β-catenin/LEF1/IGF-IIR Signaling Axis Galvanizes the Angiotensin-II- induced Cardiac Hypertrophy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
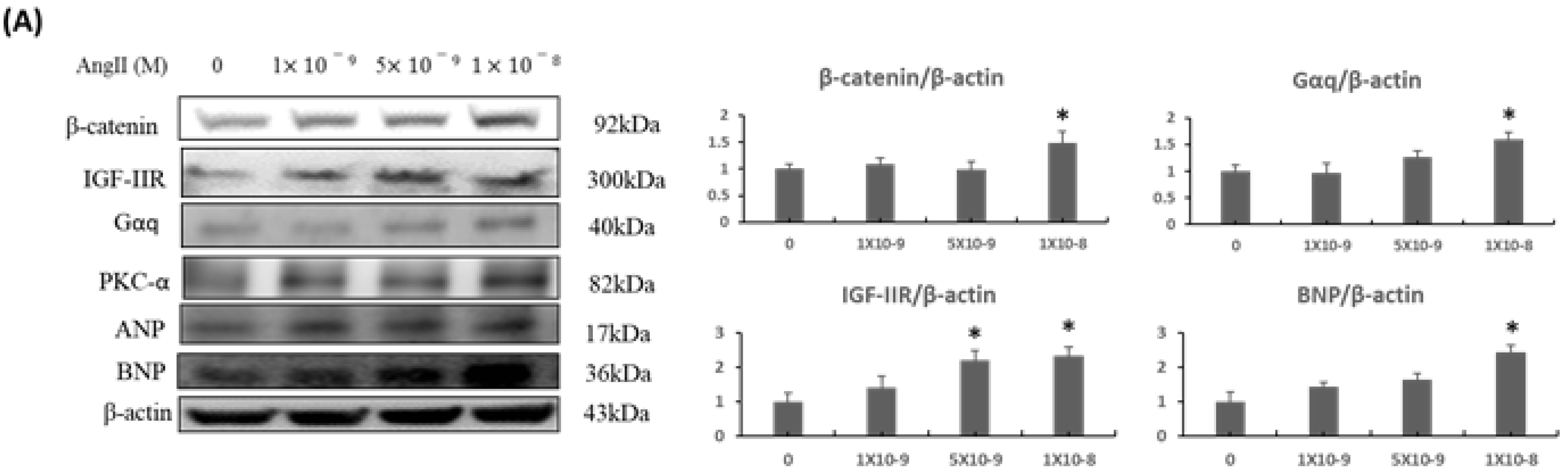
2.1. β-Catenin Expression Was Increased upon Ang-II Treatment in a Dose-Dependent Manner and Activated IGF-IIR/Gaq Mediated Cardiac Hypertrophy Pathway in H9c2 Cardiomyoblasts
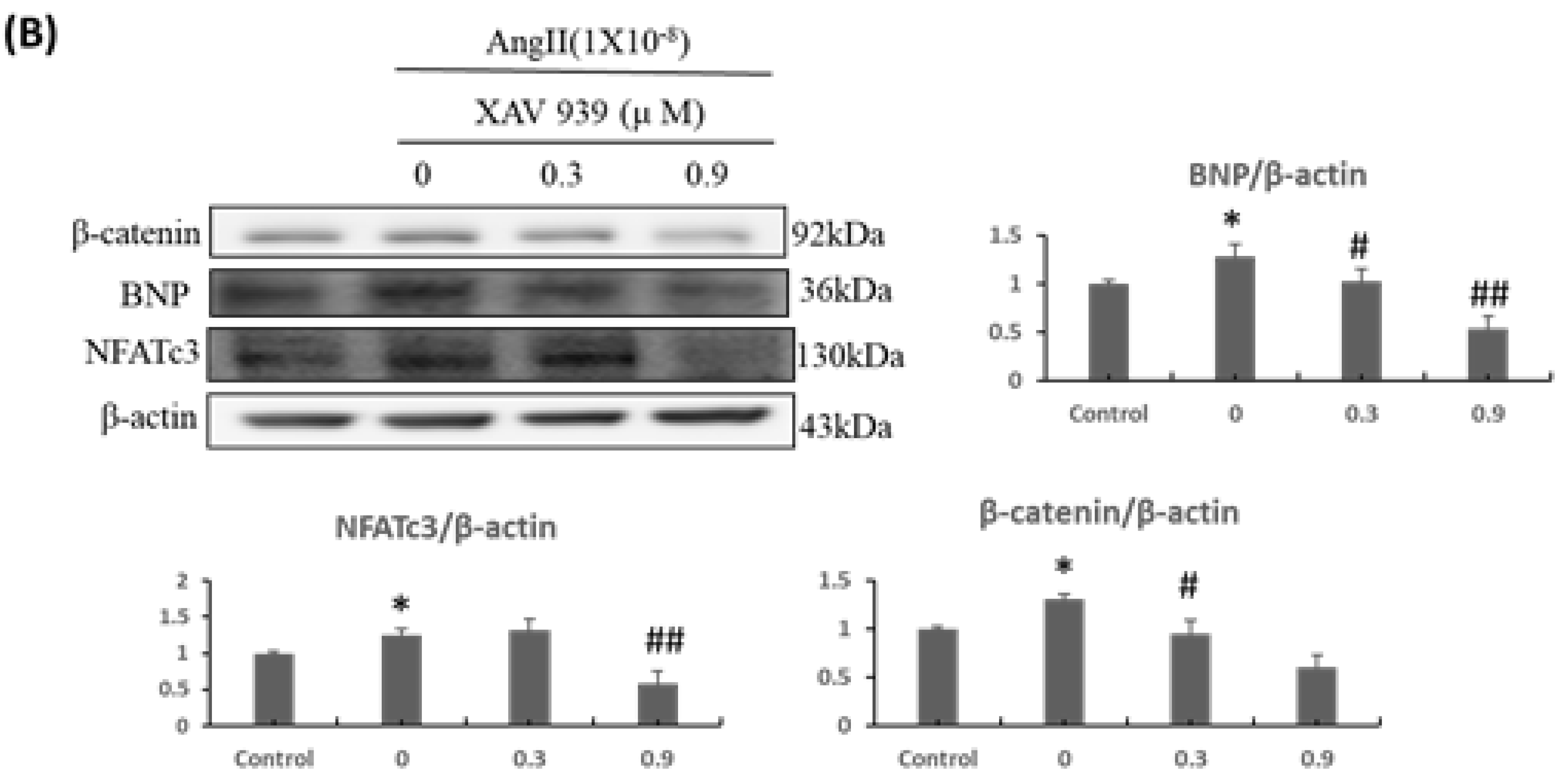
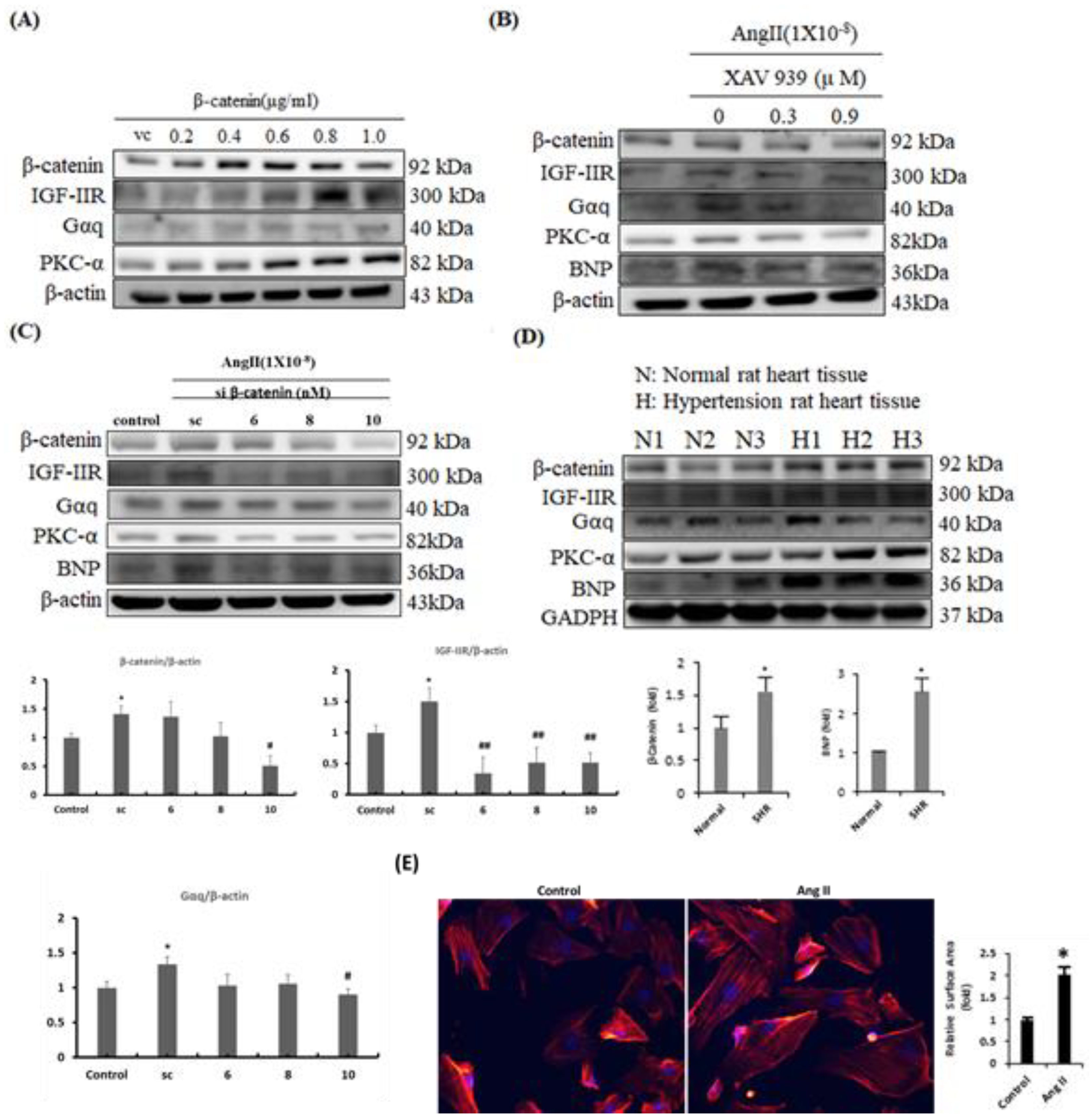
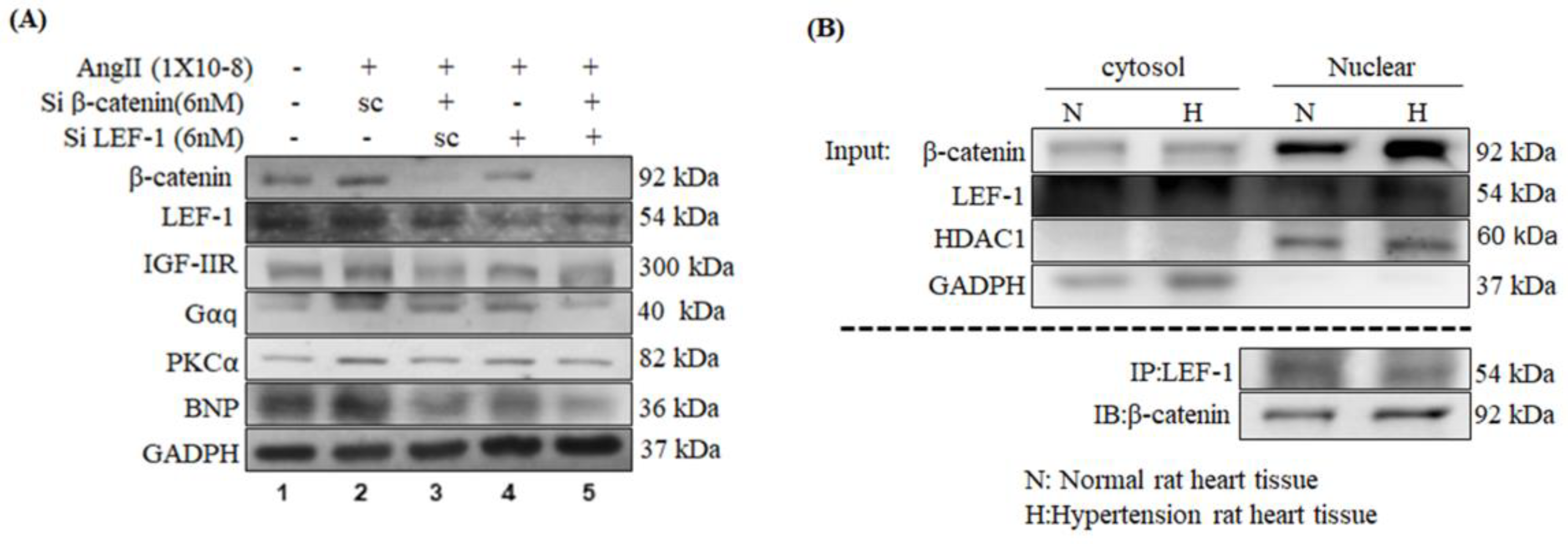
2.2. β-Catenin Alterations Modulated the Cardiac Hypertrophy Associated Signaling Effectors both In Vitro and In Vivo
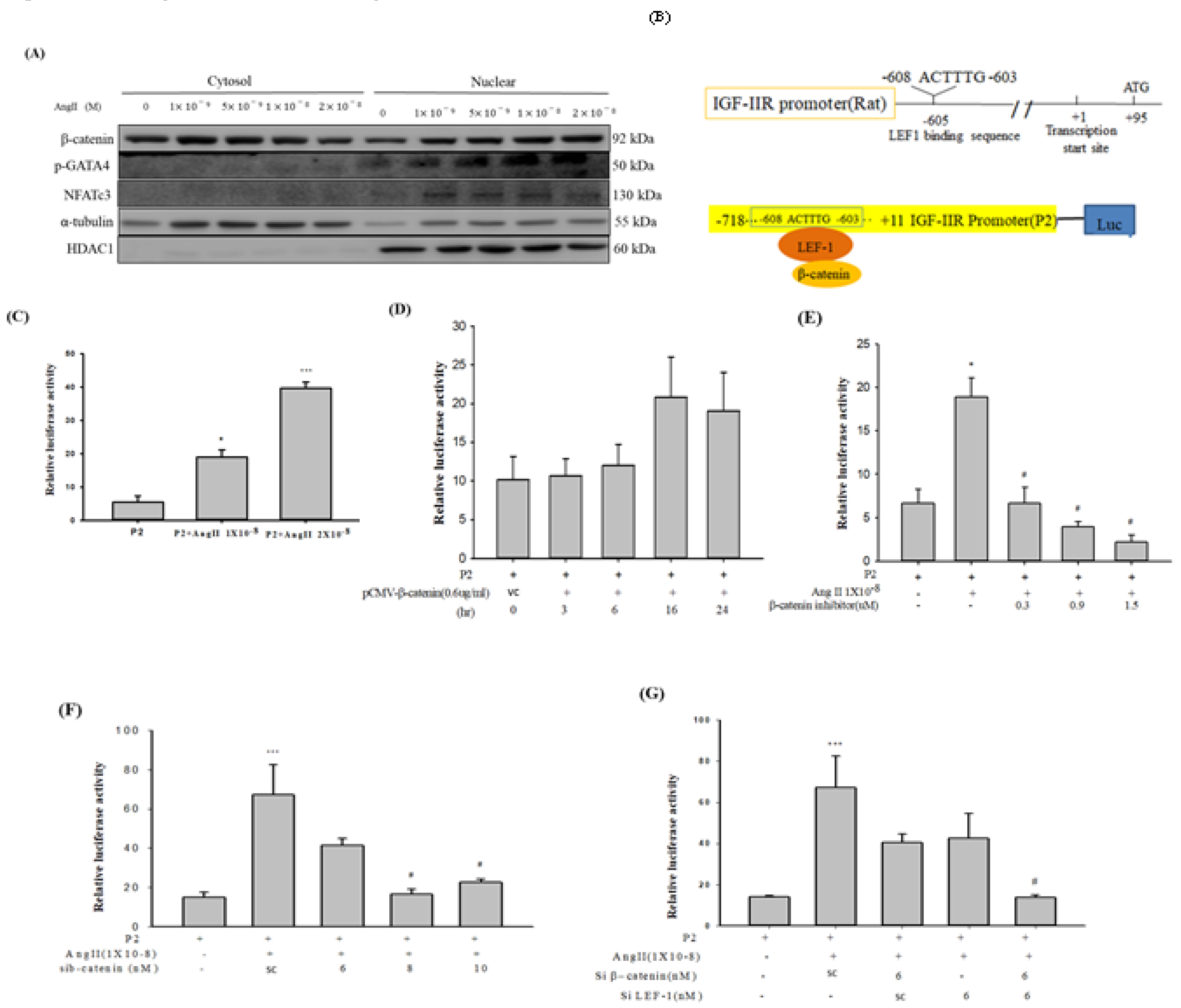
2.3. Ang-II Treatment Enhanced Nuclear Enrichment of β-catenin, GATA-4 and NFATc3 in H9c2 Cardiomyoblast Cells
2.4. IGF-IIR Promoter was Regulated by β-catenin and LEF1
2.5. Inhibition of β-Catenin and/or LEF1 Alleviated the Hypertrophic Effect induced by Ang-II Treatment via Suppressing IGF-IIR/Gαq/PKCα Pathway
3. Methodology
3.1. Cell Culture
3.2. Animal Model
3.3. Western Blotting
3.4. Actin Staining
3.5. Immunoprecipitations (IP)
3.6. Luciferase Assay
3.7. Nuclear Protein Extraction
3.8. Statistical Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Mills, K.T.; Bundy, J.D.; Kelly, T.N.; Reed, J.E.; Kearney, P.M.; Reynolds, K.; Chen, J.; He, J. Global Disparities of Hypertension Prevalence and Control: A Systematic Analysis of Population-Based Studies from 90 Countries. Circulation 2016, 134, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Blankesteijn, W.M.; van de Schans, V.A.; ter Horst, P.; Smits, J.F. The Wnt/frizzled/GSK-3 beta pathway: A novel therapeutic target for cardiac hypertrophy. Trends Pharmacol. Sci. 2008, 29, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Kuo, W.W.; Yeh, Y.L.; Ho, T.J.; Lin, J.Y.; Lin, D.Y.; Chu, C.H.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y. ANG II promotes IGF-IIR expression and cardiomyocyte apoptosis by inhibiting HSF1 via JNK activation and SIRT1 degradation. Cell Death Differ. 2014, 21, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.S.; Rocic, P.; Mellis, A.M.; Laude, K.; Lyle, A.N.; Harrison, D.G.; Griendling, K.K. Angiotensin II-induced hypertrophy is potentiated in mice overexpressing p22phox in vascular smooth muscle. Am. J. Physiol.-Heart Circ. Physiol. 2005, 288, H37–H42. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.C.; Pandey, S.; Lin, C.Y.; Shen, C.Y.; Chang, R.L.; Chang, T.T.; Chen, R.J.; Viswanadha, V.P.; Lin, Y.M.; Huang, C.Y. Cardiac apoptosis induced under high glucose condition involves activation of IGF2R signaling in H9c2 cardiomyoblasts and streptozotocin-induced diabetic rat hearts. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 97, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.S.; Wang, H.F.; Pai, P.Y.; Jong, G.P.; Lai, C.H.; Chung, L.C.; Hsieh, D.J.; HsuanDay, C.; Kuo, W.W.; Huang, C.Y. Tanshinone IIA Prevents Leu27IGF-II-Induced Cardiomyocyte Hypertrophy Mediated by Estrogen Receptor and Subsequent Akt Activation. Am. J. Chin. Med. 2015, 43, 1567–1591. [Google Scholar] [CrossRef] [PubMed]
- ter Horst, P.; Smits, J.F.M.; Blankesteijn, W.M. The Wnt/Frizzled pathway as a therapeutic target for cardiac hypertrophy: Where do we stand? Acta Physiol. 2012, 204, 110–117. [Google Scholar] [CrossRef]
- Naito, A.T.; Shiojima, I.; Akazawa, H.; Hidaka, K.; Morisaki, T.; Kikuchi, A.; Komuro, I. Developmental stage-specific biphasic roles of Wnt/β-catenin signaling in cardiomyogenesis and hematopoiesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19812–19817. [Google Scholar] [CrossRef]
- Baurand, A.; Zelarayan, L.; Betney, R.; Gehrke, C.; Dunger, S.; Noack, C.; Busjahn, A.; Huelsken, J.; Taketo, M.M.; Birchmeier, W.; et al. Beta-catenin downregulation is required for adaptive cardiac remodeling. Circ. Res. 2007, 100, 1353–1362. [Google Scholar] [CrossRef]
- Chen, B.C.; Hung, M.Y.; Wang, H.F.; Yeh, L.J.; Pandey, S.; Chen, R.J.; Chang, R.L.; Viswanadha, V.P.; Lin, K.H.; Huang, C.Y. GABA tea attenuates cardiac apoptosis in spontaneously hypertensive rats (SHR) by enhancing PI3K/Akt-mediated survival pathway and suppressing Bax/Bak dependent apoptotic pathway. Environ. Toxicol. 2018, 33, 789–797. [Google Scholar] [CrossRef]
- Wu, M.H.; Lee, T.H.; Lee, H.P.; Li, T.M.; Lee, I.T.; Shieh, P.C.; Tang, C.H. Kuei-Lu-Er-Xian-Jiao extract enhances BMP-2 production in osteoblasts. BioMedicine 2017, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Kuo, W.W.; Shen, C.Y.; Yeh, Y.L.; Ho, T.J.; Chen, R.J.; Chang, R.L.; Pai, P.Y.; Viswanadha, V.P.; Huang, C.Y.; et al. IGF-IIRalpha is a novel stress-inducible contributor to cardiac damage underpinning doxorubicin-induced oxidative stress and perturbed mitochondrial autophagy. Am. J. Physiol. Cell Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Kuo, W.W.; Ho, T.J.; Yeh, Y.L.; Shen, C.Y.; Chen, R.J.; Chang, R.L.; Pai, P.Y.; Padma, V.V.; Huang, C.Y.; et al. Upregulation of IGF-IIRalpha intensifies doxorubicin-induced cardiac damage. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Kuo, W.W.; Baskaran, R.; Chen, M.C.; Ho, T.J.; Chen, R.J.; Chen, Y.F.; Vijaya Padma, V.; Lay, I.S.; Huang, C.Y. Enhancement of beta-catenin in cardiomyocytes suppresses survival protein expression but promotes apoptosis and fibrosis. Cardiol. J. 2017, 24, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; De Windt, L.J.; Witt, S.A.; Kimball, T.R.; Markham, B.E.; Molkentin, J.D. The transcription factors GATA4 and GATA6 regulate cardiomyocyte hypertrophy in vitro and in vivo. J. Biol. Chem. 2001, 276, 30245–30253. [Google Scholar] [CrossRef] [PubMed]
- Molkentin, J.D. Calcineurin-NFAT signaling regulates the cardiac hypertrophic response in coordination with the MAPKs. Cardiovasc. Res. 2004, 63, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.; Michael, A.; Andreucci, M.; Bhattacharya, K.; Dotto, P.; Walters, B.; Woodgett, J.; Kilter, H.; Force, T. Stabilization of beta-catenin by a Wnt-independent mechanism regulates cardiomyocyte growth. Proc. Natl. Acad. Sci. USA 2003, 100, 4610–4615. [Google Scholar] [CrossRef] [PubMed]
- Deb, A. Cell–cell interaction in the heart via Wnt/β-catenin pathway after cardiac injury. Cardiovasc. Res. 2014, 102, 214–223. [Google Scholar] [CrossRef]
- Ozhan, G.; Weidinger, G. Wnt/beta-catenin signaling in heart regeneration. Cell Regen. 2015, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Kuo, W.W.; Baskaran, R.; Day, C.H.; Pai, P.Y.; Lai, C.H.; Chen, Y.F.; Chen, R.J.; Padma, V.V.; Huang, C.Y. Increased beta-catenin accumulation and nuclear translocation are associated with concentric hypertrophy in cardiomyocytes. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2017, 31, 9–16. [Google Scholar] [CrossRef]
- Lorenzon, A.; Calore, M.; Poloni, G.; De Windt, L.J.; Braghetta, P.; Rampazzo, A. Wnt/beta-catenin pathway in arrhythmogenic cardiomyopathy. Oncotarget 2017, 8, 60640–60655. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, C.; Wang, C.; Hong, X.; Miao, J.; Liao, Y.; Zhou, L.; Liu, Y. An essential role for Wnt/β-catenin signaling in mediating hypertensive heart disease. Sci. Rep. 2018, 8, 8996. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, C.-H.; Pandey, S.; Day, C.H.; Ho, T.-J.; Chen, R.-J.; Chang, R.-L.; Pai, P.-Y.; Padma, V.V.; Kuo, W.-W.; Huang, C.-Y. β-catenin/LEF1/IGF-IIR Signaling Axis Galvanizes the Angiotensin-II- induced Cardiac Hypertrophy. Int. J. Mol. Sci. 2019, 20, 4288. https://doi.org/10.3390/ijms20174288
Lai C-H, Pandey S, Day CH, Ho T-J, Chen R-J, Chang R-L, Pai P-Y, Padma VV, Kuo W-W, Huang C-Y. β-catenin/LEF1/IGF-IIR Signaling Axis Galvanizes the Angiotensin-II- induced Cardiac Hypertrophy. International Journal of Molecular Sciences. 2019; 20(17):4288. https://doi.org/10.3390/ijms20174288
Chicago/Turabian StyleLai, Chin-Hu, Sudhir Pandey, Cecilia Hsuan Day, Tsung-Jung Ho, Ray-Jade Chen, Ruey-Lin Chang, Pei-Ying Pai, V. Vijaya Padma, Wei-Wen Kuo, and Chih-Yang Huang. 2019. "β-catenin/LEF1/IGF-IIR Signaling Axis Galvanizes the Angiotensin-II- induced Cardiac Hypertrophy" International Journal of Molecular Sciences 20, no. 17: 4288. https://doi.org/10.3390/ijms20174288
APA StyleLai, C.-H., Pandey, S., Day, C. H., Ho, T.-J., Chen, R.-J., Chang, R.-L., Pai, P.-Y., Padma, V. V., Kuo, W.-W., & Huang, C.-Y. (2019). β-catenin/LEF1/IGF-IIR Signaling Axis Galvanizes the Angiotensin-II- induced Cardiac Hypertrophy. International Journal of Molecular Sciences, 20(17), 4288. https://doi.org/10.3390/ijms20174288