Effects of a Sudden Drop in Salinity on Immune Response Mechanisms of Anadara kagoshimensis



Abstract
1. Introduction
2. Results
2.1. Total Count of Blood Lymphocytes and Phagocytic Activity
2.2. O2− Levels Produced from Respiratory Burst of Blood Lymphocytes and ROS Content
2.3. Lysozyme (LZM) and Acid Phosphatase (ACP) Activities
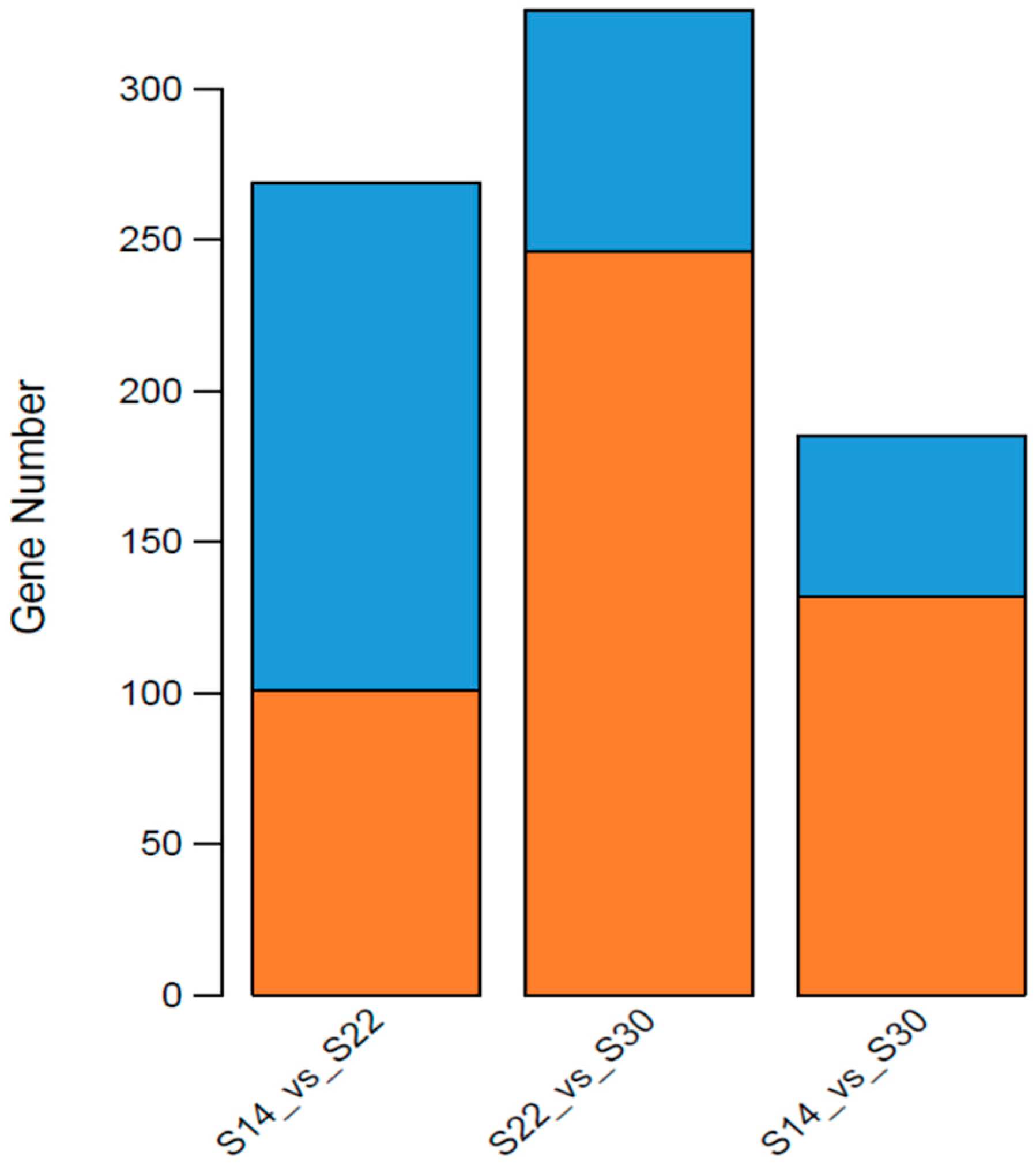
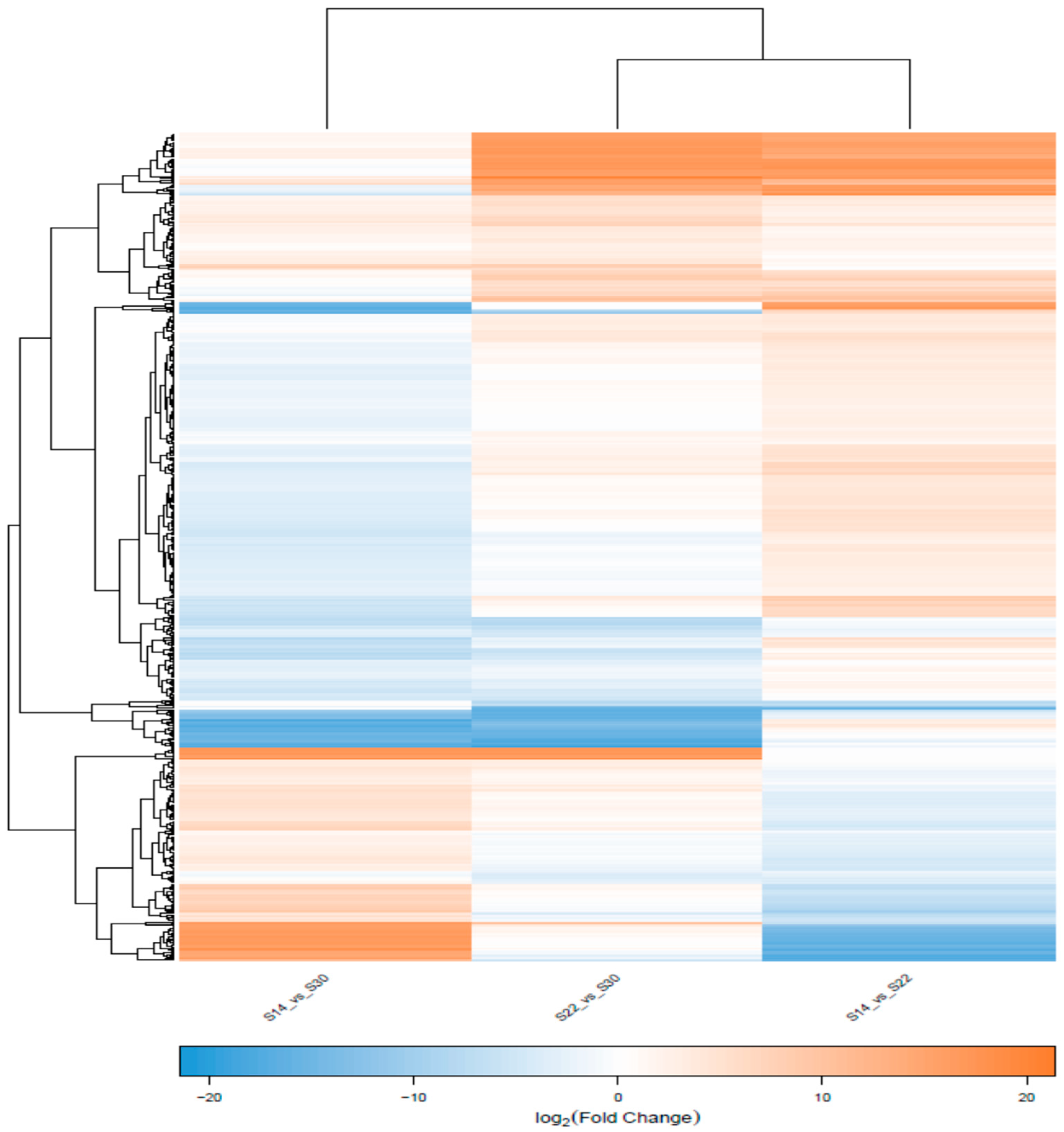
2.4. Transcriptome Sequencing and Splicing Assembly
2.5. Functional Annotation of Transcriptome Sequences
2.6. Functional Classification Based on GO and KEGG Analysis
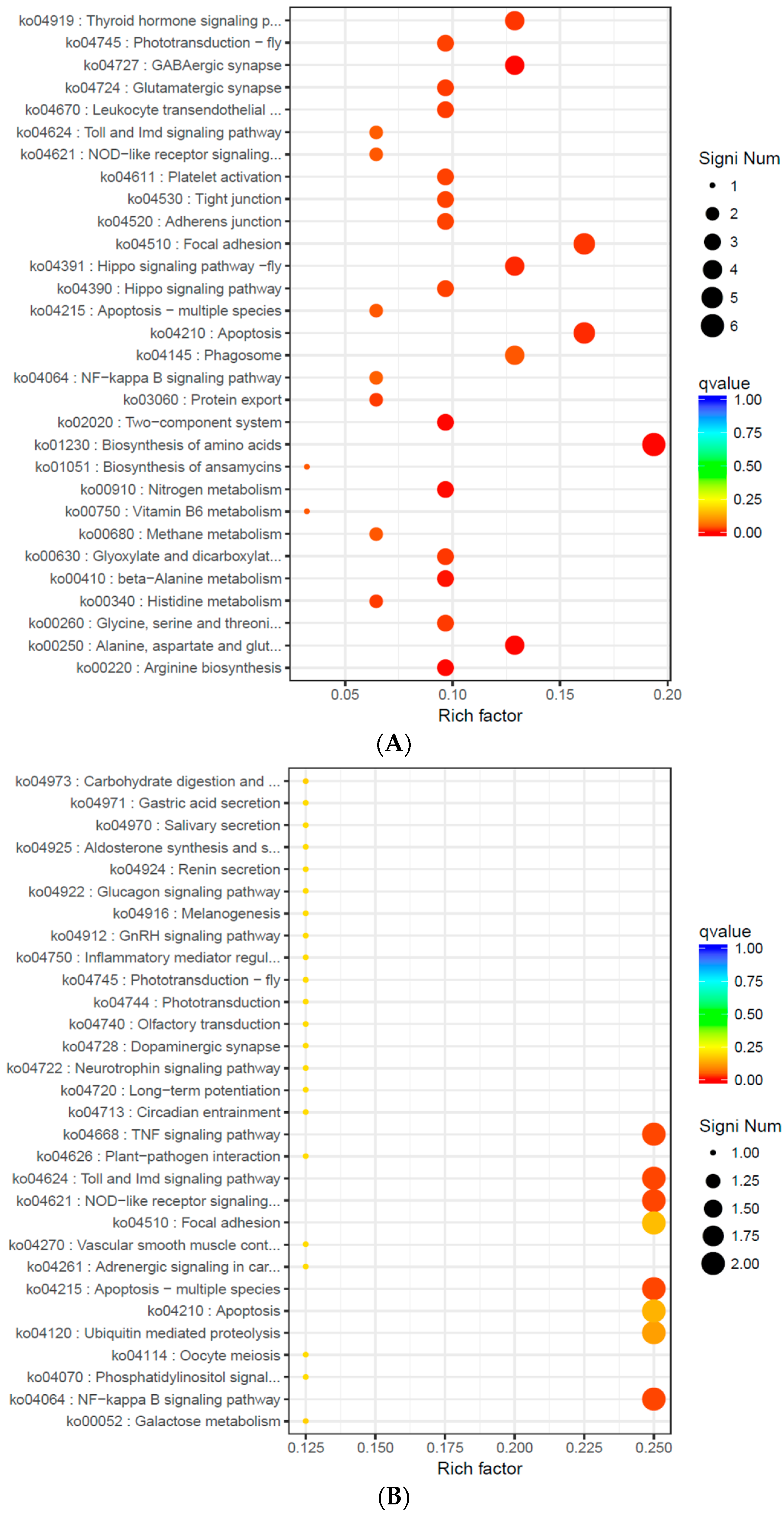
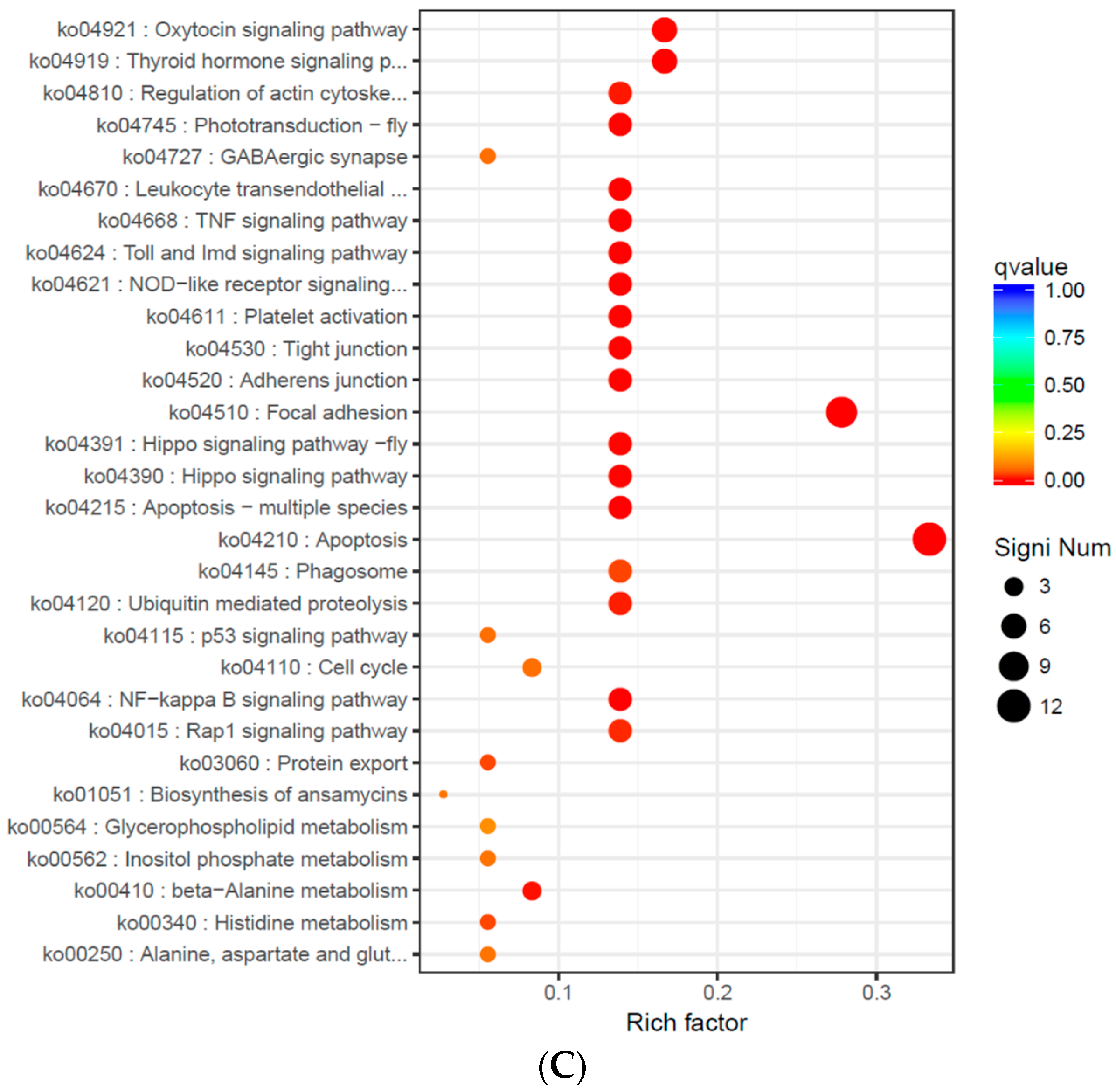
2.7. Screening of DEGs and GO and KEGG Pathway Enrichment Analysis
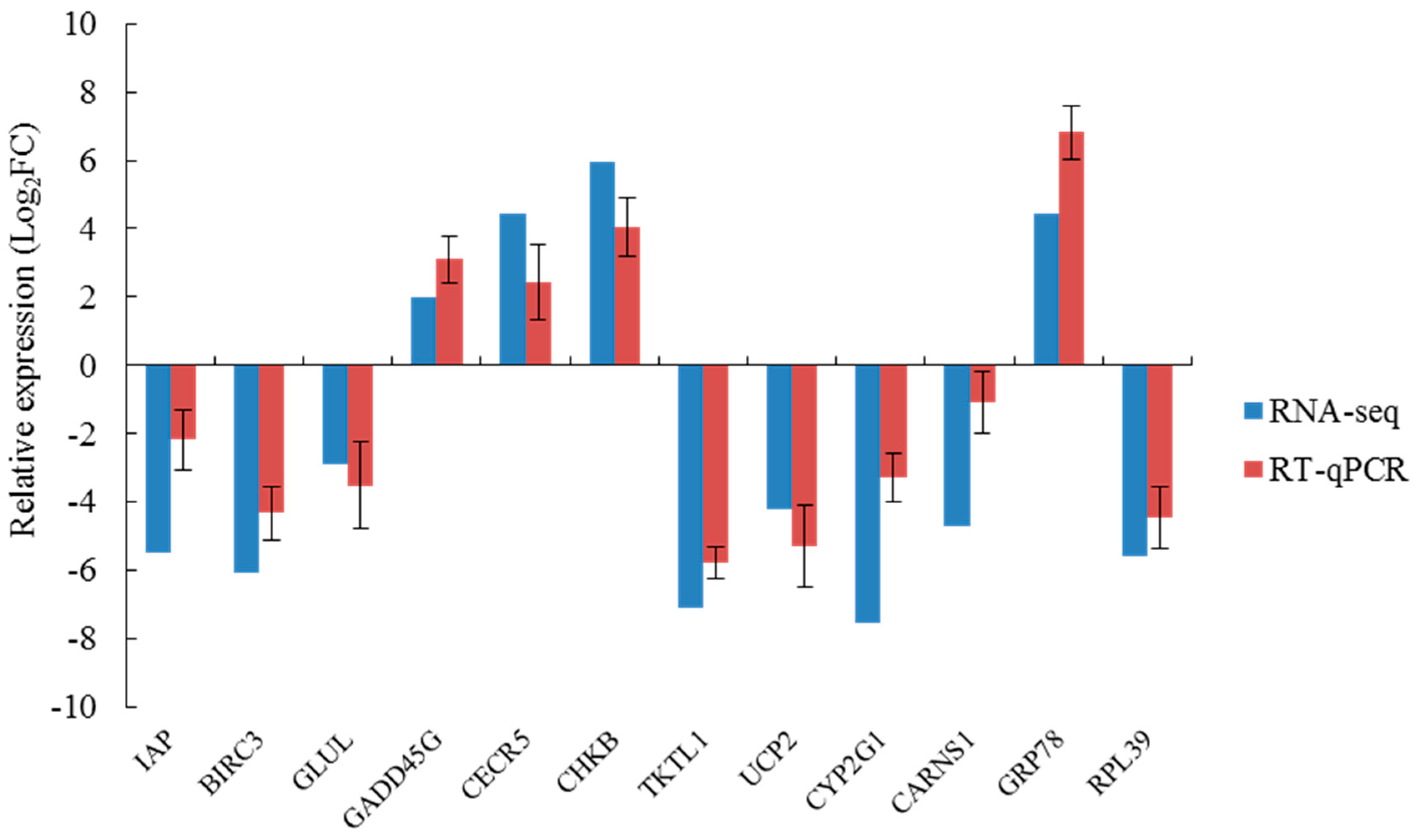
2.8. qRT-PCR Verification
3. Discussion
4. Materials and Methods
4.1. Source and Acclimation of A. kagoshimensis
4.2. Experimental Design
4.3. Assay of Total Count of Blood Lymphocytes and Phagocytic Activity
4.4. Measurement of O2− Levels Produced by the Respiratory Burst of Blood Lymphocytes
4.5. ROS and Immune-Related Enzyme Activity Assays
4.6. RNA Extraction, cDNA Library Construction, and Transcriptome Sequencing
4.6.1. Quality Control of Transcriptome Sequencing Data and Trinity Splicing
4.6.2. Functional Annotation and Analysis of Differences
4.6.3. qRT-PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Philippart, C.; Anadon, R.; Danovaro, R.; Dippner, J.; Drinkwater, K.; Hawkins, S.; Oguz, T.; O’Sullivan, G.; Reid, P.; Oǧuz, T. Impacts of climate change on European marine ecosystems: Observations, expectations and indicators. J. Exp. Mar. Biol. Ecol. 2011, 400, 52–69. [Google Scholar] [CrossRef]
- Johnson, C.R.; Banks, S.C.; Barrett, N.S.; Cazassus, F.; Dunstan, P.K.; Edgar, G.J.; Frusher, S.D.; Gardner, C.; Haddon, M.; Helidoniotis, F.; et al. Climate change cascades: Shifts in oceanography, species’ ranges and subtidal marine community dynamics in eastern Tasmania. J. Exp. Mar. Biol. Ecol. 2011, 400, 17–32. [Google Scholar] [CrossRef]
- Navarro, J.M. The effects of salinity on the physiological ecology of Choromytilus chorus (Molina, 1782) (Bivalvia: Mytilidae). J. Exp. Mar. Biol. Ecol. 1988, 122, 19–33. [Google Scholar] [CrossRef]
- Kim, W.S.; Kim, J.M.; Kim, M.S.; Park, C.W.; Huh, H.T. Effects of sudden changes in salinity on endogenous rhythms of the spotted sea bass Lateolabrax sp. Mar. Biol. 1998, 131, 219–225. [Google Scholar] [CrossRef]
- Garçon, D.P.; Masui, D.C.; Mantelatto, F.L.M.; Furriel, R.P.M.; McNamara, J.C.; Leone, F.A. Hemolymph ionic regulation and adjustments in gill (Na+, K+)-ATPase activity during salinity acclimation in the swimming crab Callinectes ornatus (Decapoda, Brachyura). Comp. Biochem. Phys. A 2009, 154, 44–55. [Google Scholar] [CrossRef]
- Huong, D.T.T.; Jasmani, S.; Jayasankar, V.; Wilder, M. Na/K-ATPase activity and osmo-ionic regulation in adult whiteleg shrimp Litopenaeus vannamei exposed to low salinities. Aquaculture 2010, 304, 88–94. [Google Scholar] [CrossRef]
- Cheng, W.; Yeh, S.-P.; Wang, C.-S.; Chen, J.-C. Osmotic and ionic changes in Taiwan abalone Haliotis diversicolor supertexta at different salinity levels. Aquaculture 2002, 203, 349–357. [Google Scholar] [CrossRef]
- Berger, V.J.; Kharazova, A.D. Mechanisms of salinity adaptations in marine molluscs. Hydrobiologia 1997, 355, 115–126. [Google Scholar] [CrossRef]
- Wang, R.C.; Wang, Z.P. Science of Marine Shellfish Culture; China Ocean University Press: Qingdao, China, 2008; pp. 370–371. [Google Scholar]
- Gunter, G. Some relations of estuarine organisms to salinity. Limnol. Oceanogr. 1961, 6, 182–190. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Macrae, T.H.; Sorgeloos, P.; Bossier, P. Stress response for disease control in aquaculture. Rev. Aquac. 2011, 3, 120–137. [Google Scholar] [CrossRef]
- Pierce, S.K. Invertebrate cell volume control mechanisms: A coordinated use of intracellular amino acids and inorganic ions as osmotic solute. Biol. Bull. 1982, 163, 405–419. [Google Scholar] [CrossRef]
- Hosoi, M.; Shinzato, C.; Takagi, M.; Hosoi-Tanabe, S.; Sawada, H.; Terasawa, E.; Toyohara, H. Taurine transporter from the giant Pacific oyster Crassostrea gigas: Function and expression in response to hyper- and hypo-osmotic stress. Fish. Sci. 2007, 73, 385–394. [Google Scholar] [CrossRef]
- Ballantyne, J.S.; Moyes, C.D. The role of divalent cations and ionic strength in the osmotic sensitivity of glutamate oxidation in oyster gill mitochondria. J. Exp. Biol. 1987, 130, 203–217. [Google Scholar]
- Greenaway, P. Calcium regulation in the freshwater mollusc, Limnaea stagnalis (L.) (Gastropoda: Pulmonata). J. Exp. Biol. 1971, 54, 199–214. [Google Scholar]
- Toyohara, H.; Ikeda, M.; Goto, C.; Sawada, H.; Hosoi, M.; Takeuchi, K.; Hayashi, I.; Imamura, S.; Yamashita, M. Osmo-responsive expression of oyster amino acid transporter gene and analysis of the regulatory region involved. Fish. Sci. 2005, 71, 465–470. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, H.; Kong, L.; Li, Q. Transcriptomic Responses to Salinity Stress in the Pacific Oyster Crassostrea gigas. PLoS ONE 2012, 7, e46244. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, X.; Huang, X.; Gu, L.; Chen, Y.; Yang, Z. Combined effects of cadmium and salinity on juvenile Takifugu obscurus: Cadmium moderates salinity tolerance; salinity decreases the toxicity of cadmium. Sci. Rep. 2016, 6, 30968. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, F.; Dong, S.; Lu, Y. De novo assembly and transcriptome analysis of osmoregulation in Litopenaeus vannamei under three cultivated conditions with different salinities. Gene 2016, 578, 185–193. [Google Scholar] [CrossRef]
- Morris, S. Neuroendocrine regulation of osmoregulation and the evolution of air-breathing in decapod crustaceans. J. Exp. Biol. 2001, 204, 979–989. [Google Scholar]
- Towle, D.W. Role of Na+ + K+-ATPase in ionic regulation by marine and estuarine animals. Mar. Biol. Lett. 1981, 2, 107–122. [Google Scholar]
- Cheng, W.; Juang, F.M.; Chen, J.C. The immune respone of Taiwan abalone Haliotis diversicolor supertexta and its susceptibility to Vibrio parahaemolyticus at different salinity levels. Fish Shellfish Immunol. 2004, 16, 295–306. [Google Scholar] [CrossRef]
- Hooper, C.; Day, R.; Slocombe, R.; Handlinger, J.; Benkendorff, K.; Day, R. Stress and immune responses in abalone: Limitations in current knowledge and investigative methods based on other models. Fish Shellfish Immunol. 2007, 22, 363–379. [Google Scholar] [CrossRef]
- Zhao, Q.; Wu, B.; Liu, Z.; Sun, X.; Zhou, L.; Yang, A.; Zhang, G. Molecular cloning, expression and biochemical characterization of hemoglobin gene from ark shell Scapharca broughtonii. Fish Shellfish Immunol. 2018, 78, 60–68. [Google Scholar] [CrossRef]
- Geng, X.; Dong, X.-H.; Tan, B.-P.; Yang, Q.-H.; Chi, S.-Y.; Liu, H.-Y.; Liu, X.-Q. Effects of dietary chitosan and Bacillus subtilis on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Fish Shellfish Immunol. 2011, 31, 400–406. [Google Scholar] [CrossRef]
- Cha, J.-H.; Rahimnejad, S.; Yang, S.-Y.; Kim, K.-W.; Lee, K.-J. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402, 50–57. [Google Scholar] [CrossRef]
- Malham, S.K.; Lacoste, A.; Gélébart, F.; Cueff, A.; Poulet, S.A. Evidence for a direct link between stress and immunity in the mollusc Haliotis tuberculate. J. Exp. Zool. Part A 2003, 295, 136–144. [Google Scholar] [CrossRef]
- Martello, L. Combined effects of pentachlorophenol and salinity stress on phagocytic and chemotactic function in two species of abalone. Aquat. Toxicol. 2000, 49, 213–225. [Google Scholar] [CrossRef]
- Ottaviani, E.; Franchini, A.; Franceschi, C. Pro-opiomelanocortin-derived peptides, cytokines, and nitric oxide in immune responses and stress: An evolutionary approach. Int. Rev. Cytol. 1997, 170, 79–141. [Google Scholar]
- Stefano, G.B.; Leung, M.K.; Zhao, X.H.; Scharrer, B. Evidence for the involvement of opioid neuropeptides in the adherence and migration of immunocompetent invertebrate hemocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 626–630. [Google Scholar] [CrossRef]
- Anderson, R.S.; Brubacher, L.L.; Calvo, L.R.; Unger, M.A.; Burreson, E.M. Effects of tributyltin and hypoxia on the progression of Perkinsus marinus infections and host defence mechanisms in oyster, Crassostrea virginica (Gmelin). J. Fish Dis. 1998, 21, 371–380. [Google Scholar] [CrossRef]
- Ordás, M.; Ordás, A.; Beloso, C.; Figueras, A. Immune parameters in carpet shell clams naturally infected with Perkinsus atlanticus. Fish Shellfish Immunol. 2000, 10, 597–609. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, D.; Wu, B.; Sun, X.; Liu, Z.; Zhao, F.; Lv, Z.; Yang, A.; Zhao, Q.; Zhang, G.; et al. Ark shell Scapharca broughtonii hemocyte response against Vibrio anguillarum challenge. Fish Shellfish Immunol. 2019, 84, 304–311. [Google Scholar] [CrossRef]
- Yang, C.; Kong, J.; Wang, Q.; Liu, Q.; Tian, Y.; Luo, K. Heterosis of haemolymph analytes of two geographic populations in Chinese shrimp Fenneropenaeus chinensis. Fish Shellfish Immunol. 2007, 23, 62–70. [Google Scholar] [CrossRef]
- Ross, N.; Firth, K.; Wang, A.; Burka, J.; Johnson, S. Changes in hydrolytic enzyme activities of naïve Atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Dis. Aquat. Org. 2000, 41, 43–51. [Google Scholar] [CrossRef]
- Qin, J.; Huang, Z.; Chen, J.; Zou, Q.; You, W.; Ke, C. Sequencing and de novo Analysis of Crassostrea angulata (Fujian Oyster) from 8 Different Developing Phases Using 454 GSFlx. PLoS ONE 2012, 7, e43653. [Google Scholar] [CrossRef]
- Ho, K.K.; Leung, P.T.; Ip, J.C.; Qiu, J.-W.; Leung, K.M. De novo transcriptomic profile in the gonadal tissues of the intertidal whelk Reishia clavigera. Mar. Pollut. Bull. 2014, 85, 499–504. [Google Scholar] [CrossRef]
- Song, H.; Yu, Z.-L.; Sun, L.-N.; Gao, Y.; Zhang, T.; Wang, H.-Y. De novo transcriptome sequencing and analysis of Rapana venosa from six different developmental stages using Hi-seq 2500. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 17, 48–57. [Google Scholar] [CrossRef]
- Hou, R.; Bao, Z.M.; Wang, S.; Su, H.L.; Li, Y.; Du, H.X.; Hu, J.J.; Wang, S.; Hu, X.L. Transcriptome sequencing and de novo analysis for Yesso scallop (Patinopecten yessoensis) using 454 GS FLX. PLoS ONE 2011, 6, e21560. [Google Scholar] [CrossRef]
- Huan, P.; Wang, H.X.; Liu, B.Z. Transcriptomic analysis of the clam Meretrix meretrix on different larval stages. Mar. Biotechnol. 2012, 14, 69–78. [Google Scholar] [CrossRef]
- Fokina, N.N.; Nefedova, Z.A.; Nemova, N.N.; Khalaman, V.V.; Fokina, N. Modulating role of lipids and their fatty acids in adaptation of the White Sea mussels Mytilus edulis L. to environmental salinity change. J. Evol. Biochem. Physiol. 2007, 43, 379–387. [Google Scholar] [CrossRef]
- Burton, R.S. Regulation of proline synthesis in osmotic response: Effects of protein synthesis inhibitors. J. Exp. Zool. Part A 1991, 259, 272–277. [Google Scholar] [CrossRef]
- Gilles, R. Intracellular organic osmotic effectors. In Gilles(ed) Mechanisms of Osmoregulation in Animals: Maintenance of Cell Volume; Wiley: New York, NY, USA, 1979; pp. 111–154. [Google Scholar]
- McNamara, J.C.; Rosa, J.C.; Greene, L.J.; Augusto, A. Free amino acid pools as effectors of osmostic adjustment in different tissues of the freshwater shrimp Macrobrachium olfersii (Crustacea, Decapoda) during long-term salinity acclimation. Mar. Freshw. Behav. Phy. 2004, 37, 193–208. [Google Scholar] [CrossRef]
- Augusto, A.; Greene, L.J.; Laure, H.J.; McNamara, J.C. Adaptive shifts in osmoregulatory strategy and the invasion of freshwater by brachyuran crabs: Evidence from Dilocarcinus pagei (Trichodactylidae). J. Exp. Zool. Part A 2007, 307, 688–698. [Google Scholar] [CrossRef]
- Pascual-Anaya, J.; D’Aniello, S. Free amino acids in the nervous system of the amphioxus Branchiostoma lanceolatum. A comparative study. Int. J. Biol. Sci. 2006, 2, 87–92. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, L.; Zhou, Z.; Liu, Y.; Dong, M.; Wang, W.; Song, X.; Wang, M.; Gao, Q.; Song, L. Transcriptomic analysis of oyster Crassostrea gigas larvae illustrates the response patterns regulated by catecholaminergic system upon acute heat and bacterial stress. Dev. Comp. Immunol. 2017, 73, 52–60. [Google Scholar] [CrossRef]
- Stefano, G.B.; Smith, E.M.; Cadet, P.; Hughes, T.K. HIV gp120 alteration of DAMA and IL-1 alpha induced chemotaxic responses in human and invertebrate immunocytes. J. Neuroimmunol. 1993, 43, 177–184. [Google Scholar] [CrossRef]
- Ottaviani, E. The mollusc as a suitable model for mammalian immune-neuroendocrine investigations. Invert. Surviv. J. 2004, 1, 2–4. [Google Scholar]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef]
- Deveraux, Q.L.; Reed, J.C. IAP family proteins-suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef]
- Goedken, M.; Morsey, B.; Sunila, I.; Dungan, C.; De Guise, S. The effects of temperature and salinity on apoptosis of Crassostrea virginica hemocytes and Perkinsus marinus. J. Shellfish Res. 2005, 24, 177–183. [Google Scholar]
- Cherkasov, A.A.; Overton, R.A.; Sokolov, E.P.; Sokolova, I.M. Temperature-dependent effects of cadmium and purine nucleotides on mitochondrial aconitase from a marine ectotherm, Crassostrea virginica: A role of temperature in oxidative stress and allosteric enzyme regulation. J. Exp. Biol. 2007, 210, 46–55. [Google Scholar] [CrossRef]
- Lannig, G.; Flores, J.F.; Sokolova, I.M. Temperature-dependent stress response in oysters, Crassostrea virginica: Pollution reduces temperature tolerance in oysters. Aquat. Toxicol. 2006, 79, 278–287. [Google Scholar] [CrossRef]
- Baršienė, J.; Andreikėnaitė, L.; Garnaga, G.; Rybakovas, A. Genotoxic and cytotoxic effects in bivalve mollusks Macoma balthica and Mytilus edulis from the Baltic Sea. Ekologija 2008, 54, 44–50. [Google Scholar] [CrossRef]
- Russo, J.; Madec, L.; Brehelin, M. Effect of a toxicant on phagocytosis pathways in the freshwater snail Lymnaea stagnalis. Cell Tissue Res. 2008, 333, 1147–1158. [Google Scholar] [CrossRef]
- Abele, D.; Heise, K.; O Pörtner, H.; Puntarulo, S. Temperature-dependence of mitochondrial function and production of reactive oxygen species in the intertidal mud clam Mya arenaria. J. Exp. Biol. 2002, 205, 1831–1841. [Google Scholar]
- Zhang, Q.; Ma, H.M.; Mai, K.S.; Zhang, W.B.; Liufu, Z.G.; Xu, W. Interaction of dietary Bacillus subtilis and fructooligosaccharide on the growth performance, nonspecific immunity of sea cucumber, Apostichopus japonicas. Fish Shellfish Immunol. 2010, 29, 204–211. [Google Scholar] [CrossRef]
- Secombes, C.J. Isolation of salmonid macrophages and analysis of their killing activity. Tech. Fish Immunol. 1990, 1, 137–154. [Google Scholar]
- Gao, X.L.; Li, X.; Li, M.J.; Song, C.B.; Liu, Y. Effects of light intensity on metabolism and antioxidant defense in Haliotis discus hannai Ino. Aquaculture 2017, 465, 78–87. [Google Scholar]
- Ellis, A.E. Lysozyme assay. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Robertsen, B.S., Van Muiswinkel, W.B., Eds.; SOS Publication: Fair Haven, NJ, USA, 1990; pp. 101–103. [Google Scholar]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Gen. Biol. 2010, 11, R106. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Salinity Treatments | ANOVA p | ||
|---|---|---|---|---|
| 30‰ | 22‰ | 14‰ | ||
| Total hemocyte counts (×107 cells·mL−1) | 2.43 ± 0.19 b | 2.97 ± 0.31 a | 2.15 ± 0.24 b | 0.002 |
| Phagocytic activity (%) | 15.26 ± 2.26 c | 22.46 ± 4.17 b | 30.95 ± 3.55 a | <0.001 |
| Respiratory burst activity (OD630/107 cells·mL−1) | 0.36 ± 0.03 b | 0.41 ± 0.07 b | 0.53 ± 0.04 a | 0.001 |
| Reactive Oxygen Species (A.U.) | 48.21 ± 8.77 b | 106.38 ± 15.61 b | 462.19 ± 39.24 a | <0.001 |
| Items | All Number | >=500 bp Number | >=1000 bp Number | N50 Length of Unigenes (bp) | N90 Length of Unigenes (bp) | Max Length (bp) | Min Length (bp) | Total Length (bp) | Average Length (bp) |
|---|---|---|---|---|---|---|---|---|---|
| Transcript | 480,426 | 206,812 | 94,603 | 1070 | 293 | 16,628 | 201 | 344,014,385 | 716.06 |
| Unigene | 194,861 | 71,917 | 30,728 | 933 | 265 | 16,628 | 201 | 125,490,202 | 644 |
| Database | Number of Genes | Percentage (%) |
|---|---|---|
| Annotated in CDD | 22,654 | 11.63 |
| Annotated in KOG | 19,868 | 10.2 |
| Annotated in NR | 36,240 | 18.6 |
| Annotated in NT | 15,928 | 8.17 |
| Annotated in PFAM | 16,875 | 8.66 |
| Annotated in Swissprot | 30,091 | 15.44 |
| Annotated in TrEMBL | 35,539 | 18.24 |
| Annotated in GO | 33,522 | 17.2 |
| Annotated in KEGG | 12,644 | 6.49 |
| Annotated in at least one database | 45,883 | 23.55 |
| Annotated in all database | 3000 | 1.54 |
| Total genes | 194,861 | 100 |
| Comparison | Number of Differentially Expressed Genes | ||
|---|---|---|---|
| Up-Regulated | Down-Regulated | Total Number | |
| S14 vs. S22 | 101 | 168 | 269 |
| S22 vs. S30 | 246 | 80 | 326 |
| S14 vs. S30 | 132 | 53 | 185 |
| Go No. | Go Term | Significant (n/m) | Annotated (N/M) | p-Value | Corrected p-Value |
|---|---|---|---|---|---|
| S14 vs. S22 | |||||
| GO:1901605 | alpha-amino acid metabolic process | 9/54 | 499/26,336 | 0.00000073 | 0.006428865 |
| GO:0006541 | glutamine metabolic process | 5/54 | 85/26,336 | 0.00000087 | 0.006428865 |
| GO:0006520 | cellular amino acid metabolic process | 10/54 | 728/26,336 | 0.000002 | 0.00960635 |
| GO:1901607 | alpha-amino acid biosynthetic process | 6/54 | 191/26,336 | 0.0000026 | 0.00960635 |
| GO:0006542 | glutamine biosynthetic process | 3/54 | 15/26,336 | 0.0000036 | 0.01064088 |
| GO:0008652 | cellular amino acid biosynthetic process | 6/54 | 219/26,336 | 0.0000057 | 0.01404005 |
| GO:0043436 | oxoacid metabolic process | 15/54 | 1991/26,336 | 0.0000073 | 0.015412386 |
| GO:0006082 | organic acid metabolic process | 15/54 | 2015/26,336 | 0.0000085 | 0.015702688 |
| GO:0035499 | carnosine biosynthetic process | 2/54 | 3/26,336 | 0.000012 | 0.019705333 |
| GO:0019752 | carboxylic acid metabolic process | 14/54 | 1881/26,336 | 0.000018 | 0.0266022 |
| GO:0016053 | organic acid biosynthetic process | 8/54 | 586/26,336 | 0.000024 | 0.028421154 |
| GO:0046394 | carboxylic acid biosynthetic process | 8/54 | 586/26,336 | 0.000024 | 0.028421154 |
| GO:0009064 | glutamine family amino acid metabolic process | 5/54 | 169/26,336 | 0.000025 | 0.028421154 |
| GO:0006548 | histidine catabolic process | 2/54 | 5/26,336 | 0.000041 | 0.033663278 |
| GO:0010510 | regulation of acetyl-CoA biosynthetic process from pyruvate | 2/54 | 5/26,336 | 0.000041 | 0.033663278 |
| GO:0035498 | carnosine metabolic process | 2/54 | 5/26,336 | 0.000041 | 0.033663278 |
| GO:0050812 | regulation of acyl-CoA biosynthetic process | 2/54 | 5/26,336 | 0.000041 | 0.033663278 |
| GO:0052805 | imidazole-containing compound catabolic process | 2/54 | 5/26,336 | 0.000041 | 0.033663278 |
| S22 vs. S30 | |||||
| GO:0070613 | regulation of protein processing | 7/66 | 118/26,336 | 0.000000019 | 0.000136706 |
| GO:1903317 | regulation of protein maturation | 7/66 | 118/26,336 | 0.000000019 | 0.000136706 |
| GO:0010955 | negative regulation of protein processing | 6/66 | 76/26,336 | 0.000000037 | 0.000136706 |
| GO:1903318 | negative regulation of protein maturation | 6/66 | 76/26,336 | 0.000000037 | 0.000136706 |
| GO:0097340 | inhibition of cysteine-type endopeptidase activity | 5/66 | 55/26,336 | 0.00000027 | 0.000570047 |
| GO:0097341 | zymogen inhibition | 5/66 | 55/26,336 | 0.00000027 | 0.000570047 |
| GO:1990001 | inhibition of cysteine-type endopeptidase activity involved in apoptotic process | 5/66 | 55/26,336 | 0.00000027 | 0.000570047 |
| GO:0043154 | negative regulation of cysteine-type endopeptidase activity involved in apoptotic process | 5/66 | 125/26,336 | 0.000016 | 0.025527364 |
| GO:0016485 | protein processing | 7/66 | 322/26,336 | 0.000016 | 0.025527364 |
| GO:2000117 | negative regulation of cysteine-type endopeptidase activity | 5/66 | 128/26,336 | 0.000018 | 0.025527364 |
| GO:0035499 | carnosine biosynthetic process | 2/66 | 3/26,336 | 0.000019 | 0.025527364 |
| GO:0051604 | protein maturation | 7/66 | 349/26,336 | 0.000027 | 0.03325275 |
| GO:0045861 | negative regulation of proteolysis | 6/66 | 245/26,336 | 0.000035 | 0.039789615 |
| GO:0010951 | negative regulation of endopeptidase activity | 5/66 | 154/26,336 | 0.000043 | 0.043633238 |
| GO:0010466 | negative regulation of peptidase activity | 5/66 | 157/26,336 | 0.000047 | 0.043633238 |
| GO:0043569 | negative regulation of insulin-like growth factor receptor signaling pathway | 3/66 | 29/26,336 | 0.000052 | 0.043633238 |
| GO:0006548 | histidine catabolic process | 2/66 | 5/26,336 | 0.000062 | 0.043633238 |
| GO:0010510 | regulation of acetyl-CoA biosynthetic process from pyruvate | 2/66 | 5/26,336 | 0.000062 | 0.043633238 |
| GO:0035498 | carnosine metabolic process | 2/66 | 5/26,336 | 0.000062 | 0.043633238 |
| GO:0050812 | regulation of acyl-CoA biosynthetic process | 2/66 | 5/26,336 | 0.000062 | 0.043633238 |
| GO:0052805 | imidazole-containing compound catabolic process | 2/66 | 5/26,336 | 0.000062 | 0.043633238 |
| Unigene-ID | Gene Name | Sequence (5’-3’) | Efficiency | Product Size (bp) |
|---|---|---|---|---|
| TRINITY_DN64699_c0_g1 | IAP | F: GCAAACTTCTTGCCGTGCGGCA | 97.80% | 168 |
| R: CTGCATGCGACACCCAATCAG | ||||
| TRINITY_DN65676_c2_g1 | BIRC3 | F: ACGGACCAGGCAGTTGTCGT | 102.43% | 192 |
| R: CGCCGGTTGTGTGGCCACCCTGT | ||||
| TRINITY_DN65035_c3_g1 | GLUL | F: GAATCTTCTCGCAAATCCGCG | 98.15% | 177 |
| R: ATTGATTGTTGTGCTTCCC | ||||
| TRINITY_DN53170_c0_g1 | GADD45G | F: ACCGCCGAACCAGTATTTGGC | 107.81% | 204 |
| R: CAGGAAACACGGAGCCGATG | ||||
| TRINITY_DN63352_c4_g2 | CECR5 | F: AGTTTGACCTCCTGCTGACC | 101.92% | 225 |
| R: TGACATCTTGAAATCCGGGCA | ||||
| TRINITY_DN57342_c5_g1 | CHKB | F: CGCAACCTGCCTAAGTCTCT | 98.75% | 159 |
| R: GGTCCGCTACGTTGTGACCGC | ||||
| TRINITY_DN41276_c0_g1 | TKTL1 | F: CTTCTAACTGCTCCTTCGGAATTC | 105.38% | 207 |
| R: GAACACACCACGGCACTAAGCC | ||||
| TRINITY_DN63081_c2_g1 | UCP2 | F: CAGATAACATCAGTTCTGGACG | 96.75% | 190 |
| R: ACCGGTCAATGTTCCAGTCCTT | ||||
| TRINITY_DN72946_c3_g5 | CYP2G1 | F: ATCCTCTTGGACATCACCGA | 100.37% | 146 |
| R: GGAACAGGACCAGGCAGAAG | ||||
| TRINITY_DN59014_c0_g1 | CARNS1 | F: GACCGGTGATGAAGAATTG | 99.26% | 188 |
| R: GTTAAAGCCTAGCAGATT | ||||
| TRINITY_DN66154_c0_g2 | GRP78 | F: ACACCTCCGACGAAATATTCC | 101.79% | 201 |
| R: CATCAACCGGGACACGATCGCC | ||||
| TRINITY_DN64918_c2_g2 | RPL39 | F: CACTTCAATCGCCTGGTTCAAT | 98.59% | 173 |
| R: GACCCCTGGTATTTGTGCCAG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Li, L.; Liu, Y.; Gao, X. Effects of a Sudden Drop in Salinity on Immune Response Mechanisms of Anadara kagoshimensis. Int. J. Mol. Sci. 2019, 20, 4365. https://doi.org/10.3390/ijms20184365
Zhang M, Li L, Liu Y, Gao X. Effects of a Sudden Drop in Salinity on Immune Response Mechanisms of Anadara kagoshimensis. International Journal of Molecular Sciences. 2019; 20(18):4365. https://doi.org/10.3390/ijms20184365
Chicago/Turabian StyleZhang, Mo, Li Li, Ying Liu, and Xiaolong Gao. 2019. "Effects of a Sudden Drop in Salinity on Immune Response Mechanisms of Anadara kagoshimensis" International Journal of Molecular Sciences 20, no. 18: 4365. https://doi.org/10.3390/ijms20184365
APA StyleZhang, M., Li, L., Liu, Y., & Gao, X. (2019). Effects of a Sudden Drop in Salinity on Immune Response Mechanisms of Anadara kagoshimensis. International Journal of Molecular Sciences, 20(18), 4365. https://doi.org/10.3390/ijms20184365