Correlation of Autophagosome Formation with Degradation and Endocytosis Arabidopsis Regulator of G-Protein Signaling (RGS1) through ATG8a

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
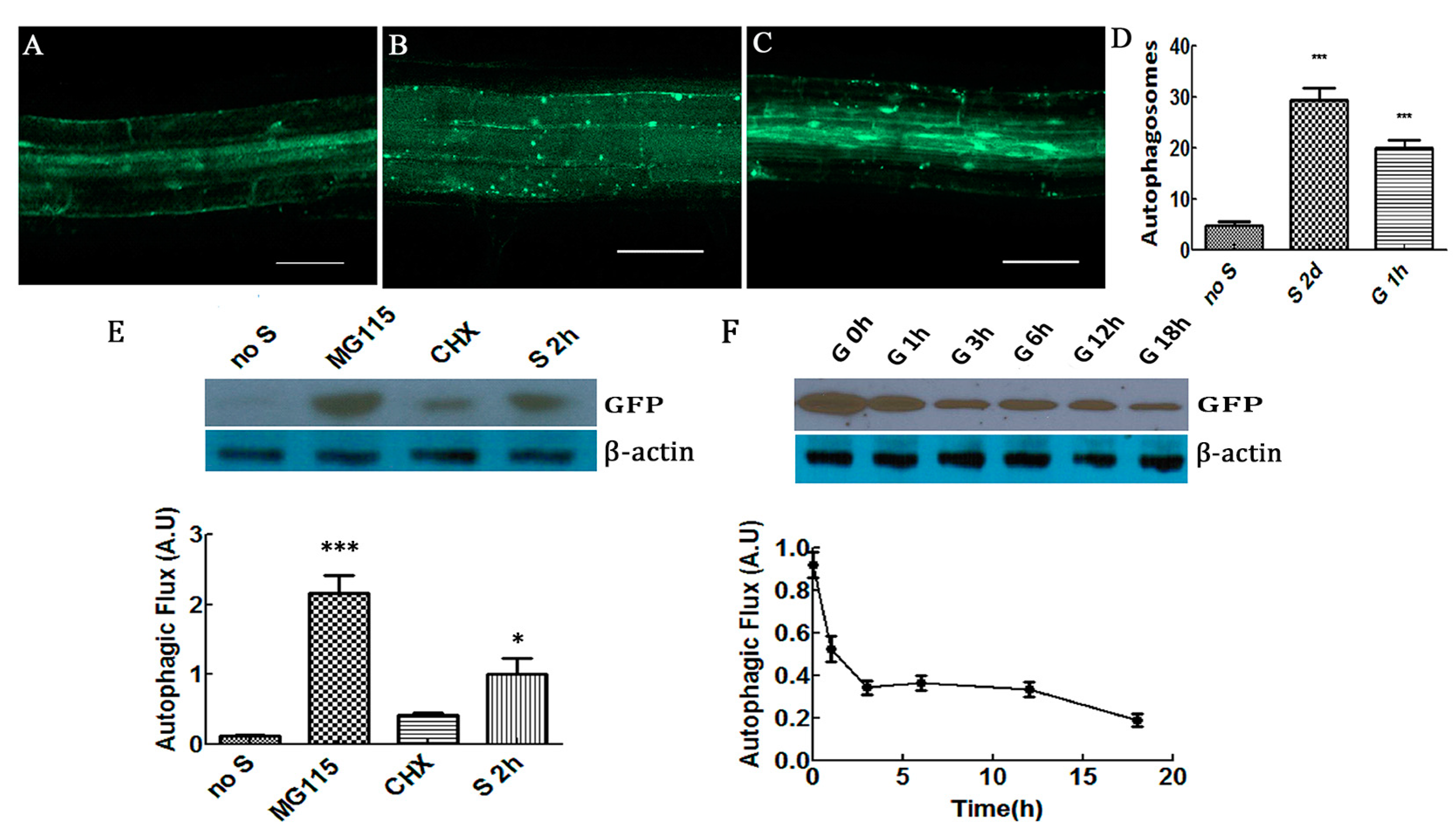
2.1. Glucose Decreased Autophagic Flux and Increased Formation of Autophagosomes in Plant Cells
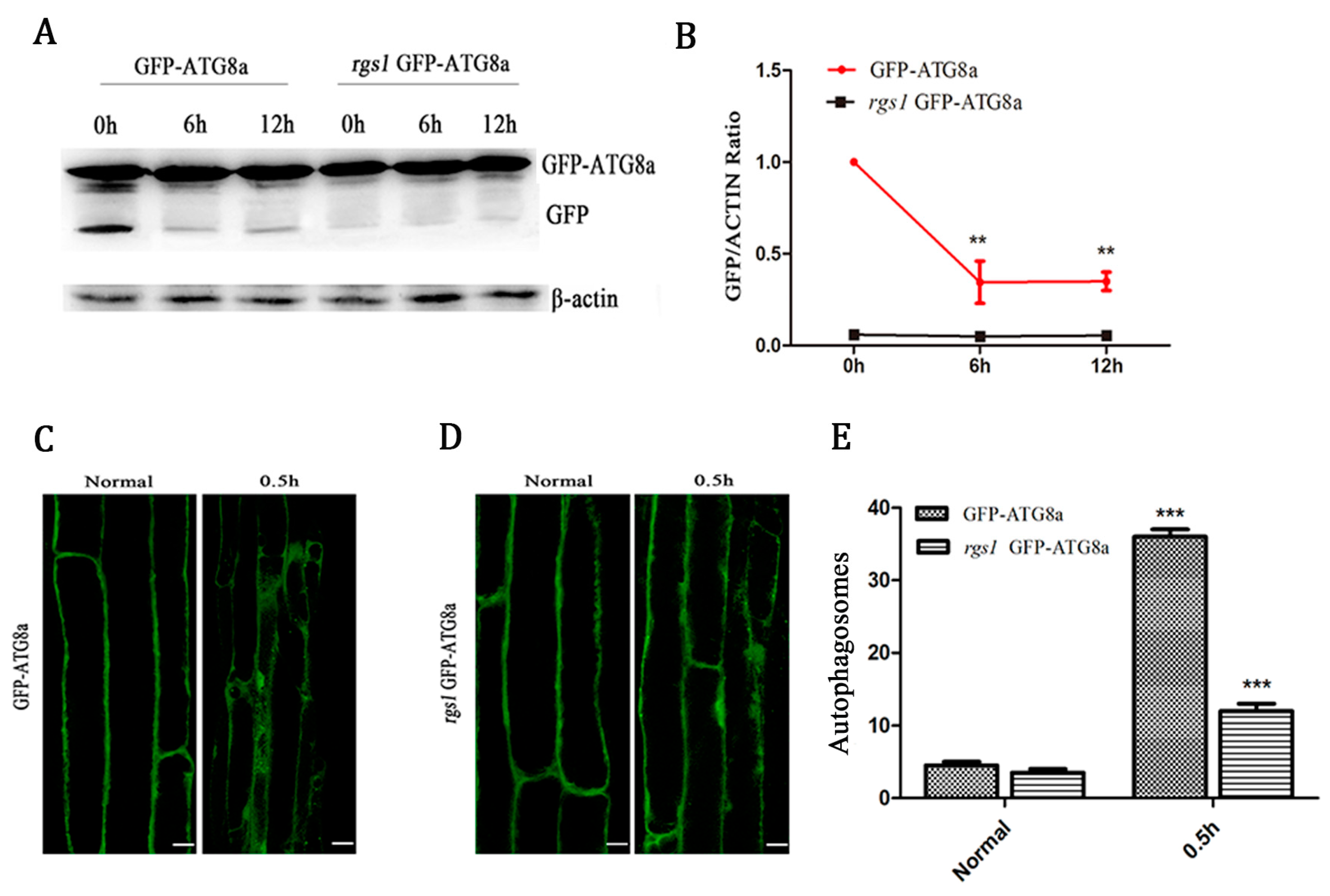
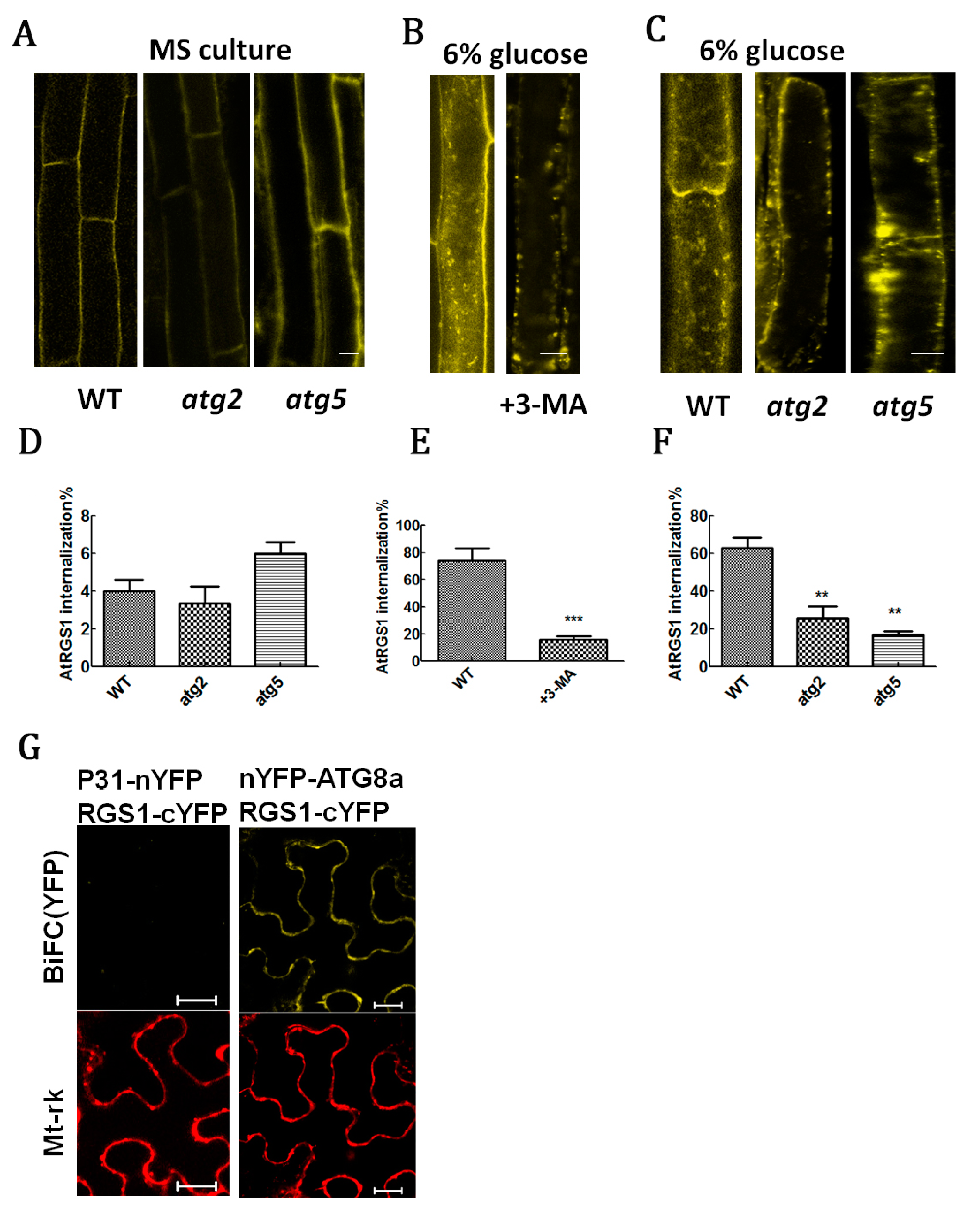
2.2. RGS1 Promoted the Production of Autophagic Flux and Autophagosomes
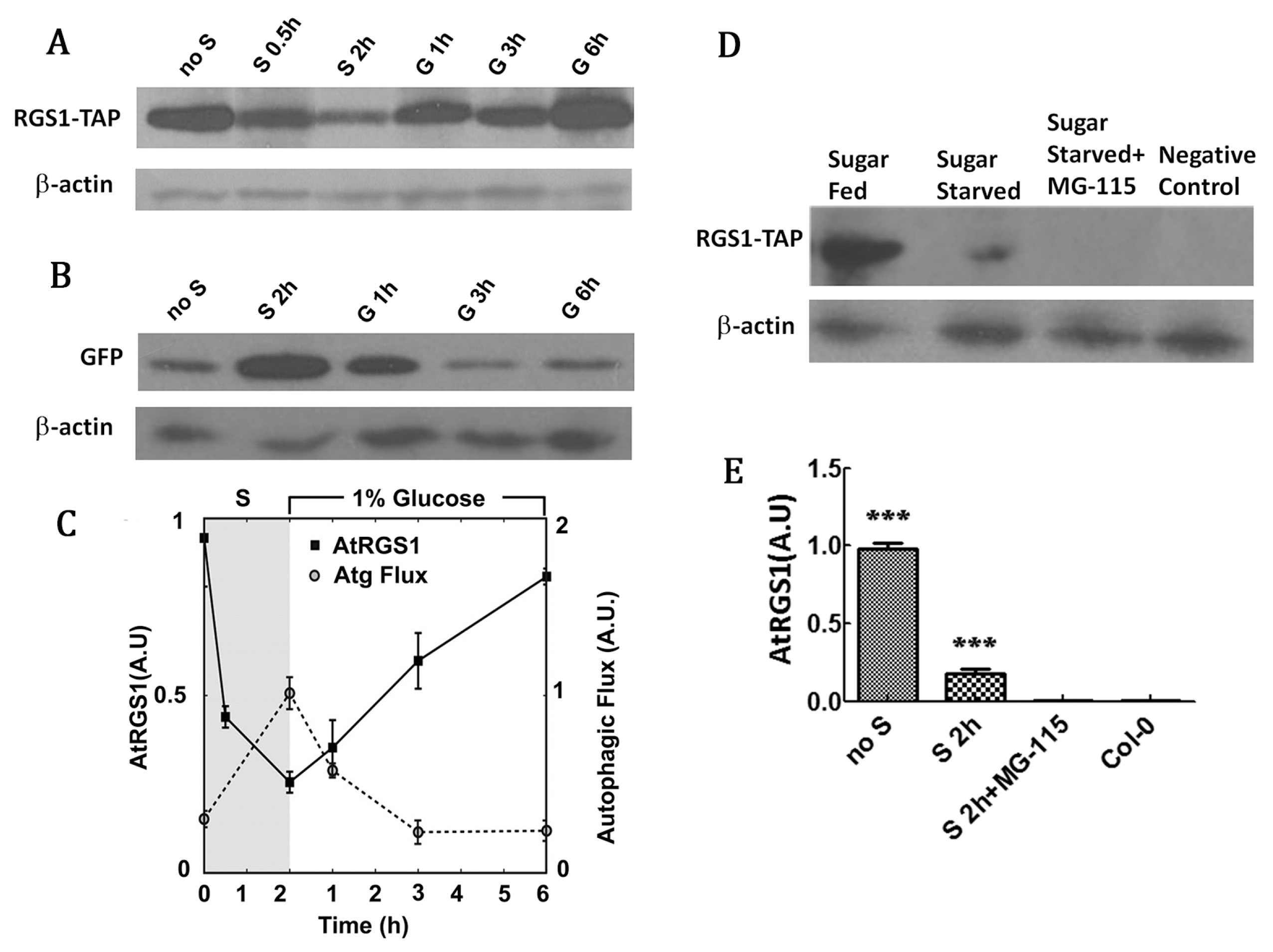
2.3. RGS1 Degradation is Correlated with Increased Autophagic Flux Independent of the Proteasome
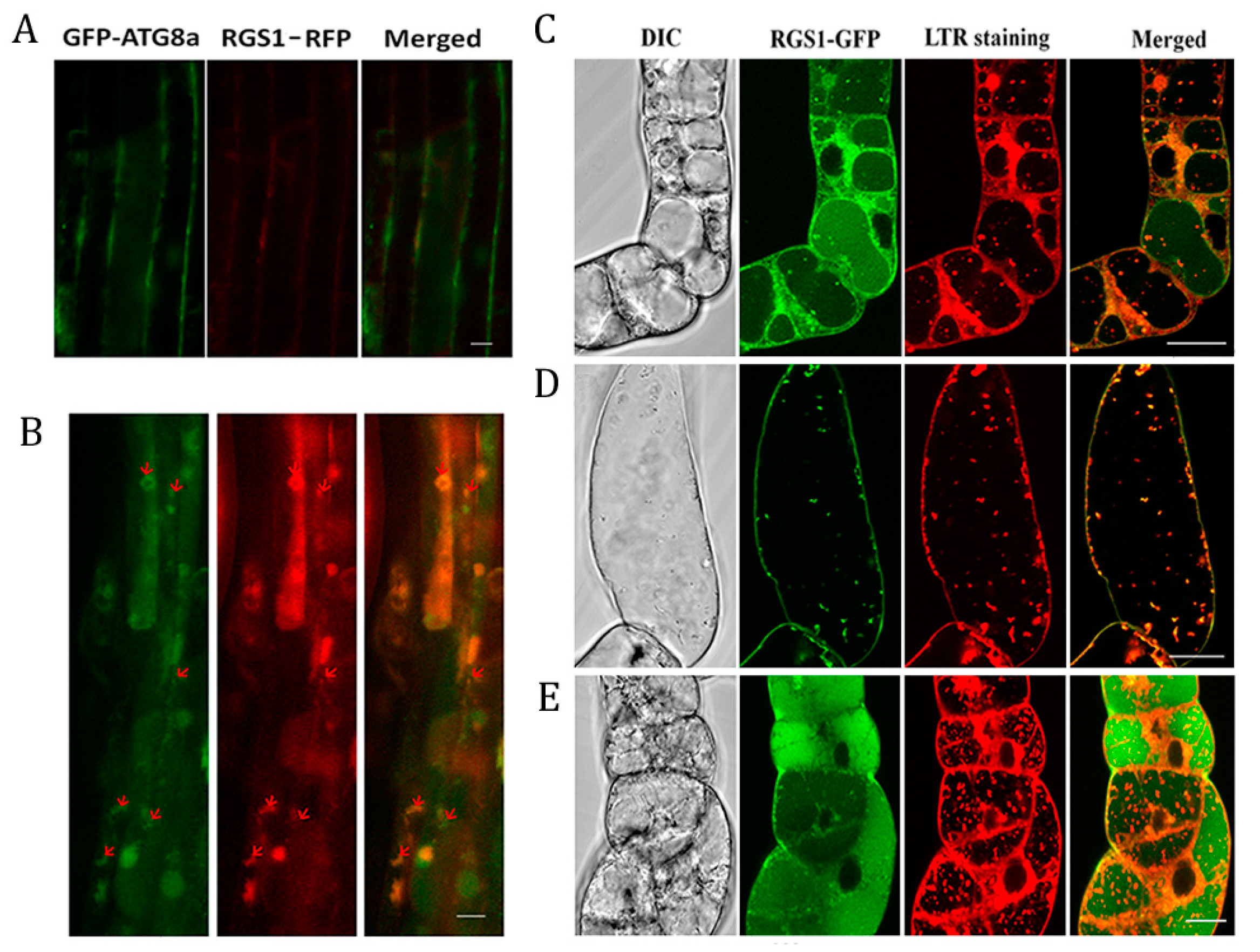
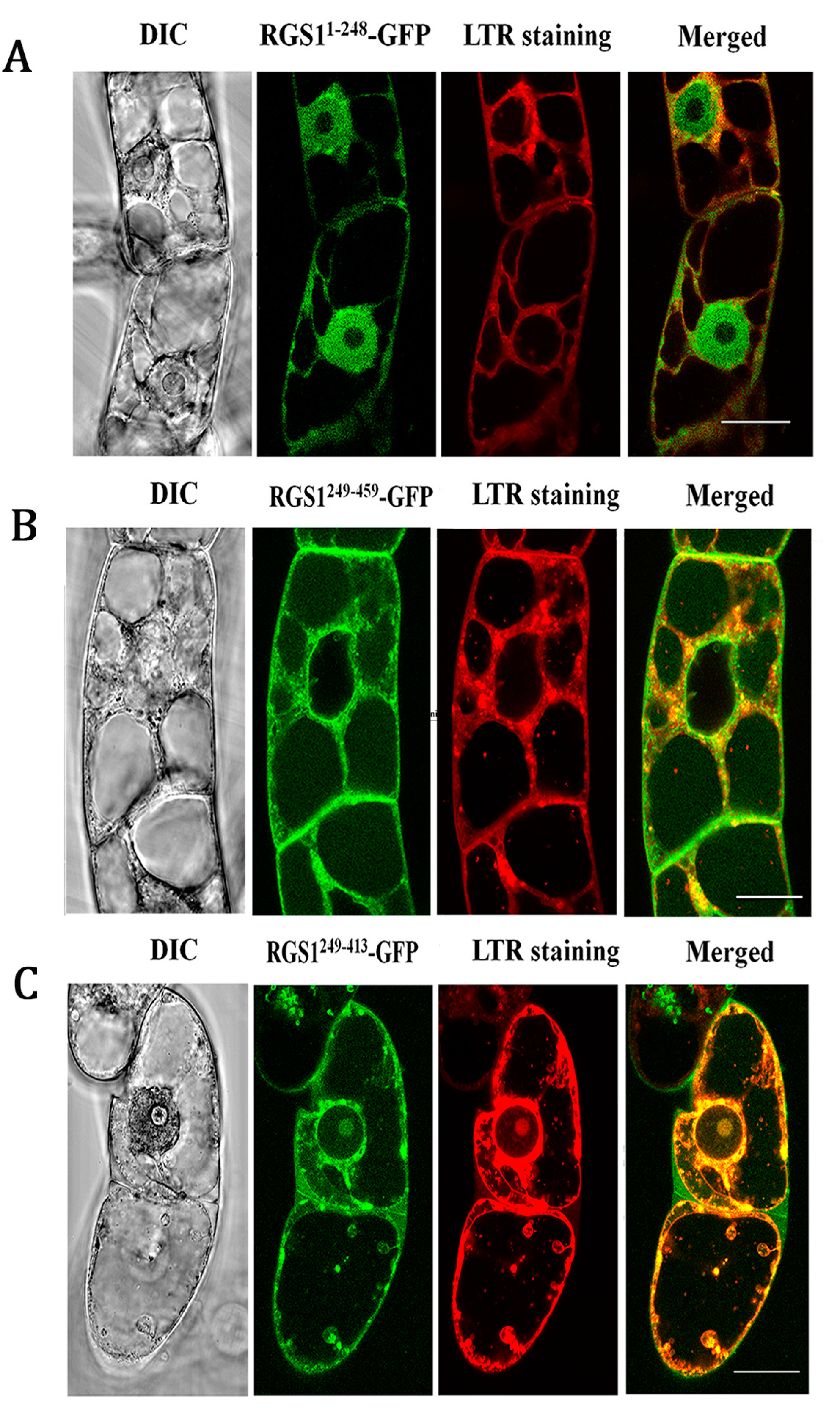
2.4. Co-Localization of RGS1 and Autophagosomes
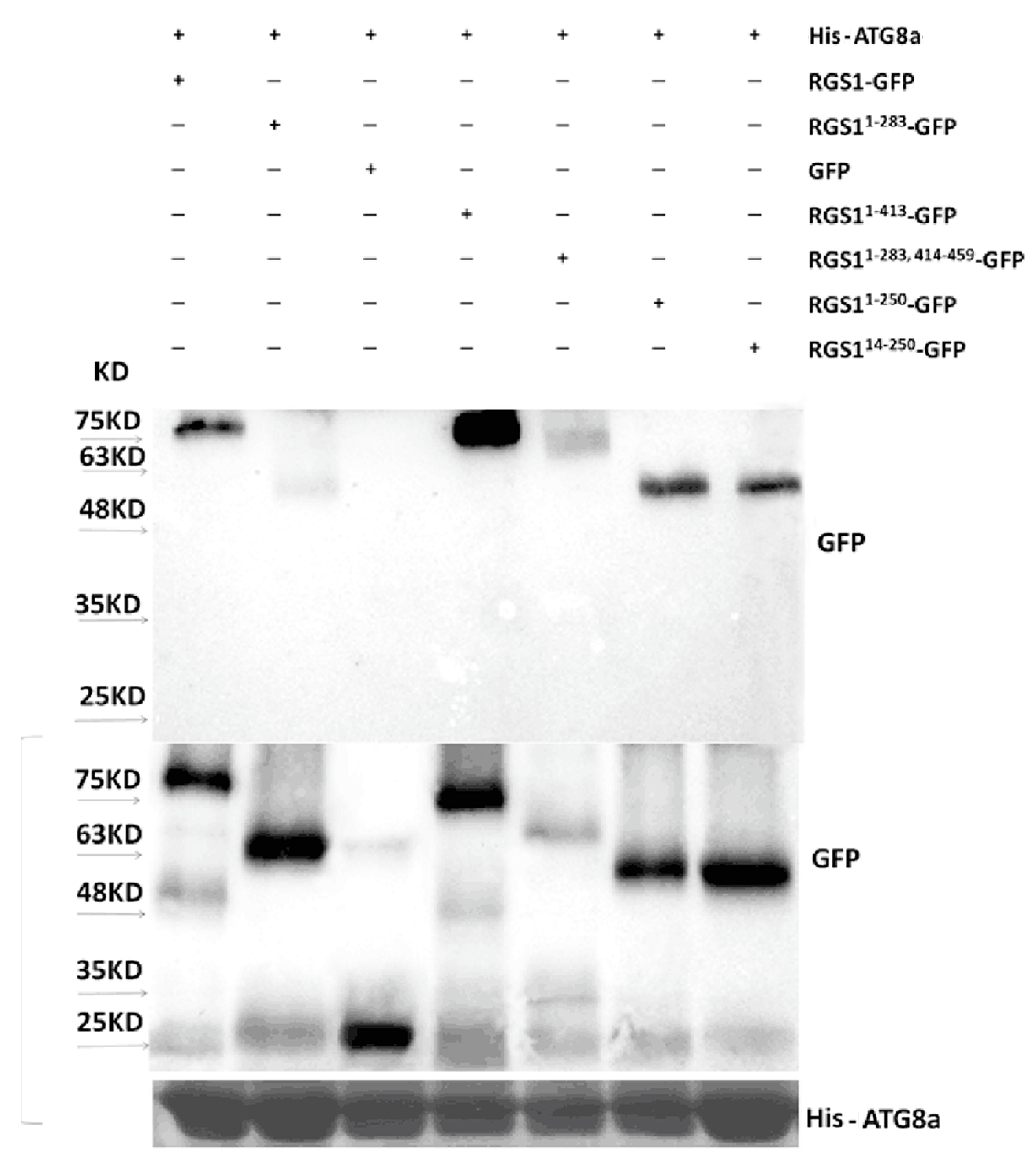
2.5. ATG8a Interacts with Full-Length and Truncated RGS1
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Arabidopsis Thaliana Mutants and Transgenic Lines
4.3. AtRGS1-YFP Internalization Analysis
4.4. BiFC
4.5. Transformation of BY-2 Cells
4.6. Co-Localization of RGS1 and Autophagosome
4.7. Pull Down Assays
4.8. Immunoblot Analyses
4.9. Confocal Microscopy
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Limanaqi, F.; Biagioni, F.; Busceti, C.L.; Ryskalin, L.; Soldani, P.; Frati, A.; Fornai, F. Cell Clearing Systems Bridging Neuro-Immunity and Synaptic Plasticity. Int. J. Mol. Sci. 2019, 20, 2197. [Google Scholar] [CrossRef] [PubMed]
- Farré, J.-C.; Subramani, S. Peroxisome turnover by micropexophagy: An autophagy-related process. Trends Cell Biol. 2004, 14, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Wang, J.; Fu, Z.Q.; Chen, W. Endocytosis of AtRGS1 Is Regulated by the Autophagy Pathway after D-Glucose Stimulation. Front. Plant Sci. 2017, 8, 1229. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Jäättelä, M. Lysosomes and autophagy in cell death control. Nat. Rev. Cancer 2005, 5, 886. [Google Scholar] [CrossRef] [PubMed]
- Bizargity, P.; Schröppel, B. Autophagy: Basic Principles and Relevance to Transplant Immunity: Autophagy and Transplant Immunity. Am. J. Transplant. 2014, 14, 1731–1739. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069. [Google Scholar] [CrossRef] [PubMed]
- Mehrpour, M.; Esclatine, A.; Beau, I.; Codogno, P. Overview of macroautophagy regulation in mammalian cells. Cell Res. 2010, 20, 748. [Google Scholar] [CrossRef] [PubMed]
- Bassham, D.C. Plant autophagy—more than a starvation response. Curr. Opin. Plant Biol. 2007, 10, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. An overview of the molecular mechanism of autophagy. In Autophagy in infection and immunity; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–32. [Google Scholar]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. Microautophagy in mammalian cells: Revisiting a 40-year-old conundrum. Autophagy 2011, 7, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Orenstein, S.J.; Cuervo, A.M. Chaperone-mediated autophagy: Molecular mechanisms and physiological relevance. In Proceedings of the Seminars in cell & developmental biology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 719–726. [Google Scholar]
- Reumann, S.; Voitsekhovskaja, O.; Lillo, C. From signal transduction to autophagy of plant cell organelles: Lessons from yeast and mammals and plant-specific features. Protoplasma 2010, 247, 233–256. [Google Scholar] [CrossRef] [PubMed]
- Bárány, I.; Berenguer, E.; Solís, M.-T.; Pérez-Pérez, Y.; Santamaría, M.E.; Crespo, J.L.; Risueño, M.C.; Díaz, I.; Testillano, P.S. Autophagy is activated and involved in cell death with participation of cathepsins during stress-induced microspore embryogenesis in barley. J. Exp. Bot. 2018, 69, 1387–1402. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Moore, B.; Sheen, J. Sugar sensing and signaling in plants. The plant cell 2002, 14, S185–S205. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Winderickx, J.; Thevelein, J.M. Glucose-sensing mechanisms in eukaryotic cells. Trends Biochem. Sci. 2001, 26, 310–317. [Google Scholar] [CrossRef]
- Huang, J.-P.; Tunc-Ozdemir, M.; Chang, Y.; Jones, A.M. Cooperative control between AtRGS1 and AtHXK1 in a WD40-repeat protein pathway in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 851. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.A.; Taylor, J.P.; Gao, Y.; Kimple, A.J.; Grigston, J.C.; Chen, J.-G.; Siderovski, D.P.; Jones, A.M.; Willard, F.S. GTPase acceleration as the rate-limiting step in Arabidopsis G protein-coupled sugar signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 17317–17322. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C.; Duffy, J.W.; Machius, M.; Temple, B.R.; Dohlman, H.G.; Jones, A.M. The crystal structure of a self-activating G protein α subunit reveals its distinct mechanism of signal initiation. Sci. Signal 2011, 4, ra8. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-G.; Willard, F.S.; Huang, J.; Liang, J.; Chasse, S.A.; Jones, A.M.; Siderovski, D.P. A seven-transmembrane RGS protein that modulates plant cell proliferation. Science 2003, 301, 1728–1731. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-G.; Jones, A.M. AtRGS1 function in Arabidopsis thaliana. In Methods in enzymology; Elsevier: Amsterdam, The Netherlands, 2004; pp. 338–350. [Google Scholar]
- Urano, D.; Chen, J.-G.; Botella, J.R.; Jones, A.M. Heterotrimeric G protein signalling in the plant kingdom. Open Biol. 2013, 3, 120186. [Google Scholar] [CrossRef]
- Zhong, C.-L.; Zhang, C.; Liu, J.-Z. Heterotrimeric G protein signaling in plant immunity. J. Exp. Bot. 2018, 70, 1109–1118. [Google Scholar] [CrossRef]
- Thompson, A.R.; Doelling, J.H.; Suttangkakul, A.; Vierstra, R.D. Autophagic nutrient recycling in Arabidopsis directed by the ATG8 and ATG12 conjugation pathways. Plant Physiol. 2005, 138, 2097–2110. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Baehrecke, E.H.; Brumell, J.H.; Chu, C.T.; Codogno, P.; Cuervo, A.M.; Debnath, J.; Deretic, V.; Elazar, Z.; Eskelinen, E.-L.; et al. A comprehensive glossary of autophagy-related molecules and processes. Autophagy 2011, 7, 1273–1294. [Google Scholar] [CrossRef]
- Kim, J.; Lee, H.; Lee, H.N.; Kim, S.-H.; Shin, K.D.; Chung, T. Autophagy-related proteins are required for degradation of peroxisomes in Arabidopsis hypocotyls during seedling growth. Plant Cell 2013, 25, 4956–4966. [Google Scholar] [CrossRef]
- Shin, K.D.; Lee, H.N.; Chung, T. A revised assay for monitoring autophagic flux in Arabidopsis thaliana reveals involvement of AUTOPHAGY-RELATED9 in autophagy. Mol. Cells 2014, 37, 399. [Google Scholar] [CrossRef]
- Meijer, A.J. Autophagy research: Lessons from metabolism. Taylor & Francis 2009, 5, 3–5. [Google Scholar]
- Wang, X.; Gao, Y.; Yan, Q.; Chen, W. Salicylic acid promotes autophagy via NPR3 and NPR4 in Arabidopsis senescence and innate immune response. Acta Physiol. Plant 2016, 38, 241. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, X.; Ma, C.; Chen, W. EDS1-mediated activation of autophagy regulates Pst DC3000 (AvrRps4)-induced programmed cell death in Arabidopsis. Acta Physiol. Plant. 2016, 38, 150. [Google Scholar] [CrossRef]
- Klionsky, D.J. For the last time, it is GFP-Atg8, not Atg8-GFP (and the same goes for LC3). Taylor & Francis 2011, 7, 1093–1094. [Google Scholar]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef]
- Agholme, L.; Hallbeck, M.; Benedikz, E.; Marcusson, J.; Kagedal, K. Amyloid-β secretion, generation, and lysosomal sequestration in response to proteasome inhibition: Involvement of autophagy. J. Alzheimer’s Dis. 2012, 31, 343–358. [Google Scholar] [CrossRef]
- Watanabe-Asano, T.; Kuma, A.; Mizushima, N. Cycloheximide inhibits starvation-induced autophagy through mTORC1 activation. Biochem. and Biophys. Res. Commun. 2014, 445, 334–339. [Google Scholar]
- Nocarova, E.; Fischer, L. Cloning of transgenic tobacco BY-2 cells; an efficient method to analyse and reduce high natural heterogeneity of transgene expression. BMC Plant Biol. 2009, 9, 44. [Google Scholar] [CrossRef]
- Hanamata, S.; Kurusu, T.; Okada, M.; Suda, A.; Kawamura, K.; Tsukada, E.; Kuchitsu, K. In vivo imaging and quantitative monitoring of autophagic flux in tobacco BY-2 cells. Plant Signaling Behav. 2013, 8, e22510. [Google Scholar] [CrossRef]
- Munafó, D.B.; Colombo, M.I. A novel assay to study autophagy: Regulation of autophagosome vacuole size by amino acid deprivation. J. Cell Sci. 2001, 114, 3619–3629. [Google Scholar]
- Liu, Y.; Schiff, M.; Czymmek, K.; Tallóczy, Z.; Levine, B.; Dinesh-Kumar, S. Autophagy regulates programmed cell death during the plant innate immune response. Cell 2005, 121, 567–577. [Google Scholar] [CrossRef]
- Dong, J.; Chen, W. The role of autophagy in chloroplast degradation and chlorophagy in immune defenses during Pst DC3000 (AvrRps4) infection. PLoS ONE 2013, 8, e73091. [Google Scholar] [CrossRef]
- Yano, K.; Matsui, S.; Tsuchiya, T.; Maeshima, M.; Kutsuna, N.; Hasezawa, S.; Moriyasu, Y. Contribution of the plasma membrane and central vacuole in the formation of autolysosomes in cultured tobacco cells. Plant Cell Physiol. 2004, 45, 951–957. [Google Scholar] [CrossRef]
- Journo, D.; Winter, G.; Abeliovich, H. Monitoring Autophagy in Yeast using FM 4–64 Fluorescence. Methods Enzymol. 2008, 451, 79–88. [Google Scholar]
- Oh-ye, Y.; Inoue, Y.; Moriyasu, Y. Detecting autophagy in Arabidopsis roots by membrane-permeable cysteine protease inhibitor E-64d and endocytosis tracer FM4–64. Plant Signaling Behav. 2011, 6, 1946–1949. [Google Scholar] [CrossRef][Green Version]
- Wang, Y.; Nishimura, M.T.; Zhao, T.; Tang, D. ATG2, an autophagy-related protein, negatively affects powdery mildew resistance and mildew-induced cell death in Arabidopsis. Plant J. 2011, 68, 74–87. [Google Scholar] [CrossRef]
- De Vries, L.; Fischer, T.; Tronchere, H.; Brothers, G.M.; Strockbine, B.; Siderovski, D.P.; Farquhar, M.G. Activator of G protein signaling 3 is a guanine dissociation inhibitor for Gαi subunits. Proc. Natl. Acad. Sci. USA 2000, 97, 14364–14369. [Google Scholar] [CrossRef]
- Zheng, Y.; Brockie, P.J.; Mellem, J.E.; Madsen, D.M.; Maricq, A.V. Neuronal control of locomotion in C. elegans is modified by a dominant mutation in the GLR-1 ionotropic glutamate receptor. Neuron 1999, 24, 347–361. [Google Scholar] [CrossRef]
- Lee, J.-K.; Kannarkat, G.T.; Chung, J.; Lee, H.J.; Graham, K.L.; Tansey, M.G. RGS10 deficiency ameliorates the severity of disease in experimental autoimmune encephalomyelitis. J. Neuroinflamm. 2016, 13, 24. [Google Scholar] [CrossRef]
- Xiong, Y.; Sheen, J. Rapamycin and glucose-target of rapamycin (TOR) protein signaling in plants. J. Biol. Chem. 2012, 287, 2836–2842. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Urano, D.; Phan, N.; Jones, J.C.; Yang, J.; Huang, J.; Grigston, J.; Taylor, J.P.; Jones, A.M. Endocytosis of the seven-transmembrane RGS1 protein activates G-protein-coupled signalling in Arabidopsis. Nat. Cell Biol. 2012, 14, 1079. [Google Scholar] [CrossRef]
- Fu, Y.; Lim, S.; Urano, D.; Tunc-Ozdemir, M.; Phan, N.G.; Elston, T.C.; Jones, A.M. Reciprocal encoding of signal intensity and duration in a glucose-sensing circuit. Cell 2014, 156, 1084–1095. [Google Scholar] [CrossRef]
- Klopffleisch, K.; Phan, N.; Augustin, K.; Bayne, R.S.; Booker, K.S.; Botella, J.R.; Carpita, N.C.; Carr, T.; Chen, J.-G.; Cooke, T.R.; et al. Arabidopsis G-protein interactome reveals connections to cell wall carbohydrates and morphogenesis. Mol. Syst. Biol. 2011, 7, 532. [Google Scholar] [CrossRef]
- Grigston, J.C.; Osuna, D.; Scheible, W.-R.; Liu, C.; Stitt, M.; Jones, A.M. D-glucose sensing by a plasma membrane regulator of G signaling protein, AtRGS1. FEBS Lett. 2008, 582, 3577–3584. [Google Scholar] [CrossRef]
- Nagata, T.; Nemoto, Y.; Hasezawa, S. Tobacco BY-2 cell line as the “HeLa” cell in the cell biology of higher plants. In International Review of Cytology; Elsevier: Amsterdam, The Netherlands, 1992; Volume 132, pp. 1–30. [Google Scholar]
- Hu, G.; Suo, Y.; Huang, J. A crucial role of the RGS domain in trans-Golgi network export of AtRGS1 in the protein secretory pathway. Mol. Plant 2013, 6, 1933–1944. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Y.; Tang, S.; Zhao, Q.; Zhang, Z.; Zhang, H.; Dong, L.; Guo, H.; Xie, Q. An efficient system to detect protein ubiquitination by agroinfiltration in Nicotiana benthamiana. Plant J. 2010, 61, 893–903. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, Y.; Srba, M.; Wang, J.; Chen, W. Correlation of Autophagosome Formation with Degradation and Endocytosis Arabidopsis Regulator of G-Protein Signaling (RGS1) through ATG8a. Int. J. Mol. Sci. 2019, 20, 4190. https://doi.org/10.3390/ijms20174190
Jiao Y, Srba M, Wang J, Chen W. Correlation of Autophagosome Formation with Degradation and Endocytosis Arabidopsis Regulator of G-Protein Signaling (RGS1) through ATG8a. International Journal of Molecular Sciences. 2019; 20(17):4190. https://doi.org/10.3390/ijms20174190
Chicago/Turabian StyleJiao, Yue, Miroslav Srba, Jingchun Wang, and Wenli Chen. 2019. "Correlation of Autophagosome Formation with Degradation and Endocytosis Arabidopsis Regulator of G-Protein Signaling (RGS1) through ATG8a" International Journal of Molecular Sciences 20, no. 17: 4190. https://doi.org/10.3390/ijms20174190
APA StyleJiao, Y., Srba, M., Wang, J., & Chen, W. (2019). Correlation of Autophagosome Formation with Degradation and Endocytosis Arabidopsis Regulator of G-Protein Signaling (RGS1) through ATG8a. International Journal of Molecular Sciences, 20(17), 4190. https://doi.org/10.3390/ijms20174190