus baculovirus-expressed virus-Like particles induce neutralizing antibodies in mice. Virol. Sin. 2018, 33, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Premanand, B.; Zhong Wee, P.; Prabakaran, M. Baculovirus surface display of immunogenic proteins for vaccine development. Viruses 2018, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Chen, X.; Ouyang, H. Interactions of porcine circovirus 2 with its hosts. Virus Genes 2016, 52, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Ni, B.; Liu, M.; Feng, Z.; Xiong, Q.; Xiao, S.; Shao, G. Mycoplasma hyopneumoniae-derived lipid-associated membrane proteins induce apoptosis in porcine alveolar macrophage via increasing nitric oxide production, oxidative stress, and caspase-3 activation. Vet. Immunol. Immunopathol. 2013, 155, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shu, J.; Chen, J.; Wu, Y.; He, Y. A concise review of vaccines against Mycoplasma hyopneumoniae. Res Vet Sci 2019, 123, 144–152. [Google Scholar] [CrossRef]
- Chen, A.Y.; Fry, S.R.; Forbes-Faulkner, J.; Daggard, G.; Mukkur, T.K. Evaluation of the immunogenicity of the P97R1 adhesin of Mycoplasma hyopneumoniae as a mucosal vaccine in mice. J. Med. Microbiol. 2006, 55, 923–929. [Google Scholar] [CrossRef]
- De Oliveira, N.R.; Jorge, S.; Gomes, C.K.; Rizzi, C.; Pacce, V.D.; Collares, T.F.; Monte, L.G.; Dellagostin, O.A. A novel chimeric protein composed of recombinant Mycoplasma hyopneumoniae antigens as a vaccine candidate evaluated in mice. Vet. Microbiol. 2017, 201, 146–153. [Google Scholar] [CrossRef]
- Galli, V.; Simionatto, S.; Marchioro, S.B.; Fisch, A.; Gomes, C.K.; Conceicao, F.R.; Dellagostin, O.A. Immunisation of mice with Mycoplasma hyopneumoniae antigens P37, P42, P46 and P95 delivered as recombinant subunit or DNA vaccines. Vaccine 2012, 31, 135–140. [Google Scholar] [CrossRef]
- Jorge, S.; de Oliveira, N.R.; Marchioro, S.B.; Fisch, A.; Gomes, C.K.; Hartleben, C.P.; Conceicao, F.R.; Dellagostin, O.A. The Mycoplasma hyopneumoniae recombinant heat shock protein P42 induces an immune response in pigs under field conditions. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Nawagitgul, P.; Morozov, I.; Bolin, S.R.; Harms, P.A.; Sorden, S.D.; Paul, P.S. Open reading frame 2 of porcine circovirus type 2 encodes a major capsid protein. J. Gen. Virol. 2000, 81, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Bucarey, S.A.; Noriega, J.; Reyes, P.; Tapia, C.; Saenz, L.; Zuniga, A.; Tobar, J.A. The optimized capsid gene of porcine circovirus type 2 expressed in yeast forms virus-like particles and elicits antibody responses in mice fed with recombinant yeast extracts. Vaccine 2009, 27, 5781–5790. [Google Scholar] [CrossRef] [PubMed]
- Nawagitgul, P.; Harms, P.A.; Morozov, I.; Thacker, B.J.; Sorden, S.D.; Lekcharoensuk, C.; Paul, P.S. Modified indirect porcine circovirus (PCV) type 2-based and recombinant capsid protein (ORF2)-based enzyme-linked immunosorbent assays for detection of antibodies to PCV. Clin. Diagn. Lab. Immunol. 2002, 9, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.G.; Wang, Z.S.; Zhang, Q.; Li, Z.C.; Ding, L.; Li, W.; Wu, H.Y.; Chang, C.D.; Lee, L.H.; Tong, D.W.; et al. Baculovirus as a PRRSV and PCV2 bivalent vaccine vector: baculovirus virions displaying simultaneously GP5 glycoprotein of PRRSV and capsid protein of PCV2. J. Virol. Methods 2012, 179, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Duivon, D.; Corrégé, I.; Hémonic, A.; Rigaut, M.; Roudaut, D.; Jolie, R. Field evaluation of piglet vaccination with a Mycoplasma hyopneumoniae bacterin as compared to a ready-to-use product including porcine circovirus 2 and M. hyopneumoniae in a conventional French farrow-to-finish farm. Porcine Health Management 2018, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Tassis, P.D.; Tsakmakidis, I.; Papatsiros, V.G.; Koulialis, D.; Nell, T.; Brellou, G.; Tzika, E.D. A randomized controlled study on the efficacy of a novel combination vaccine against enzootic pneumonia (Mycoplasma hyopneumoniae) and porcine Circovirus type 2 (PCV2) in the presence of strong maternally derived PCV2 immunity in pigs. BMC. Vet. Res. 2017, 13, 91. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, A.; Russell, M.; Larocque, L.; Gravel, C.; Li, C.; Chen, W.; Cyr, T.; Lavoie, J.R.; Farnsworth, A.; Rosu-Myles, M.; et al. Targeting CD40 enhances antibody- and CD8-mediated protection against respiratory syncytial virus infection. Sci. Rep. 2018, 8, 16648. [Google Scholar] [CrossRef]
- Li, D.; Huang, Y.; Du, Q.; Wang, Z.; Chang, L.; Zhao, X.; Tong, D. CD40 ligand and GMCSF coexpression enhance the immune responses and protective efficacy of PCV2 adenovirus vaccine. Viral. Immunol. 2016, 29, 148–158. [Google Scholar] [CrossRef]
- Contreras-Gomez, A.; Sanchez-Miron, A.; Garcia-Camacho, F.; Molina-Grima, E.; Chisti, Y. Protein production using the baculovirus-insect cell expression system. Biotechnol. Progr. 2014, 30, 1–18. [Google Scholar] [CrossRef]
- Grabherr, R.; Ernst, W. Baculovirus for eukaryotic protein display. Curr. Gene. Ther. 2010, 10, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Chung, Y.C.; Hu, Y.C. Update on baculovirus as an expression and/or delivery vehicle for vaccine antigens. Expert Rev. Vaccines 2014, 13, 1501–1521. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Park, C.; Choi, K.; Chae, C. A new single-dose bivalent vaccine of porcine circovirus type 2 and Mycoplasma hyopneumoniae elicits protective immunity and improves growth performance under field conditions. Vet. Microbiol. 2016, 182, 178–186. [Google Scholar] [CrossRef] [PubMed]
- VanCott, J.L.; Staats, H.F.; Pascual, D.W.; Roberts, M.; Chatfield, S.N.; Yamamoto, M.; Coste, M.; Carter, P.B.; Kiyono, H.; McGhee, J.R. Regulation of mucosal and systemic antibody responses by T helper cell subsets, macrophages, and derived cytokines following oral immunization with live recombinant Salmonella. J. Immunol. 1996, 156, 1504–1514. [Google Scholar] [PubMed]
- Magram, J.; Connaughton, S.E.; Warrier, R.R.; Carvajal, D.M.; Wu, C.Y.; Ferrante, J.; Stewart, C.; Sarmiento, U.; Faherty, D.A.; Gately, M.K. IL-12-deficient mice are defective in IFN gamma production and type 1 cytokine responses. Immun. 1996, 4, 471–481. [Google Scholar] [CrossRef]
- Conceicao, F.R.; Moreira, A.N.; Dellagostin, O.A. A recombinant chimera composed of R1 repeat region of Mycoplasma hyopneumoniae P97 adhesin with Escherichia coli heat-labile enterotoxin B subunit elicits immune response in mice. Vaccine 2006, 24, 5734–5743. [Google Scholar] [CrossRef] [PubMed]
- Fisch, A.; Marchioro, S.B.; Gomes, C.K.; Galli, V.; de Oliveira, N.R.; Simionatto, S.; Dellagostin, O.A.; Mendonça, M.; Moreira, Â.N.; Conceição, F.R. Commercial bacterins did not induce detectable levels of antibodies in mice against Mycoplasma hyopneumoniae antigens strongly recognized by swine immune system. Trials in Vaccinology 2016, 5, 32–37. [Google Scholar] [CrossRef][Green Version]
- Simionatto, S.; Marchioro, S.B.; Galli, V.; Brum, C.B.; Klein, C.S.; Rebelatto, R.; Silva, E.F.; Borsuk, S.; Conceicao, F.R.; Dellagostin, O.A. Immunological characterization of Mycoplasma hyopneumoniae recombinant proteins. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kang, I.; Kim, S.; Park, K.H.; Park, C.; Chae, C. Comparison of 3 vaccination strategies against porcine reproductive and respiratory syndrome virus, Mycoplasma hyopneumoniae, and porcine circovirus type 2 on a 3 pathogen challenge model. Can. J. Vet. Res. 2018, 82, 39–47. [Google Scholar] [PubMed]
- Park, C.; Kang, I.; Seo, H.W.; Jeong, J.; Choi, K.; Chae, C. Comparison of 2 commercial single-dose Mycoplasma hyopneumoniae vaccines and porcine reproductive and respiratory syndrome virus (PRRSV) vaccines on pigs dually infected with M. hyopneumoniae and PRRSV. Can. J. Vet. Res. 2016, 80, 112–123. [Google Scholar] [PubMed]
- Beffort, L.; Weiss, C.; Fiebig, K.; Jolie, R.; Ritzmann, M.; Eddicks, M. Field study on the safety and efficacy of intradermal versus intramuscular vaccination against Mycoplasma hyopneumoniae. Vet. Rec. 2017, 181, 348. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Cheng, X.; Zhang, J.; Tong, T.; Lin, W.; Liao, M.; Fan, H. Induction of robust immunity response in mice by dual-expression-system-based recombinant baculovirus expressing the capsid protein of porcine circovirus type 2. Virol. J. 2013, 10, 316. [Google Scholar] [CrossRef] [PubMed]
- Tischer, I.; Peters, D.; Rasch, R.; Pociuli, S. Replication of porcine circovirus: induction by glucosamine and cell cycle dependence. Arch. Virol. 1987, 96, 39–57. [Google Scholar] [CrossRef] [PubMed]
- Friis, N.F. Some recommendations concerning primary isolation of Mycoplasma suipneumoniae and Mycoplasma flocculare a survey. Nord. Veterinaermed. 1975, 27, 337–339. [Google Scholar]
- Abe, T.; Takahashi, H.; Hamazaki, H.; Miyano-Kurosaki, N.; Matsuura, Y.; Takaku, H. Baculovirus induces an innate immune response and confers protection from lethal influenza virus infection in mice. J. Immunol. 2003, 171, 1133–1139. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, S.; Chae, C.; Ryu, D.Y. A recombinant chimera comprising the R1 and R2 repeat regions of M. hyopneumoniae P97 and the N-terminal region of A. pleuropneumoniae ApxIII elicits immune responses. BMC Vet. Res. 2014, 10, 43. [Google Scholar] [CrossRef] [PubMed]
- Virginio, V.G.; Gonchoroski, T.; Paes, J.A.; Schuck, D.C.; Zaha, A.; Ferreira, H.B. Immune responses elicited by Mycoplasma hyopneumoniae recombinant antigens and DNA constructs with potential for use in vaccination against porcine enzootic pneumonia. Vaccine 2014, 32, 5832–5838. [Google Scholar] [CrossRef]
- Marchioro, S.B.; Simionatto, S.; Galli, V.; Conceicao, F.R.; Brum, C.B.; Fisch, A.; Gomes, C.K.; Dellagostin, O.A. Production and characterization of recombinant transmembrane proteins from Mycoplasma hyopneumoniae. Vet. Microbiol. 2012, 155, 44–52. [Google Scholar] [CrossRef]
- Balasch, M.; Fort, M.; Taylor, L.P.; Diaz, I.; Mateu, E.; Calvert, J.G. Immune response development after vaccination of 1-day-old naive pigs with a Porcine Reproductive and Respiratory Syndrome 1-based modified live virus vaccine. Porcine Health Management 2019, 5, 2. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Y.; Wang, Z.; Bai, J.; Jia, S.; Feng, B.; Jiang, Y.; Cui, W.; Tang, L.; Li, Y.; et al. Oral immunization of mice with a probiotic Lactobacillus casei constitutively expressing the alpha-toxoid induces protective immunity against Clostridium perfringens alpha-toxin. Virulence 2019, 10, 166–179. [Google Scholar] [CrossRef]
- Talaat, K.R.; Halsey, N.A.; Cox, A.B.; Coles, C.L.; Durbin, A.P.; Ramakrishnan, A.; Bream, J.H. Rapid changes in serum cytokines and chemokines in response to inactivated influenza vaccination. Influenza Other Respir. Viruses 2018, 12, 202–210. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | NCBI Accession Number | Features/Function | Original Amino Acid Length (aa) | Selected Fragment (aa) |
|---|---|---|---|---|
| Mhp P97R1 | ADQ90328.1 | Cilium adhesin | 1082 | 788–915 |
| Mhp P46 | ADQ90718.1 | 46 kDa surface antigen | 120 | 323–419 |
| Mhp P42 | ADQ90292.1 | Chaperone protein DnaK | 622 | 434–600 |
| PCV2 Cap | ARW74078.1 | Capsid protein | 234 | 42–234 |
| Group | Formulation | Immunization Time Points (DAI) | Dose |
|---|---|---|---|
| 1, PBS | PBS | 0, 14, 28 | 100 μL |
| 2, rvAc-dual | rvAc-dual | 0, 14, 28 | 108 PFU |
| 3, Mhp CV | RespiSure® ONE commercial vaccine | 0, 14, 28 | 100 μL |
| 4, PCV2 CV | Yuankexin® commercial vaccine | 0, 14, 28 | 100 μL |
| 5, rvAc-P97R1P46P42-Cap | rvAc-P97R1P46P42-Cap | 0, 14, 28 | 108 PFU |
| Group | Formulation | Immunization Time Points (DAI) | Dose |
|---|---|---|---|
| 1, rvAc-dual | rvAc-dual | 0, 14 | 109 PFU |
| 2, rvAc-P97R1P46P42-Cap | rvAc-P97R1P46P42-Cap | 0, 14 | 109 PFU |
| 3, CV | RespiSure® ONE and Yuankexin® | 0, 14 | Take 1 mL of each and mix |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Y.; Li, G.; Zheng, W.; Shu, J.; Chen, J.; Yang, F.; Wu, Y.; He, Y. Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System. Int. J. Mol. Sci. 2019, 20, 4425. https://doi.org/10.3390/ijms20184425
Tao Y, Li G, Zheng W, Shu J, Chen J, Yang F, Wu Y, He Y. Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System. International Journal of Molecular Sciences. 2019; 20(18):4425. https://doi.org/10.3390/ijms20184425
Chicago/Turabian StyleTao, Yu, Gaojian Li, Wenqian Zheng, Jianhong Shu, Jian Chen, Fang Yang, Yuehong Wu, and Yulong He. 2019. "Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System" International Journal of Molecular Sciences 20, no. 18: 4425. https://doi.org/10.3390/ijms20184425
APA StyleTao, Y., Li, G., Zheng, W., Shu, J., Chen, J., Yang, F., Wu, Y., & He, Y. (2019). Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System. International Journal of Molecular Sciences, 20(18), 4425. https://doi.org/10.3390/ijms20184425