Exploring the Relationship between Crassulacean Acid Metabolism (CAM) and Mineral Nutrition with a Special Focus on Nitrogen


{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Induction and Regulation of CAM Photosynthesis
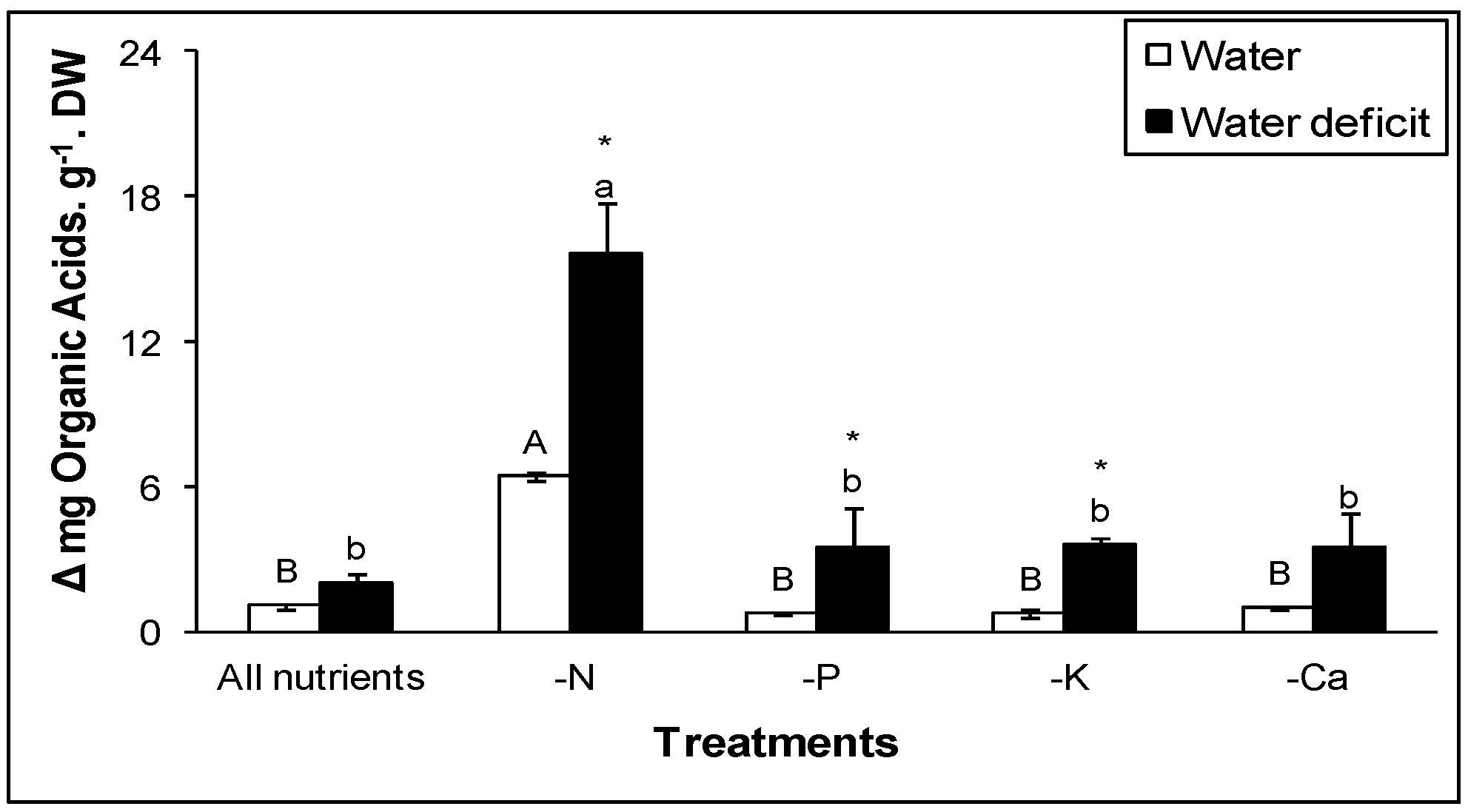
1.2. Nutrient Availability Interactions with CAM
2. Relationship between Nitrogen and CAM
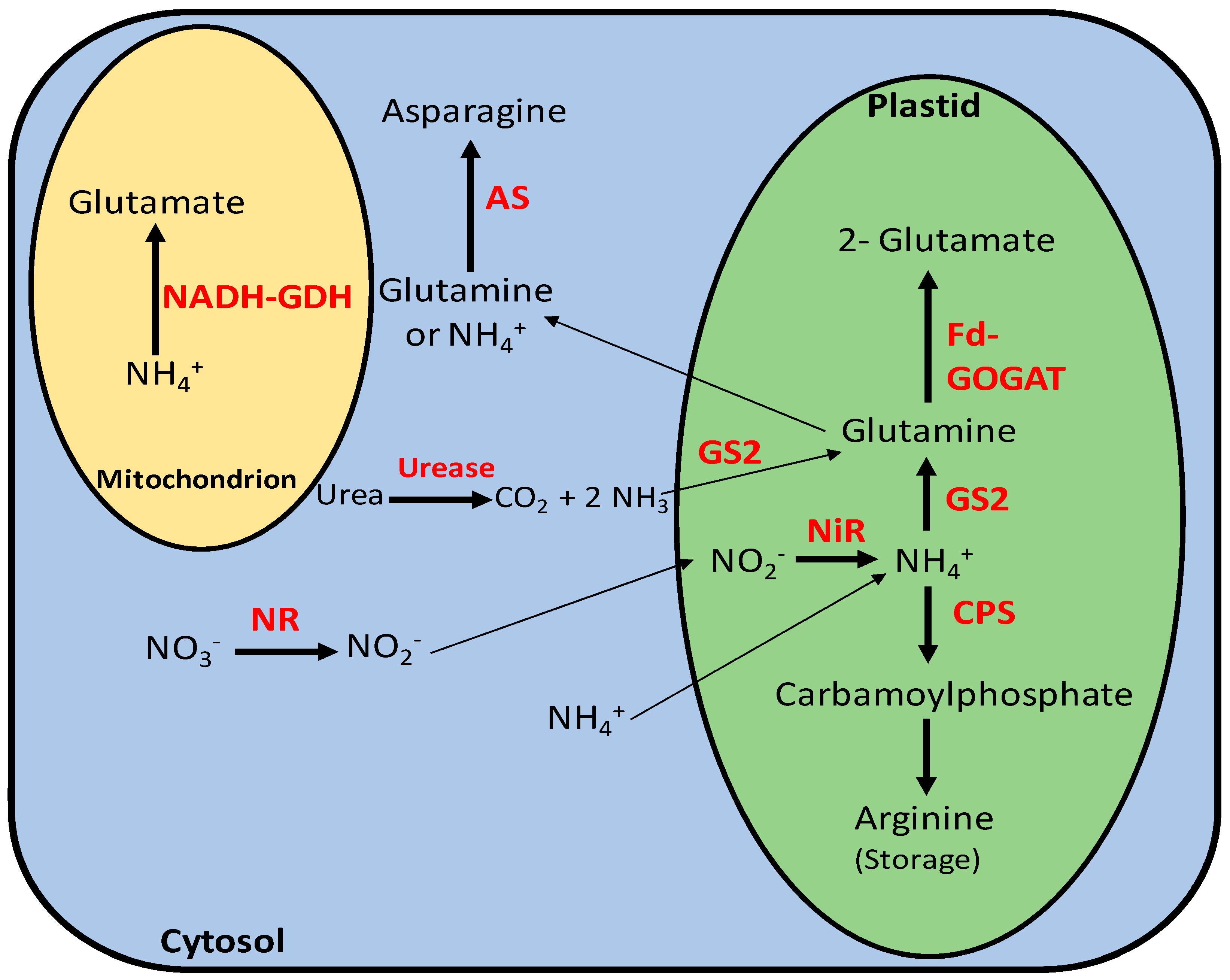
2.1. Nitrate, Ammonium, and Urea Uptake and Transport into the Cells
2.2. Nitrate, Ammonium, and Urea Assimilation
2.3. Nitrate Effects on CAM
2.4. Ammonium Effects on CAM
2.5. Organic N Source Effects on CAM
2.6. Nitrogen Deposition
3. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Griffiths, H. Crassulacean acid metabolism: A re-appraisal of physiological plasticity in form and function. Adv. Bot. Res. 1989, 15, 43–92. [Google Scholar]
- Winter, K. Crassulacean acid metabolism. In Photosynthetic Mechanisms and the Environment; Barber, J., Baker, N.R., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 329–387. [Google Scholar]
- Christopher, J.T.; Holtum, J.A.M. Patterns of carbohydrate partitioning in the leaves of Crassulacean acid metabolism species during deacidification. Plant Physiol. 1996, 112, 393–399. [Google Scholar] [CrossRef]
- Lüttge, U. CO2-concentrating: Consequences in crassulacean acid metabolism. J. Exp. Bot. 2002, 53, 2131–2142. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Monson, R.K. Evolutionary and ecological aspects of photosynthetic pathway variation. Annu. Rev. Ecol. Syst. 1993, 24, 411–439. [Google Scholar] [CrossRef]
- Ting, I.P. Crassulacean acid metabolism. Ann. Rev. Plant Physiol. 1985, 36, 595–622. [Google Scholar] [CrossRef]
- Cushman, J.C.; Bohnert, H.J. Molecular Genetics of crassulacean acid metabolism. Plant Physiol. 1997, 113, 667–676. [Google Scholar] [CrossRef]
- Lüttge, U. Ecophysiology of crassulacean acid metabolism (CAM). Ann. Bot. 2004, 93, 629–652. [Google Scholar] [CrossRef]
- Kornas, A.; Fisher-Schliebs, E.; Lüttge, U.; Miszalski, Z. Adaptation of the obligate CAM plant Clusia alata to light stress: Metabolic responses. J. Plant Physiol. 2009, 166, 1914–1922. [Google Scholar] [CrossRef]
- Wai, C.M.; VanBuren, R.; Zhang, J.; Huang, L.; Miao, W.; Edger, P.P.; Yim, W.C.; Priest, H.D.; Meyers, B.C.; Mockler, T.; et al. Temporal and spatial transcriptomic and microRNA dynamics of CAM photosynthesis in pineapple. Plant J. 2017, 92, 19–30. [Google Scholar] [CrossRef]
- Maxwell, C.; Griffiths, H.; Youngs, A.J. Photosynthetic acclimation to light regime and water stress by the C3-CAM epiphyte Guzmania monostachia: Gas-exchange characteristics, photochemical efficiency and xanthophyll cycle. Funct. Ecol. 1994, 8, 746–754. [Google Scholar] [CrossRef]
- Drennan, P.M.; Nobel, P.S. Responses of CAM species to increasing atmospheric CO2 concentrations. Plant Cell Environ. 2000, 23, 767–781. [Google Scholar] [CrossRef]
- Winter, K.; Holtum, J.A.M. Induction and reversal of crassulacean acid metabolism in Calandrinia polyandra: Effects of soil moisture and nutrients. Funct. Plant Biol. 2011, 38, 576–582. [Google Scholar] [CrossRef]
- Pereira, P.N.; Purgatto, E.; Mercier, H. Spatial division of phosphoenolpyruvate carboxylase and nitrate reductase activity and its regulation by cytokinins in CAM-induced leaves of Guzmania monostachia (Bromeliaceae). J. Plant Physiol. 2013, 170, 1067–1074. [Google Scholar] [CrossRef]
- Winter, K.; Holtum, J.A.M. Facultative crassulacean acid metabolism (CAM) plants: Powerful tools for unravelling the functional elements of CAM photosynthesis. J. Exp. Bot. 2014, 65, 3425–3441. [Google Scholar] [CrossRef]
- Harris, F.S.; Martin, C.E. Correlation between CAM-cycling and photosynthetic gas exchange in five species of Talinum (Portulacaceae). Plant Physiol. 1991, 96, 1118–1124. [Google Scholar] [CrossRef][Green Version]
- Winter, K.; Holtum, J.A.M. How closely do the δ13C values of Crassulacean acid metabolism plants reflect the proportion of CO2 fixed during day and night? Plant Physiol. 2002, 129, 1843–1851. [Google Scholar] [CrossRef]
- Montero, E.; Francisco, A.M.; Montes, E.; Herrera, A. Salinity induction of recycling crassulacean acid metabolism and salt tolerance in plants of Talinum triangulare. Ann. Bot. 2018, 121, 1333–1342. [Google Scholar] [CrossRef]
- Rayder, L.; Ting, I.P. CAM-idling in Hoya carnosa (Asclepiadaceae). Photosynth. Res. 1983, 4, 203–211. [Google Scholar] [CrossRef]
- Smith, J.A.C.; Winter, K. An introduction to crassulacean acid metabolism. Biochemical principles and ecological diversity. In Crassulacean Acid Metabolism; Springer-Verlag: Berlin, Germany, 1996; Volume 114, pp. 1–13. [Google Scholar]
- Cushman, J.C.; Borland, A.M. Induction of crassulacean acid metabolism by water limitation. Plant Cell Environ. 2002, 25, 295–310. [Google Scholar] [CrossRef]
- Ting, P.I. Effects of abscisic acid on CAM in Portulacaria afra. Photosynth Res. 1981, 2, 39–48. [Google Scholar] [CrossRef]
- Vernon, D.M.; Ostrem, J.A.; Bohnert, H.J. Stress perception and response in a facultative halophyte: The regulation of salinity-induced genes in Mesembryanthemum crystallinum. Plant Cell Environ. 1993, 16, 437–444. [Google Scholar] [CrossRef]
- Tayby, T.; Sotta, B.; Gehrig, H.; Güçlü, S.; Kluge, M.; Brulfert, J. Differential effects of abscisic acid on phosphoenolpyruvate carboxylase and CAM operation in Kalanchoë blossfeldiana. Bot. Acta 1995, 108, 240–246. [Google Scholar] [CrossRef]
- Taybi, T.; Cushman, J.C. Signaling events leading to crassulacean acid metabolism induction in the common ice plant. Plant Physiol. 1999, 121, 545–555. [Google Scholar] [CrossRef]
- Taybi, T.; Cushman, J.C. Abscisic acid signaling and protein synthesis requirements for phosphoenolpyruvate carboxylase transcript induction in the common ice plant. J. Plant Physiol. 2002, 159, 1235–1243. [Google Scholar] [CrossRef]
- Thomas, J.C.; Bohnert, H.J. Salt stress perception and plant growth regulators in the halophyte Mesembryanthemum crystallinum. Plant Physiol. 1993, 103, 1299–1304. [Google Scholar] [CrossRef][Green Version]
- Ceusters, J.; Borland, A.M.; De Proft, M.P. Drought adaptation in plants with crassulacean acid metabolism involves the flexible use of different storage carbohydrate pools. Plant Signal Behav. 2009, 4, 212–214. [Google Scholar] [CrossRef][Green Version]
- Young, D.R.; Nobel, P.S. Predictions of soil-water potentials in the north-western Sonoran Desert. J. Ecol. 1986, 74, 143–154. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernández, R.J. Modifying the ‘pulse-reserve’ paradigm for desert of North America: Precipitation pulses, soil water, and plant responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef]
- Nobel, P.S.; Quero, E.; Linares, H. Differential growth responses of agaves to nitrogen, phosphorus, potassium, and boron applications. J. Plant Nutr. 1988, 11, 1683–1700. [Google Scholar] [CrossRef]
- Nobel, P.S. A nutrient index quantifying productivity of Agaves and Cacti. J. Appl. Ecol. 1989, 26, 635–645. [Google Scholar] [CrossRef]
- Chen, Y.F.; Wang, Y.; Wu, W.H. Membrane Transporters for Nitrogen, Phosphate and Potassium Uptake in Plants. J. Integr. Plant Biol. 2008, 50, 835–848. [Google Scholar] [CrossRef]
- Peoples, T.R.; Koch, D.W. Role of potassium in carbon dioxide assimilation in Medicago sativa L. Plant Physiol. 1979, 63, 878–881. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef]
- Rychter, A.M.; Rao, I.M. Role of phosphorus in photosynthetic carbon metabolism. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; Taylor and Francis Group: London, UK, 2005; pp. 123–148. [Google Scholar]
- Maiquetía, M.; Cáceres, A.; Herrera, A. Mycorrhization and phosphorus nutrition affect water relations and CAM induction by drought in seedlings of Clusia minor. Ann. Bot. 2009, 103, 525–532. [Google Scholar] [CrossRef]
- Schulze, E.D.; Beck, E.; Müller-Hohenstein, K. Pflanzenökologie. In Plant Ecology; Springer: Heidelberg, Germany, 2002. [Google Scholar]
- Paul, M.J.; Cockburn, W. The simulation of CAM activity in Mesembryanthemum crystallinum in nitrate-and phosphate-deficient conditions. New Phytol. 1990, 114, 391–398. [Google Scholar] [CrossRef]
- Paul, M.J.; Cockburn, W. Pinitol, a compatible solute in Mesembryanthemum crystallinum. J. Exp. Bot. 1989, 40, 1093–1098. [Google Scholar] [CrossRef]
- Rodrigues, M.A.; Freschi, L.; Pereira, P.N.; Mercier, H. Interactions between nutrients and crassulacean acid metabolism. Prog Bot. 2014, 75, 167–186. [Google Scholar]
- Santos, I.; Salema, R. Nitrogen nutrition and the level of crassulacean acid metabolism in Kalanchoë lateritia Engl. Plant Cell Environ. 1991, 14, 311–317. [Google Scholar] [CrossRef]
- Winter, K.; Foster, J.G.; Schmitt, M.R.; Edwards, G.E. Activity and quantity of ribulose bisphosphate carboxylase- and phosphoenolpyruvate carboxylase protein in two Crassulacean acid metabolism plants in relation to leaf age, nitrogen nutrition, and point in time during a day/night cycle. Planta 1982, 154, 309–317. [Google Scholar] [CrossRef]
- Nobel, P.S. Nutrient levels in cacti relation to nocturnal acid accumulation and growth. Amer. J. Bot. 1983, 70, 1244–1253. [Google Scholar] [CrossRef]
- Ota, K. CAM photosynthesis under drought conditions in Kalanchoë blossfeldiana grown with nitrate or ammonium as the sole nitrogen source. Plant Cell Physiol. 1988, 29, 801–806. [Google Scholar]
- Yu, K.; D’Odorico, P.; Carr, D.E.; Personium, A.; Collins, S.L. The effect of nitrogen availability and water conditions on competition between a facultative CAM plant and an invasive grass. Ecol. Evol. 2017, 7, 7739–7749. [Google Scholar] [CrossRef]
- Yu, K.L.; D’Odorico, P.; Li, W.; He, Y.L. Effects of competition on induction of crassulacean acid metabolism in a facultative CAM plant. Oecologia 2017, 184, 351–361. [Google Scholar] [CrossRef]
- Tucker, M.R. Essential Plant Nutrients: Their Presence in North Carolina Soils and Role in Plant Nutrition; Dept. of Agriculture and Consumer Services, Agronomic Division: Raleigh, NC, USA, 1999.
- Mokhele, B.; Zhan, X.; Yang, G.; Zhang, X. Review: Nitrogen assimilation in crop plants and its affecting factors. J. Plant Sci. 2012, 92, 399–405. [Google Scholar] [CrossRef]
- Tobin, A.K.; Yamaya, T. Cellular compartmentation of ammonium assimilation in rice and barley. J. Exp. Bot. 2001, 52, 591–604. [Google Scholar] [CrossRef]
- Kojima, S.; Bohner, A.; von Wirén, N. Molecular mechanisms of urea transport in plants. J. Membr. Biol. 2006, 212, 83–91. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.H.; Wang, P.F.; Hou, J.; Li, W.; Zhang, W.J. Metabolic adaptations to ammonium-induced oxidative stress in leaves of the submerged macrophyte Vallisneria natans (Lour.) Hara. Aquat. Toxicol. 2008, 87, 88–98. [Google Scholar] [CrossRef]
- Witte, C.P. Urea metabolism in plants. Plant Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular mechanisms of ammonium transport and accumulation in plants. FEBS Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signaling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef]
- Liu, L.H.; Ludewig, U.; Frommer, W.B.; von Wirén, N. AtDUR3 encodes a new type of high-affinity urea/H+ symporter in Arabidopsis. Plant Cell. 2003, 15, 790–800. [Google Scholar] [CrossRef]
- Dynowski, M.; Schaaf, G.; Loque, D.; Moran, O.; Ludewig, U. Plant plasma membrane water channels conduct the signalling molecule H2O2. Biochem. J. 2008, 414, 53–61. [Google Scholar] [CrossRef]
- Kouadio, J.K.; Kouakou, H.T.; Kone, M.; Zouzou, M.; Anno, P.A. Optimum conditions for cotton nitrate reductase extraction and activity measurement. Afr. J. Biotechnol. 2007, 6, 923–928. [Google Scholar]
- Cao, Y.; Fan, X.R.; Sun, S.B.; Xu, G.H.; Hu, J.; Shen, Q.R. Effect of nitrate on activities and transcript levels of nitrate reductase and glutamine synthetase in rice. Pedosphere 2008, 18, 664–673. [Google Scholar] [CrossRef]
- Rosales, E.P.; Iannone, M.F.; Groppa, M.D.; Benavides, M.P. Nitric oxide inhibits nitrate reductase activity in wheat leaves. Plant Physiol. Biochem. 2011, 49, 124–130. [Google Scholar] [CrossRef]
- Cánovas, F.M.; Conceptión, A.; Cantón, F.R.; Cañas, R.A.; De La Torre, F. Ammonium assimilation and amino acid metabolism in conifers. J. Exp. Bot. 2007, 58, 2307–2318. [Google Scholar] [CrossRef]
- Lea, P.J.; Miflin, B.J. Alternative route for nitrogen assimilation in higher plants. Nature 1974, 251, 614–616. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechrgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef]
- Vanoni, M.A.; Dossena, L.; van den Heuvel, R.H.H.; Curti, B. Structure-function studies on the complex iron-sulfur flavoprotein glutamate synthase: The key enzyme of ammonium assimilation. Photosynth. Res. 2005, 83, 219–238. [Google Scholar] [CrossRef]
- Lam, H.M.; Wong, P.; Chan, H.K.; Yam, K.M.; Chen, L.; Chow, C.M.; Coruzzi, G.M. Overexpression of the ASN1 gene enhances nitrogen status in seeds of Arabidopsis. Plant Physiol. 2003, 132, 926–935. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Pageau, K.; Lelandais, M.; Grandjean, O.; Kronenberger, J.; Valadier, M.H.; Jouglet, T.; Suzuki, A. Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco. Plant Physiol. 2006, 140, 444–456. [Google Scholar] [CrossRef]
- Potel, F.; Valadier, M.H.; Ferrario-Méry, S.; Grandjean, O.; Gaufichon, L.; Boutet-Mercey, S.; Lothier, J.; Rothstein, S.J.; Hirose, N.; Suzuki, A. Assimilation of excess ammonium into amino acids and nitrogen translocation in Arabidopsis thaliana—Roles of glutamate synthases and carbamoylphosphate synthetase in leaves. FEBS J. 2009, 276, 4061–4076. [Google Scholar] [CrossRef]
- Skopelitis, D.S.; Paranychianakis, N.V.; Paschalidis, K.A.; Pliakonis, E.D.; Delis, I.D.; Yakoumakis, D.I.; Kouvarakis, A.; Ppadakis, A.K.; Stephanou, E.G.; Roubelakis-Angelakis, K.A. Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenase to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell. 2006, 18, 2767–2781. [Google Scholar] [CrossRef]
- Ota, K. Stimulation of CAM photosynthesis in Kalanchoë blossfeldiana by transferring to nitrogen-deficient conditions. Plant Physiol. 1988, 87, 454–457. [Google Scholar] [CrossRef]
- Pereira, P.N.; Smith, J.A.C.; Mercier, H. Nitrate enhancement of CAM activity in two Kalanchoë species is associated with increased vacuolar proton transport capacity. Physiol Plant. 2017, 160, 361–372. [Google Scholar] [CrossRef]
- Pereira, P.N.; Gaspar, M.; Smith, J.A.C.; Mercier, H. Ammonium intensifies CAM photosynthesis and counteracts drought effects by increasing malate transport and antioxidant capacity in Guzmania monostachia. J. Exp. Bot. 2018, 69, 1993–2003. [Google Scholar] [CrossRef]
- Ota, K.; Tezuka, T.; Yamamoto, Y. Changes in crassulacean acid metabolism of Kalanchoë blossfeldiana by different nitrogen sources. Plant Cell Physiol. 1988, 29, 533–537. [Google Scholar]
- Ota, K.; Yamamoto, Y. Effects of different nitrogen sources and concentrations on CAM photosynthesis in Kalanchoë blossfeldiana. J. Exp. Bot. 1991, 42, 1271–1277. [Google Scholar] [CrossRef]
- Smith, J.A.C.; Uribe, E.G.; Ball, E.; Heuer, S.; Lüttge, U. Characterization of the vacuolar ATPase activity of the CAM plant Kalanchoë daigremontiana Receptor modulating. Eur. J. Biochem. 1984, 141, 415–420. [Google Scholar] [CrossRef]
- Inselsbacher, E.; Cambui, C.A.; Richter, A.; Stange, C.F.; Mercier, H.; Wanek, W. Microbial activities and foliar uptake of nitrogen in the epiphytic bromeliad Vriesea gigantea. New Phytol. 2007, 175, 311–320. [Google Scholar] [CrossRef]
- Osmond, C.B.; Bender, M.M.; Burris, R.H. Pathways of CO2 Fixation in the Cam Plant Kalanchoë daigremontiana. III. Correlation With δ13C Value During Growth and Water Stress. Funct. Plant Biol. 1975, 3, 787–789. [Google Scholar] [CrossRef]
- Schmitt, J.M.; Piepenbrock, M. Regulation of phosphoenolpyruvate carboxylase and crassulacean acid metabolism induction in Mesembryanthemum crystallinum L. by cytokinin: Modulation of leaf gene expression by roots? Plant Physiol. 1992, 99, 1664–1669. [Google Scholar] [CrossRef][Green Version]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arabidopsis Book 2014, 12, e0168. [Google Scholar] [CrossRef]
- Princic, A.; Mahne, I.; Megusar, F.; Paul, E.A.; Tiedje, J.M. Effects of pH and oxygen and ammonium concentrations on the community structure of nitrifying bacteria from wastewater. Appl. Environ. Microbiol. 1998, 64, 3584–3590. [Google Scholar]
- Nieder, R.; Benbi, D.K.; Scherer, H.W. Fixation and defixation of ammonium in soils: A review. Biol. Fertil. Soils 2011, 47, 1–14. [Google Scholar] [CrossRef]
- Cao, X.; Ma, Q.; Wu, L.; Zhu, L.; Jin, Q. Effects of ammonium application rate on uptake of soil adsorbed amino acids by rice. Biomed. Biotechnol. 2016, 17, 294–302. [Google Scholar] [CrossRef]
- Angle, J.S.; Gross, C.M.; Hill, R.L.; McIntosh, M.S. Soil nitrate concentrations under corn as affected by tillage, manure, and fertilizer applications. J. Environ. Qual. 1993, 22, 141–147. [Google Scholar] [CrossRef]
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils; Pearson Education: London, UK, 1996. [Google Scholar]
- Uenishi, Y.; Nakabayashi, Y.; Tsuchihira, A.; Takusagawa, M.; Hashimoto, K.; Maeshima, M.; Sato-Nara, K. Accumulation of tip2;2 aquaporin during dark adaptation is partially PhyA dependent in roots of Arabidopsis seedlings. Plants 2014, 3, 177–195. [Google Scholar] [CrossRef]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signaling pathways and transcriptome response signatures differentiate ammonium- and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Jahn, T.P.; Moller, A.L.B.; Zeuthen, T.; Holm, L.M.; klaerke, D.A.; Mohsin, B.; Kühlbrandt, W.; Schjoerring, J.K. Aquaporin homologues in plants and mammals transport ammonia. FEBS Lett. 2004, 574, 31–36. [Google Scholar] [CrossRef]
- Li, B.; Li, G.; Kronzucker, H.J.; Baluska, F.; Shi, W. Ammonium stress in Arabidopsis signaling, genetic loci, and physiological targets. Trends Plant Sci. 2014, 19, 107–114. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Britto, D.T.; Davenport, R.J.; Tester, M. Ammonium toxicity and the real cost of transport. Trends Plant Sci. 2001, 6, 335–337. [Google Scholar] [CrossRef]
- Hachiya, T.; Watanabe, C.K.; Boom, C.; Tholen, D.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Ammonium-dependent respiratory increase is dependent on the cytochrome pathway in Arabidopsis thaliana shoots. Plant Cell Environ. 2010, 33, 1888–1897. [Google Scholar] [CrossRef]
- Bittsánszky, A.; Pilinszky, K.; Gyulai, G.; Komives, T. Overcoming ammonium toxicity. Plant Sci. 2015, 231, 184–190. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Ding, L.; Shen, Q.; Guo, S. Ammonium enhances the tolerance of rice seedlings (Oryza sativa L.) to drought condition. Agric. Water Manag. 2009, 96, 1746–1750. [Google Scholar] [CrossRef]
- Hessini, K.; Hamed, K.B.; Gandour, M.; Mejri, M.; Abdelly, C.; Cruz, C. Ammonium nutrition in the halophyte Spartina alterniflora under salt stress: Evidence for a priming effect of ammonium? Plant Soil. 2013, 370, 163–173. [Google Scholar] [CrossRef]
- Ding, L.; Gao, C.; Li, Y.; Li, Y.; Zhu, Y.; Xu, G.; Shen, Q.; Kaldenhoff, R.; Kai, L.; Guo, S. The enhanced drought tolerance of rice plants under ammonium is related to aquaporin (AQP). Plant Sci. 2015, 234, 14–21. [Google Scholar] [CrossRef]
- Winter, K.; Usuda, H.; Tsuzuki, M.; Schmitt, M.; Edwards, G.E.; Thomas, R.J.; Evert, R.F. Influence of nitrate and ammonia on photosynthetic characteristics and leaf anatomy of Moricandia arvensis. Plant Physiol. 1982, 70, 616–625. [Google Scholar] [CrossRef]
- Cramer, M.D.; Lewis, O.A. The influence of nitrate and ammonium nutrition on growth of wheat (Triticum aestivum) and maize (Zea mays) plants. Ann. Bot. 1993, 72, 359–365. [Google Scholar] [CrossRef]
- Gerendas, J.; Zhu, Z.; Bendixen, R.; Ratcliffe, R.G.; Sattelmacher, B. Physiological and Biochemical processes related to ammonium toxicity in higher plants. Z. Pflonzenerniihr. Bodenk. 1997, 160, 239–251. [Google Scholar] [CrossRef]
- Britto, D.T.; Siddiqi, M.Y.; Glass, A.D.M.; Kronzucker, H.J. Futile transmembrane NH4+ cycling: A cellular hypothesis to explain ammonium toxicity in plants. Plant Biol. 2001, 98, 4255–4258. [Google Scholar] [CrossRef]
- Lightfoot, D.A.; Mungur, R.; Ameziane, R.; Nolte, S.; Long, L.; Bernhard, K.; Colter, A.; Jones, K.; Iqbal, M.J.; Varsa, E.; et al. Improved drought tolerance of transgenic Zea mays plants that express the glutamate dehydrogenase gene (gdhA) of E. coli. Euphytica 2007, 156, 103–116. [Google Scholar] [CrossRef]
- Cao, X.; Zhong, C.; Zhu, C.; Zhu, L.; Zhang, J.; Wu, L.; Jin, Q. Ammonium uptake and metabolism alleviate PEG-induced water stress in rice seedlings. Plant Physiol. Biochem. 2018, 132, 128–137. [Google Scholar] [CrossRef]
- Cambui, C.A.; Svennerstam, H.; Gruffman, L.; Nordin, A.; Ganeteg, U.; Nasholm, T. Patterns of Plant Biomass Partitioning Depend on Nitrogen Source. PLoS ONE 2011, 6, e19211. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. Nitrogen acquisition, PEPC carboxylase, and cellular pH homeostasis: New views on old paradigms. Plant Cell Environ. 2005, 28, 1396–1409. [Google Scholar] [CrossRef]
- Liu, L.H.; Ludewig, U.; Gassert, B.; Frommer, W.B.; Wirén, N.V. Urea Transport by Nitrogen-Regulated Tonoplast Intrinsic Proteins in Arabidopsis. Plant Physiol. 2003, 133, 1220–1228. [Google Scholar] [CrossRef]
- Cambui, C.A.; Gaspar, M.; Mercier, H. Detection of urease in the cell wall and membranes from leaf tissues of bromeliad species. Physiol. Plant. 2009, 136, 86–93. [Google Scholar] [CrossRef]
- Dignon, J.; Hameed, S. Global emissions of nitrogen and sulfur-Ooxides from 1860 to 1980. JAPCA 1989, 39, 180–189. [Google Scholar] [CrossRef]
- Ackerman, D.; Millet, D.B.; Chen, X. Global estimates of inorganic deposition across four decades. Glob. Biogeochem. Cycles 2019, 33, 100–107. [Google Scholar] [CrossRef]
- Payne, R.J.; Dise, N.B.; Field, C.D.; Dore, A.J.; Carpon, S.J.; Stevens, C.J. Nitrogen deposition and plant biodiversity: Past, present, and future. Front. Ecol. Environ. 2017, 15, 431–436. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef]
- Valliere, J.M.; Irvine, I.C.; Allen, E.B. High N, dry: Experimental nitrogen deposition exacerbates native shrub loss and nonnative plant invasion during extreme drought. Glob. Chang. Biol. 2017, 23, 23–4333. [Google Scholar] [CrossRef]
- Reynolds, H.; D’antonio, C. The ecological significance of plasticity in root weight ratio in response to nitrogen: Opinion. Plant Soil 1996, 185, 75–97. [Google Scholar] [CrossRef]
- Friedrich, U.; von Oheimb, G.; Kriebitzsch, W.U.; Schlesselmann, K.; Weber, M.S.; Hardtle, W. Nitrogen deposition increases susceptibility to drought—Experimental evidence with the perennial grass Molinia caerulea (L) Moench. Plant Soil 2012, 353, 59–71. [Google Scholar] [CrossRef]
- Raven, J.A.; Spicer, R.A. The evolution of Crassulacean acid metabolism. In Crassulacean Acid Metabolism—Biochemistry, Ecophysiology and Evolution; Winter, K., Smith, J.A.C., Eds.; Springer-Verlag: Berlin, Germany, 1996; Volume 114, pp. 360–385. [Google Scholar]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, P.N.; Cushman, J.C. Exploring the Relationship between Crassulacean Acid Metabolism (CAM) and Mineral Nutrition with a Special Focus on Nitrogen. Int. J. Mol. Sci. 2019, 20, 4363. https://doi.org/10.3390/ijms20184363
Pereira PN, Cushman JC. Exploring the Relationship between Crassulacean Acid Metabolism (CAM) and Mineral Nutrition with a Special Focus on Nitrogen. International Journal of Molecular Sciences. 2019; 20(18):4363. https://doi.org/10.3390/ijms20184363
Chicago/Turabian StylePereira, Paula Natália, and John C. Cushman. 2019. "Exploring the Relationship between Crassulacean Acid Metabolism (CAM) and Mineral Nutrition with a Special Focus on Nitrogen" International Journal of Molecular Sciences 20, no. 18: 4363. https://doi.org/10.3390/ijms20184363
APA StylePereira, P. N., & Cushman, J. C. (2019). Exploring the Relationship between Crassulacean Acid Metabolism (CAM) and Mineral Nutrition with a Special Focus on Nitrogen. International Journal of Molecular Sciences, 20(18), 4363. https://doi.org/10.3390/ijms20184363