Systematic Analysis of Differentially Expressed Maize ZmbZIP Genes between Drought and Rewatering Transcriptome Reveals bZIP Family Members Involved in Abiotic Stress Responses

Abstract
:
1. Introduction
2. Results
2.1. Genome-Wide Identification and Classification of bZIP Genes in Maize
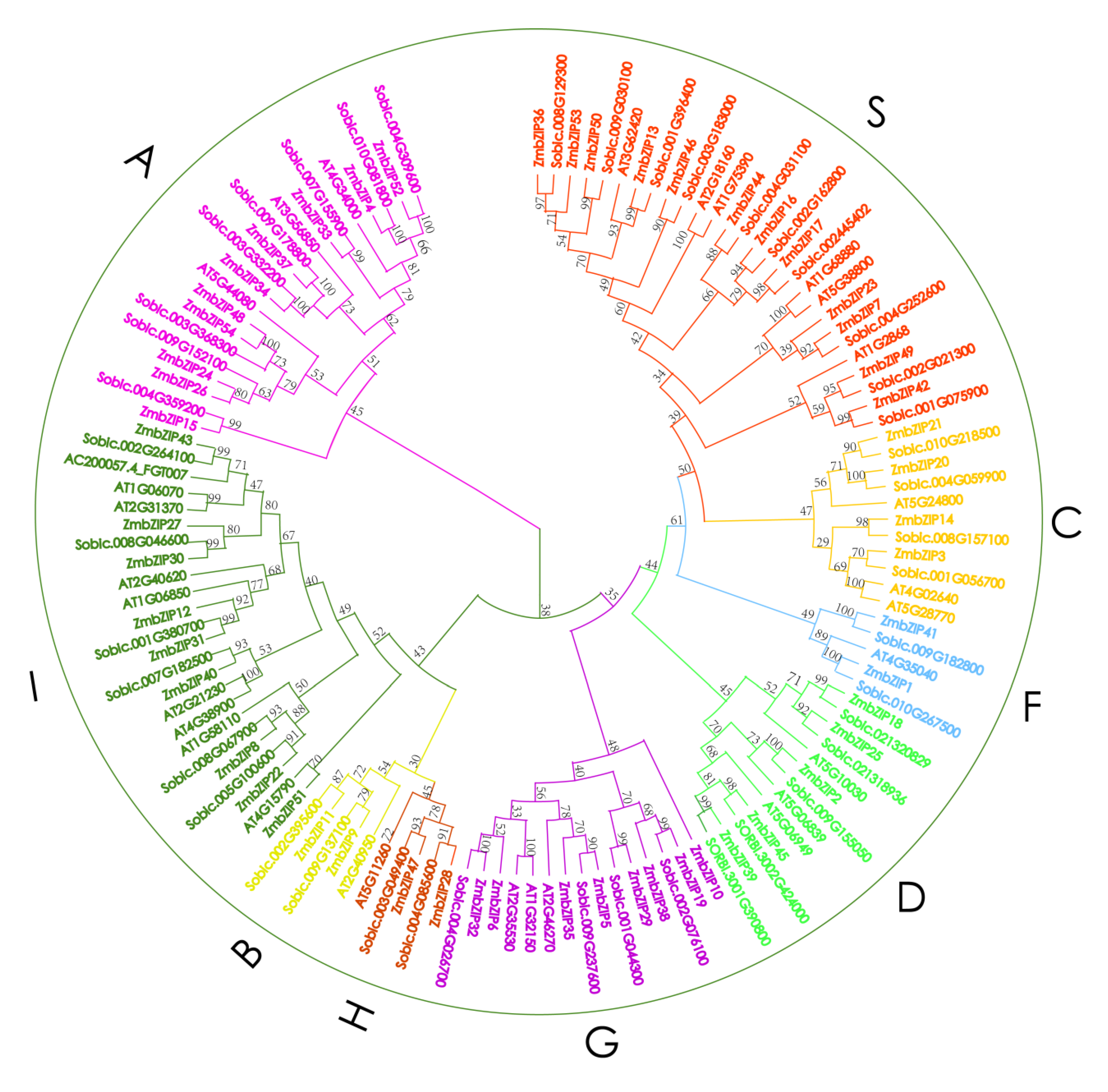
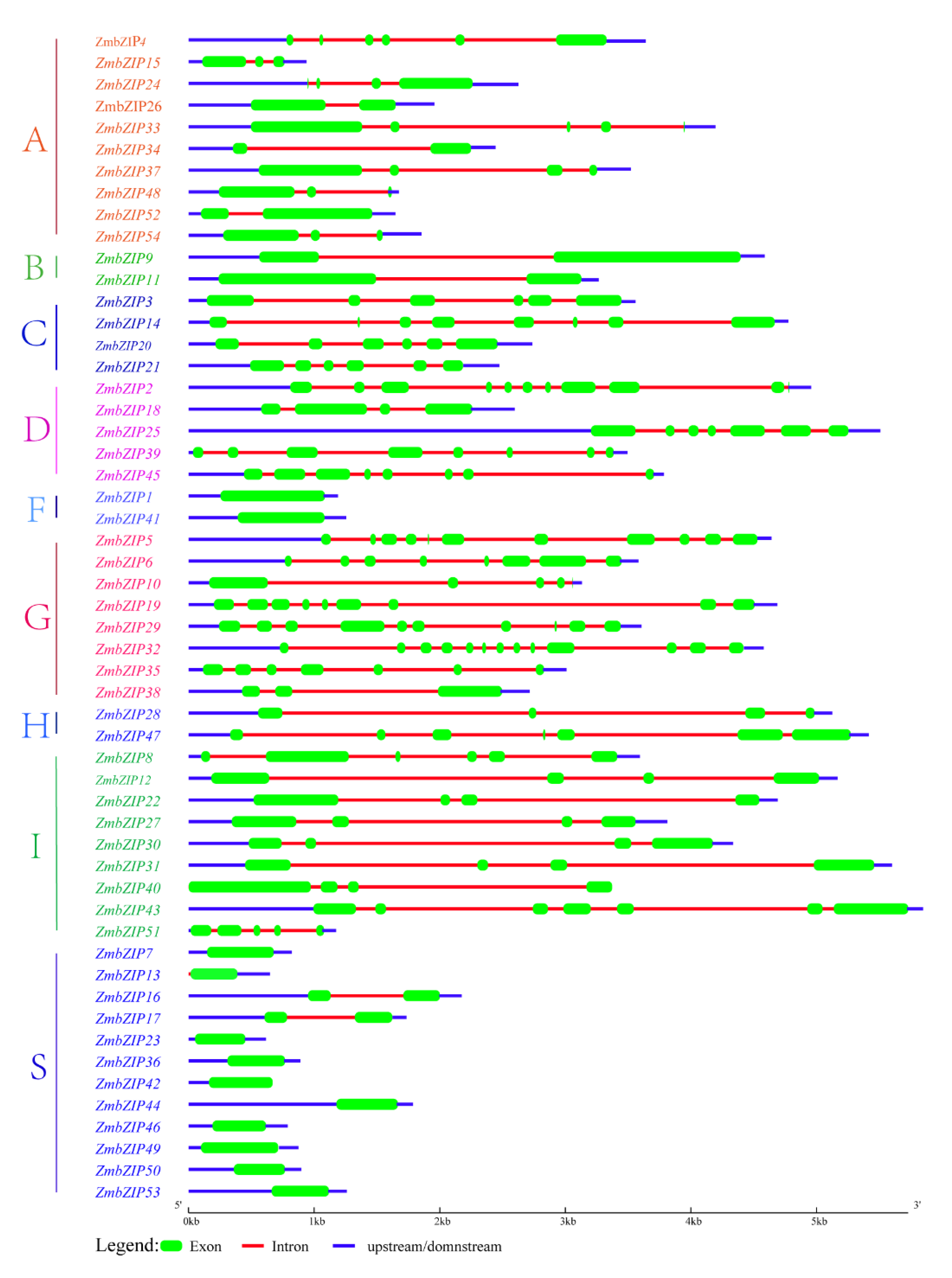
2.2. Structure of bZIP Genes in Maize
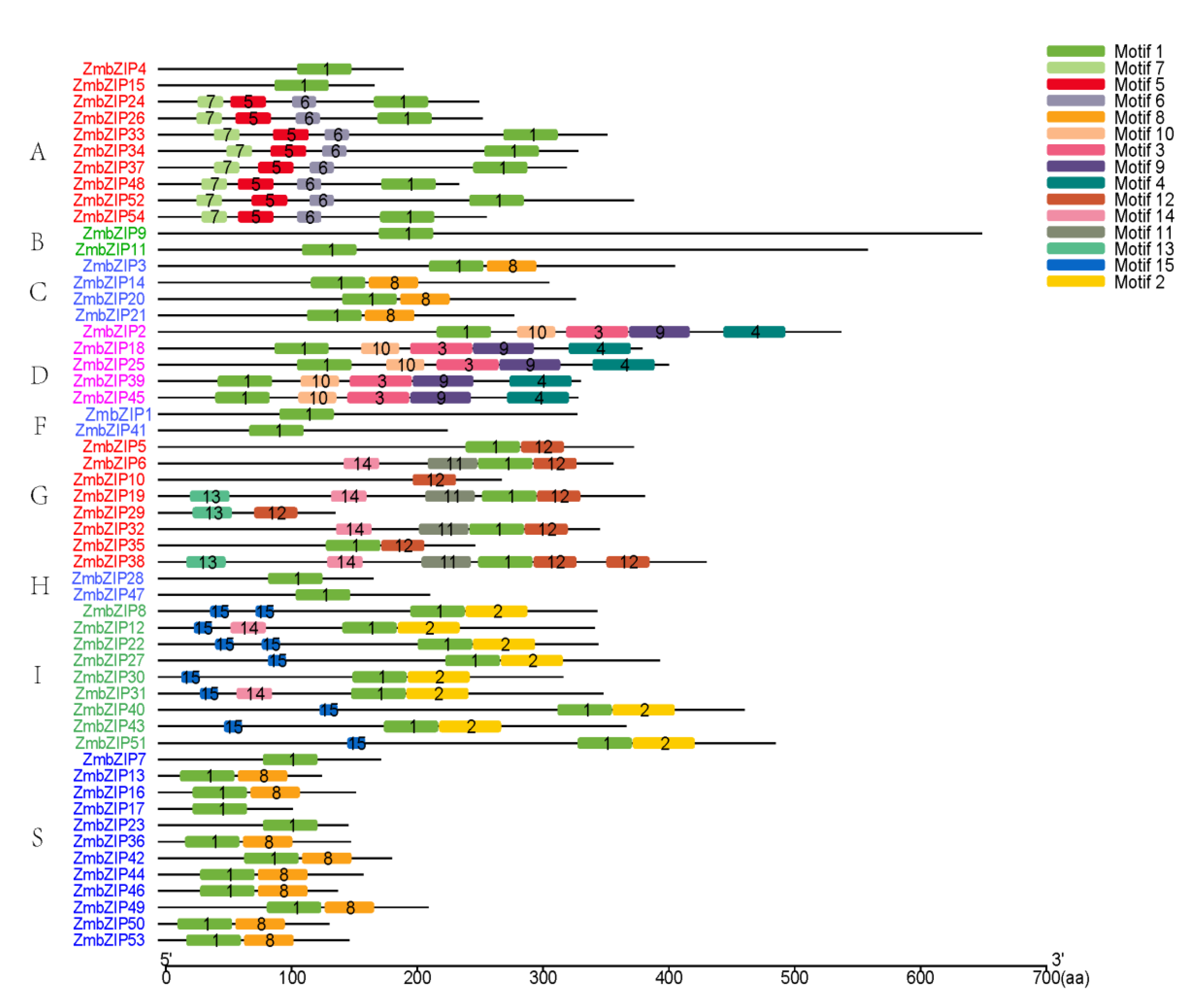
2.3. Additional Conserved Motifs in bZIP Genes in Maize
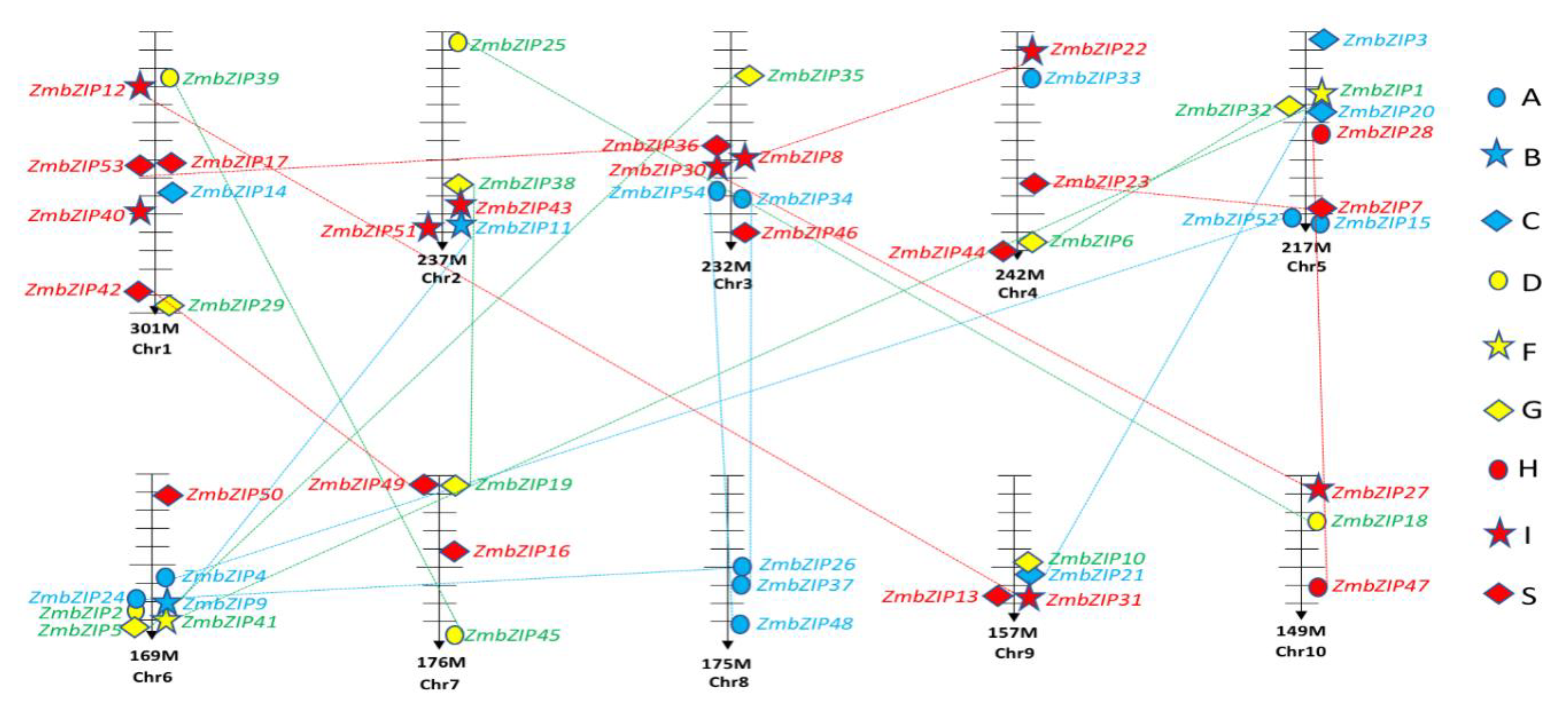
2.4. Chromosomal Locations and Duplications of ZmbZIPs
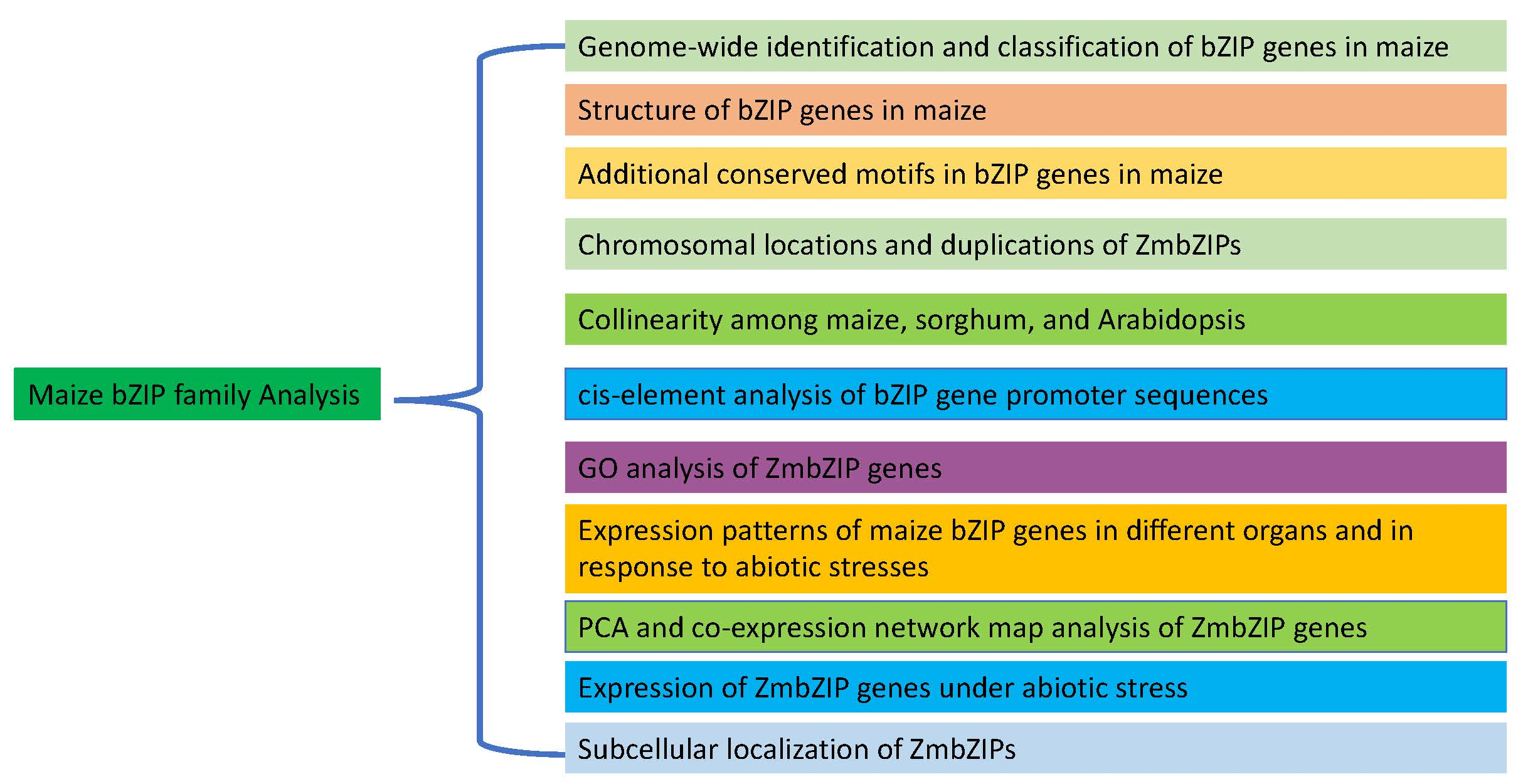
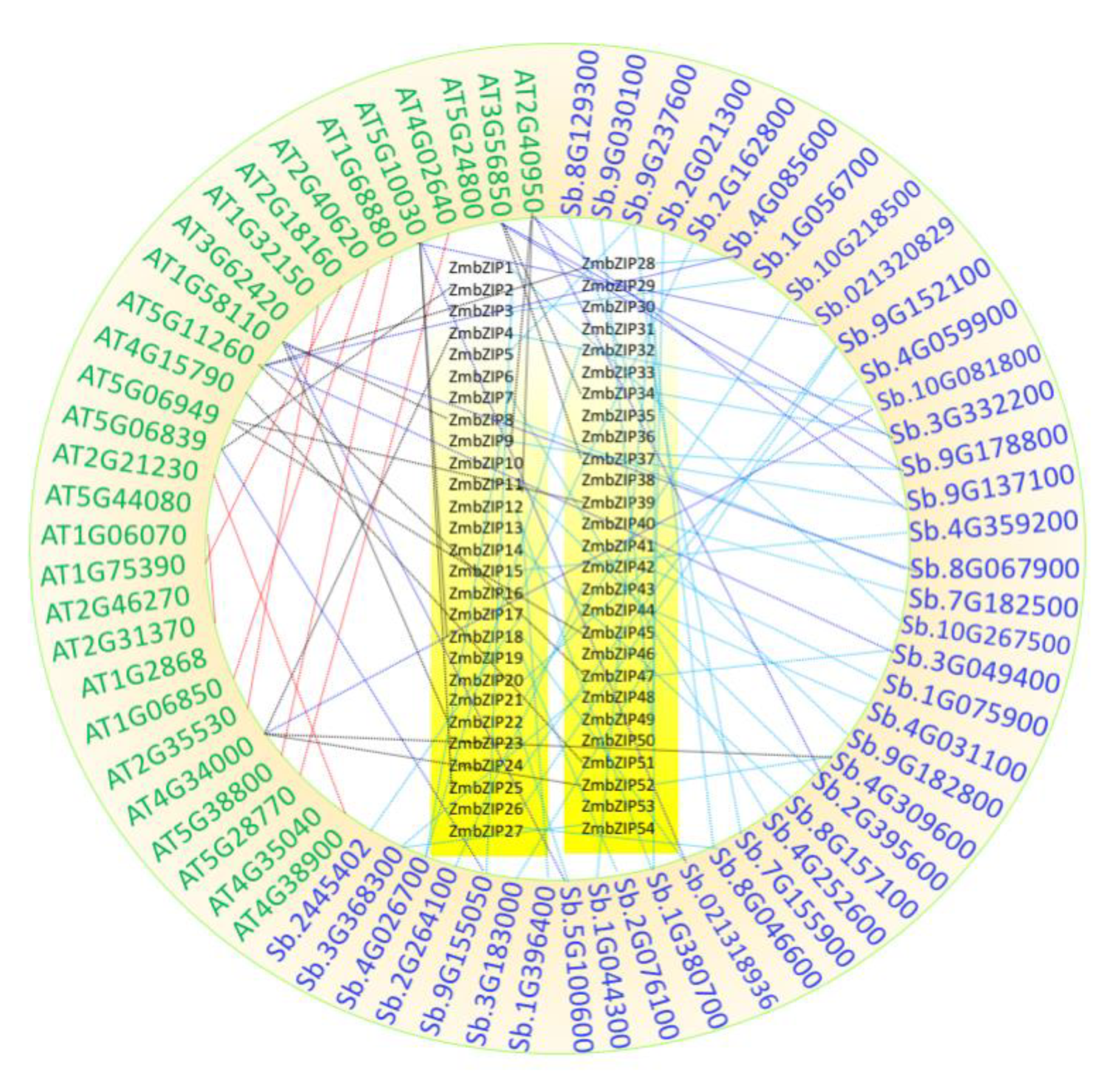
2.5. Collinearity Among Maize, Sorghum, and Arabidopsis
2.6. Cis-Element Analysis of bZIP Gene Promoter Sequences
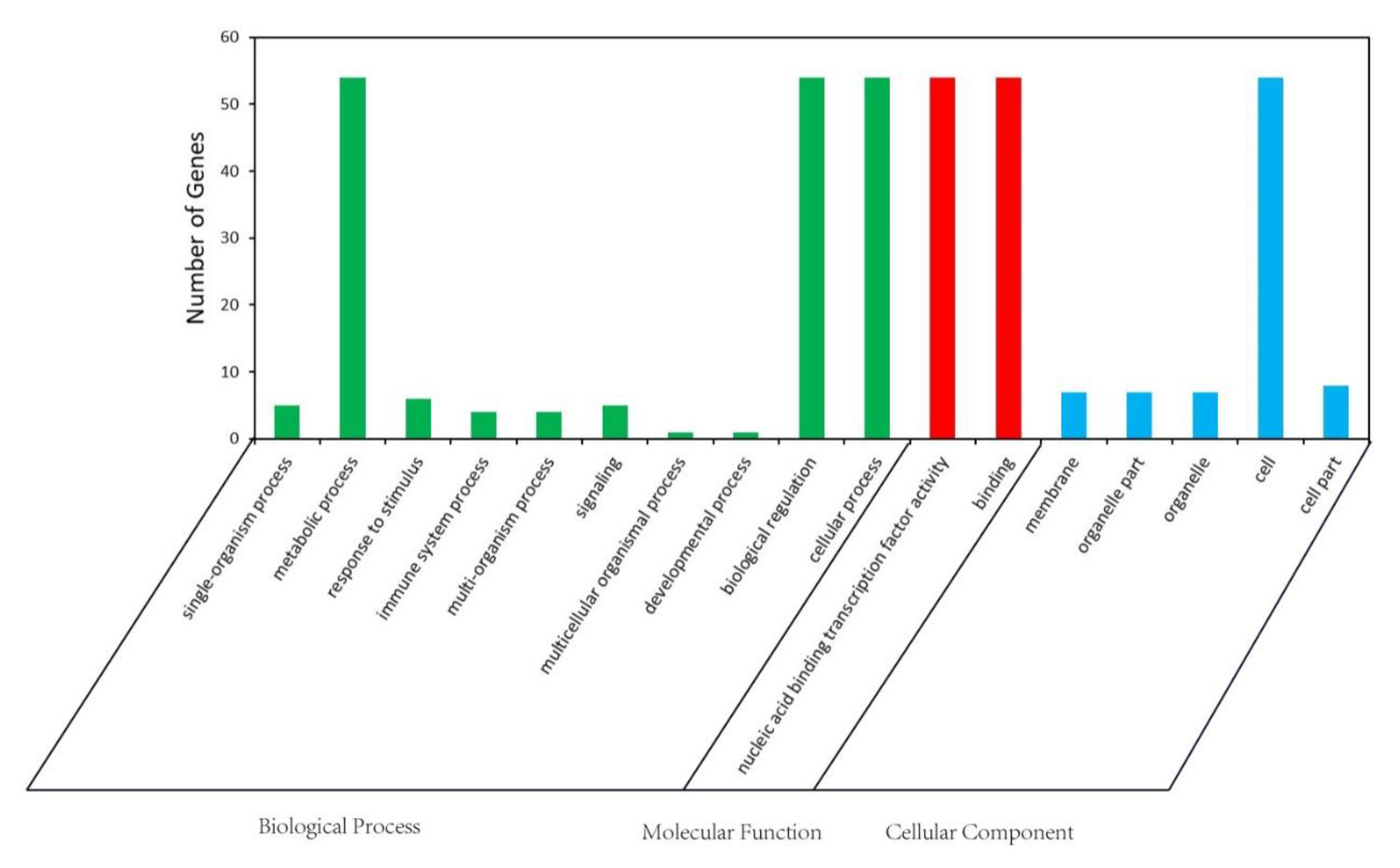
2.7. GO Analysis of ZmbZIP Genes
2.8. Expression Patterns of Maize bZIP Genes in Different Organs and in Response to Abiotic Stresses
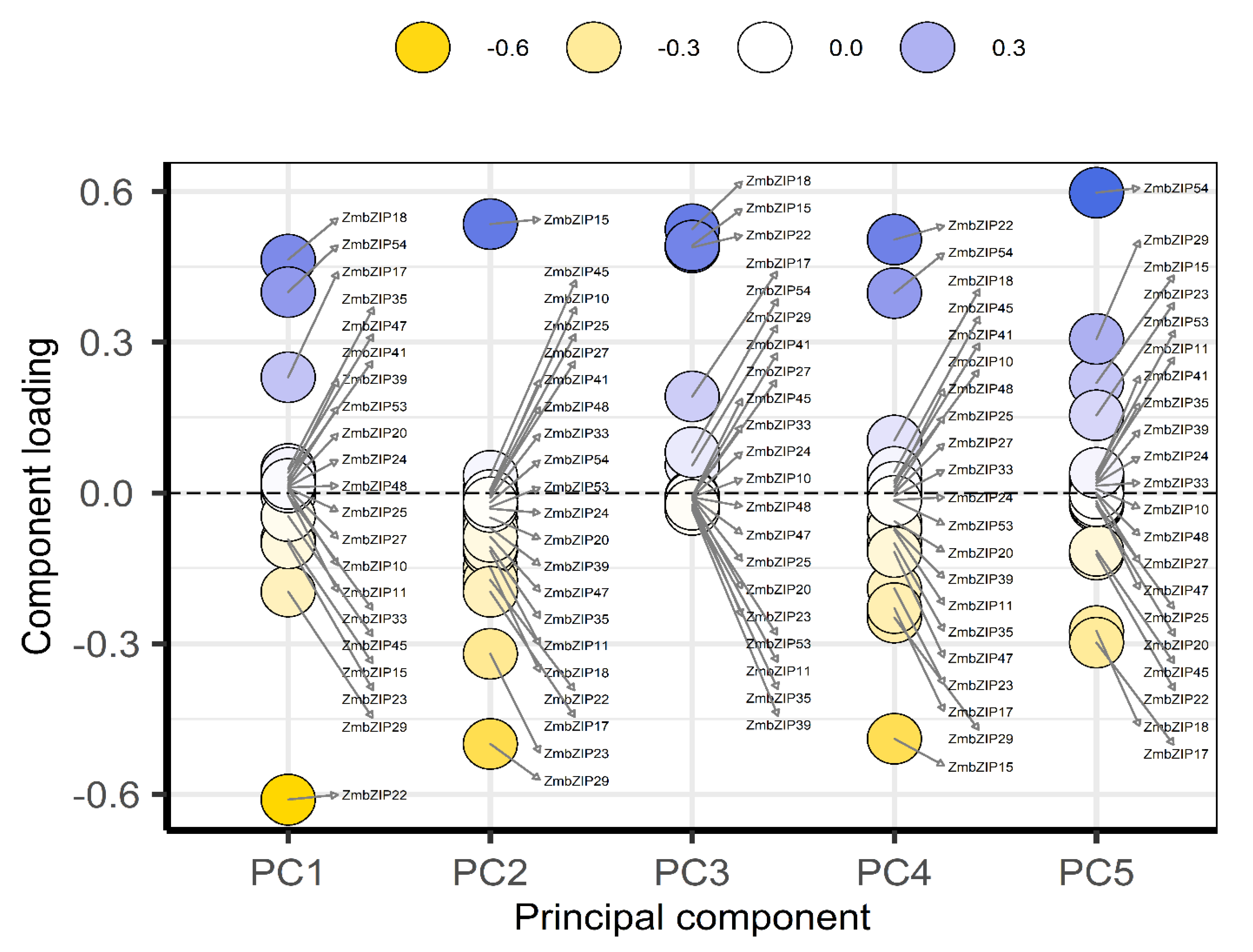
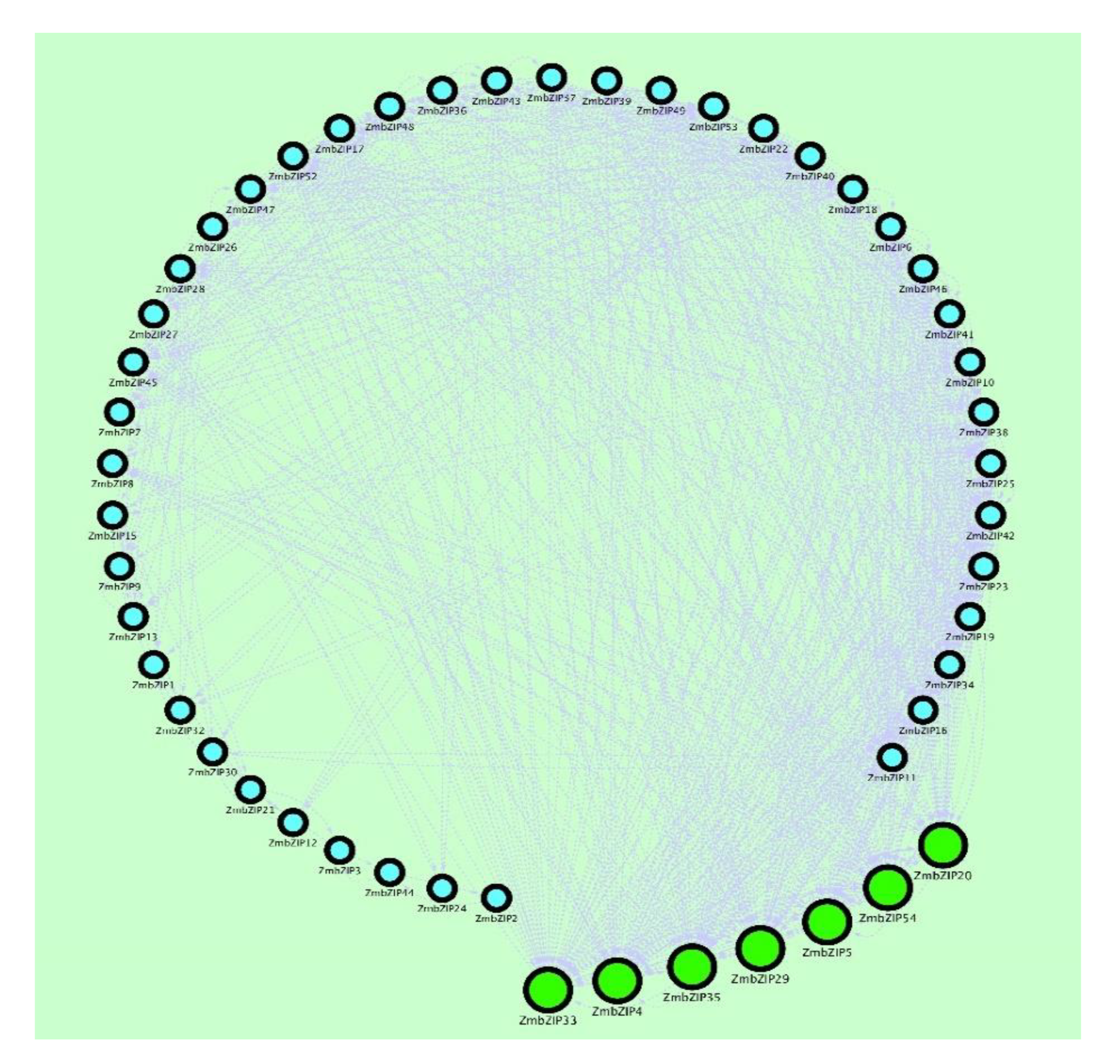
2.9. PCA and Co-Expression Network Map Analysis of ZmbZIP Genes
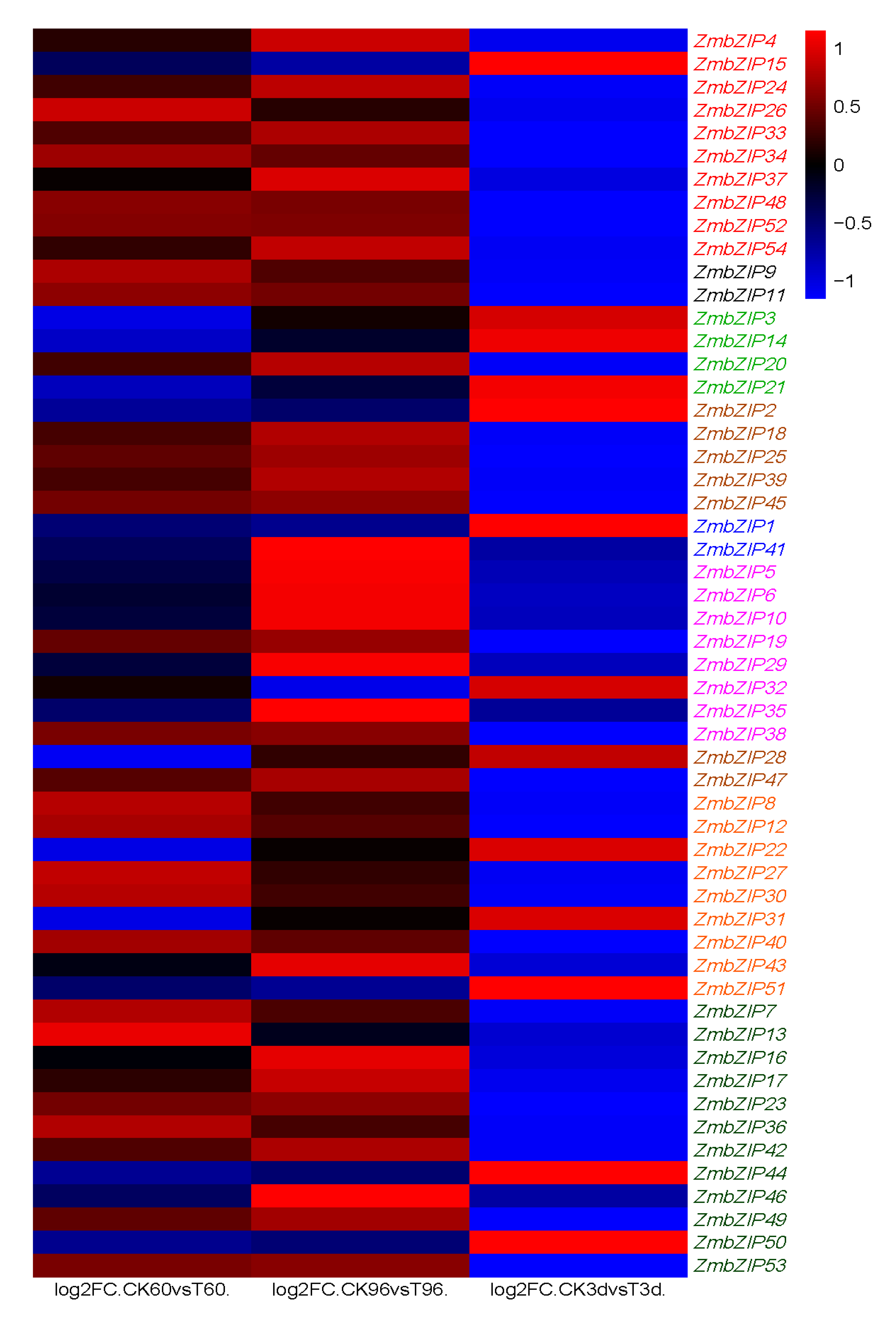
2.10. Expression of ZmbZIP Genes under Abiotic Stress
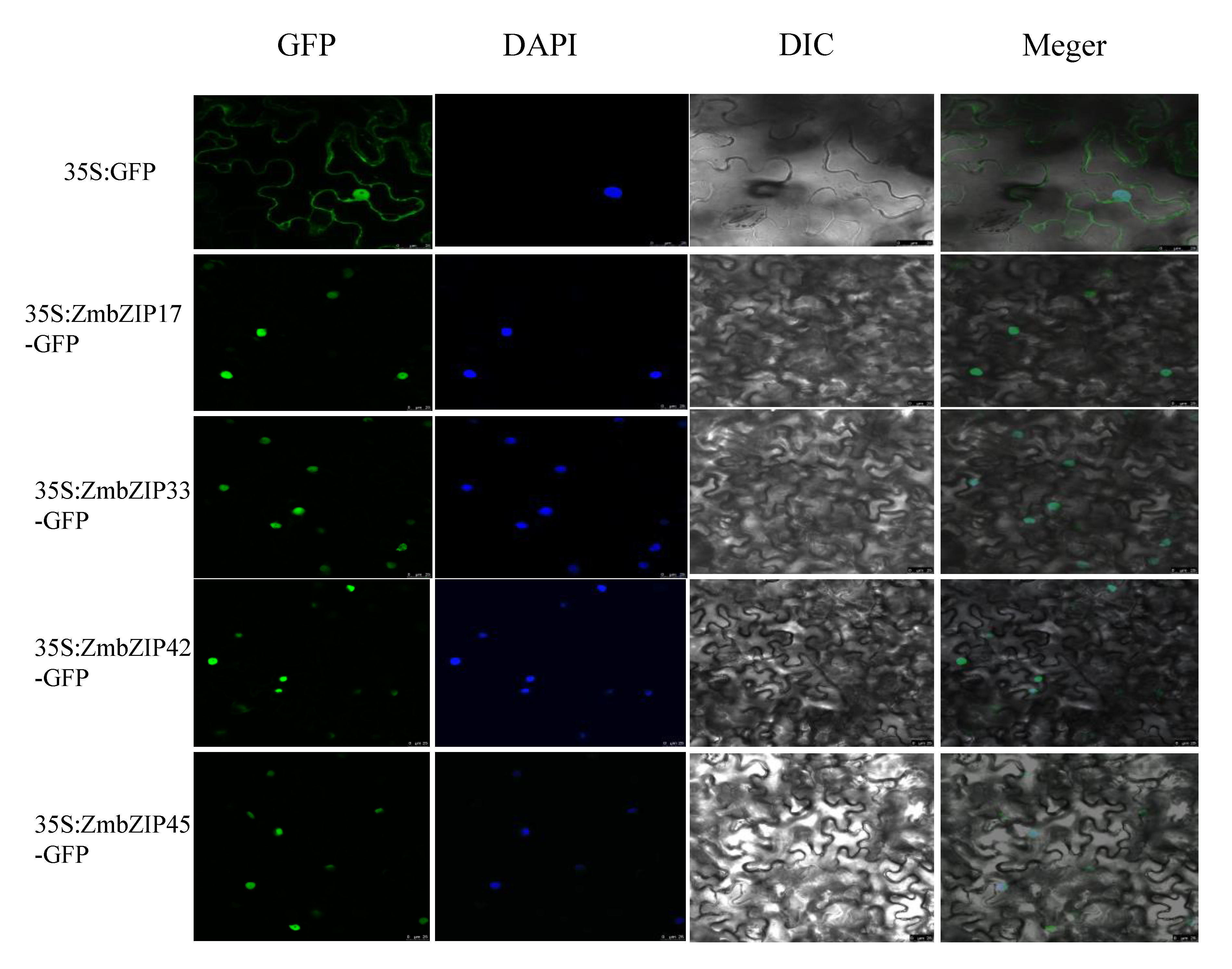
2.11. Subcellular Localization of ZmbZIPs
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatment
4.2. RNA Isolation and qRT-PCR Analysis
4.3. Collection and Classification of bZIP Transcription Factors
4.4. Compound Phylogenetic Tree, Gene Structure, Additional Conserved Motifs Analysis, and Chromosomal Locations
4.5. Interspecies Microsynteny Analysis
4.6. Cis-Elements in the Promoter Regions of Abiotic Stress-Responsive and Microarray-Based Expression Analysis of ZmbZIP Genes
5.7. The PCA and Co-Expression Network Map Analysis of ZmbZIP Genes
5.8. Determination of Subcellular Localization of ZmbZIPs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, D.J.; Yu, G. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Broun, P. Transcription factors as tools for metabolic engineering in plants. Curr. Opin. Plant Biol. 2004, 7, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchishinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic survey and gene expression analysis of the basic leucine zipper transcription factor family in rice. Plant Physiol. 2008, 146, 333–350. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Nishiyama, M.Y.; Fuentes, B.G.; Souza, G.M.; Janies, D.; Gray, J.; Grotewold, E. GRASSIUS: A platform for comparative regulatory genomics across the grasses. Plant Physiol. 2009, 149, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, B.M.; Vahap, E.; Mortaza, H.; Turgay, U.; Zhang, B.H. Genome-wide analysis of the bzip transcription factors in cucumber. PLoS ONE 2014, 9, e96014. [Google Scholar]
- Meng, D. Genome-wide identification and expression analysis of the bzip gene family in apple (malus domestica). Tree Genet. Genomes 2016, 12, 82. [Google Scholar]
- Wang, X.L.; Chen, X.; Yang, T.B.; Cheng, Q.; Cheng, Z.M. Genome-wide identification of bzip family genes involved in drought and heat stresses in strawberry (fragaria vesca). Int. J. Genom. 2017, 1–14. [Google Scholar] [CrossRef]
- Kim, S.; Kang, J.Y.; Cho, D.I.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef]
- Schütze, K.; Harter, K.; Chaban, C. Post-translational regulation of plant bzip factors. Cell 2008, 13, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Sibéril, Y.; Doireau, P.; Gantet, P. Plant bZIP G-box binding factors. Modular structure and activation mechanisms. Eur. J. Biochem. 2001, 268, 5655–5666. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Yun, K.Y.; Ressom, H.; Mohanty, B.; Bajic, V.B.; Jia, Y.; Yun, S.J.; De los Reyes, B.G. An early response regulatory cluster induced by low temperature and hydrogen peroxide in seedlings of chilling-tolerant japonica rice. BMC Genom. 2007, 8, 175. [Google Scholar] [CrossRef]
- Correa, L.G.; Riano-Pachon, D.M.; Schrago, C.G.; Mueller-Roeber, B.; Vincents, M. The role of bZIP transcription factors in green plant evolution: Adaptive features emerging from four founder genes. PLoS ONE 2008, 8, e2994. [Google Scholar] [CrossRef]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef]
- Silveira, A.B.; Gauer, L.; Tomaz, J.P.; Cardoso, P.R.; Carmello-Guerreiro, S.; Vincentz, M. The Arabidopsis AtbZIP9 protein fused to the VP16 transcriptional activation domain alters leaf and vascular development. Plant Sci. 2007, 172, 1148–1156. [Google Scholar] [CrossRef]
- Fukazawa, J.; Sakai, T.; Ishida, S.; Yamaguchi, I.; Kamiya, Y.; Takahashi, Y. Repression of shoot growth, a bZIP transcriptional activator, regulates cell elongation by controlling the level of gibberellins. Plant Cell 2000, 12, 901–915. [Google Scholar] [CrossRef]
- Guan, Y.; Ren, H.; Xie, H.; Ma, Z.; Chen, F. Identification and characterization of bZIP-type transcription factors involved in carrot (Daucus carota L.) somatic embryogenesis. Plant J. 2009, 60, 207–217. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Lynch, T.J. Abscisic acid inhibition of radicle emergence but not seedling growth is suppressed by sugars. Plant Physiol. 2000, 122, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [PubMed]
- Nieva, C.; Busk, P.K.; Domínguez-Puigjaner, E.; Lumbreras, V.; Testillano, P.S.; Risueño, M.C.; Pagès, M. Isolation and functional characterisation of two new bZIP maize regulators of the ABA responsive generab28. Plant Mol. Biol. 2005, 58, 899–914. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.G.; Price, J.; Lin, P.C.; Hong, J.C.; Jang, J.C. The Arabidopsis bZIP1 transcription factor is involved in sugar signaling, protein networking, and DNA binding. Mol. Plant. 2010, 3, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Ulm, R.; Baumann, A.; Oravecz, A.; Mate, Z.; Adam, E.; Oakeley, E.J.; Schafer, E.; Nagy, F. Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 1397–1402. [Google Scholar] [CrossRef] [PubMed]
- Weltmeier, F.; Ehlert, A.; Mayer, C.S.; Dietrich, K.; Wang, X.; Schutze, K.; Alonso, R.; Harter, K.; Vicente-Carbajosa, J.; Droge-Laser, W. Combinatorial control of Arabidopsis proline dehydrogenase transcription by specific heterodimerisation of bZIP transcription factors. EMBO J. 2006, 25, 3133–3143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.X.; Srivastava, R.; Howell, S.H. Stress-induced expression of an activated form of AtbZIP17 provides protection from salt stress in Arabidopsis. Plant Cell Environ. 2008, 31, 1735–1743. [Google Scholar] [CrossRef]
- Weltmeier, F.; Rahmani, F.; Ehlert, A.; Dietrich, K.; Schutze, K.; Wang, X.; Chaban, C.; Hanson, J.; Teige, M.; Harter, K.; et al. Expression patterns within the Arabidopsis C/S1 bZIP transcription factor network: Availability of heterodimerization partners controls gene expression during stress response and development. Plant Mol. Biol. 2009, 69, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Yang, O.; Popova, O.V.; Suthoff, U.; Luking, I.; Dietz, K.J.; Golldack, D. The Arabidopsis basic leucine zipper transcription factor AtbZIP24 regulates complex transcriptional networks involved in abiotic stress resistance. Gene 2009, 436, 45–55. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2 and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef]
- Tang, W.; Page, M.; Fei, Y.; Liu, L.; Xu, F.; Cai, X. Overexpression ofatbzip60deltacgene alleviates salt-induced oxidative damage in transgenic cell cultures. Plant Mol. Biol. Rep. 2012, 30, 1183–1195. [Google Scholar] [CrossRef]
- Mukherjee, K.; Choudhury, A.R.; Gupta, B.; Gupta, S.; Sengupta, D.N. An ABRE-binding factor, OSBZ8, is highly expressed in salt tolerant cultivars than in salt sensitive cultivars of indica rice. BMC Plant Biol. 2006, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Guan, Y.; Ren, H.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef]
- Yun, K.Y.; Park, M.R.; Mohanty, B.; Herath, V.; Xu, F.; Mauleon, R.; Wijaya, E.; Bajic, V.B.; Bruskiewich, R.; Reyes, B.G. Transcriptional regulatory network triggered by oxidative signals configures the early response mechanisms of japonica rice to chilling stress. BMC Plant Biol. 2010, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Citao, L.; Bigang, M.; Shujun, O.; Wang, W.; Liu, L.C.; Wu, Y.B.; Chu, C.C.; Wang, X.P. Correction to: Osbzip71, a bzip transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2018, 97, 467–468. [Google Scholar]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar, K.; Singla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (oshbp1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J. Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, F.; Maeta, E.; Terashima, A.; Takumi, S. Positive role of a wheat HvABI5 ortholog in abiotic stress response of seedlings. Physiol. Plant. 2008, 134, 74–86. [Google Scholar] [CrossRef]
- Liao, Y.; Zhang, J.S.; Chen, S.Y.; Zhang, W.K. Role of soybean GmbZIP132 under abscisic acid and salt stresses. J. Integr. Plant Biol. 2008a, 50, 221–230. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress. Protoplasma 2017, 254, 3–16. [Google Scholar] [CrossRef]
- Hsieh, T.H.; Li, C.W.; Su, R.C.; Cheng, C.P.; SanjayaTsai, Y.C.; Chan, M.T. A tomato bZIP transcription factor, SlAREB, is involved in water deficit and salt stress response. Planta 2010, 231, 1459–1473. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, Y.; Jiang, H.; Li, X.; Gan, D.; Peng, X.; Zhu, S.; Cheng, B. Systematic analysis of sequences and expression patterns of drought-responsive members of the HD-Zip gene family in maize. PLoS ONE 2011, 6, e28488. [Google Scholar] [CrossRef]
- Vanitha, J.; Ramachandran, S. Genome-wide Expansion and Expression Divergence of the Basic Leucine Zipper Transcription Factors in Higher Plants with an Emphasis on Sorghum. J. Integr. Plant Biol. 2011, 53, 212–231. [Google Scholar]
- Wei, K.F.; Wu, L.J.; Chen, J.; Chen, Y.F.; Xie, D.X. Structural evolution and functional diversification analyses of argonaute protein. J. Cell. Biochem. 2012, 113, 2576–2585. [Google Scholar] [CrossRef]
- Ji, L.; Wang, J.; Ye, M.; Li, Y.; Guo, B.; Chen, Z.; Li, H.; An, X. Identification and characterization of the Populus AREB/ABF subfamily. J. Integr. Plant Biol. 2013, 55, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujita, M.; Satoh, R.; Maruyama, K.; Parvez, M.M.; Seki, M.; Hiratsu, K.; OhmeTakagi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell 2005, 17, 3470–3488. [Google Scholar] [CrossRef]
- Ciccri, P.; Gianazza, E.B.; Lazzari, G. Phosphorylation of Opaque2 changes diurnally and impacts its DNA binding activity. Plant Cell 1997, 9, 97–108. [Google Scholar]
- Schwartz, S.; Meshorer, E.; Ast, G. Chromatin organization marks exon-intron structure. Nat. Struct. Mol. Biol. 2009, 16, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.Y.; Fu, X.L.; Tan, Q.P.; Liu, L.; Chen, M.; Zhu, C.Y. Analysis of basic leucine zipper genes and their expression during bud dormancy in peach (prunus persica). Plant Physiol. Biochem. 2016, 104, 54–70. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.X.; Jiang, H.Y.; Chu, Z.X.; Tang, X.L.; Zhu, S.W.; Cheng, B.J. Genome-wide identification, classification and analysis of heat shock transcription factor family in maize. BMC Genom. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4. [Google Scholar] [CrossRef] [PubMed]
- Yves, V.D.P.; Maere, S.; Meyer, A. The evolutionary significance of ancient genome duplications. Nat. Rev. Genet. 2009, 10, 725–732. [Google Scholar] [Green Version]
- Deppmann, C.D.; Alvania, R.S.; Taparowsky, E.J. Cross-species annotation of basic leucine zipper factor interactions: Insight into the evolution of closed interaction networks. Mol. Biol. Evol. 2006, 23, 1480–1492. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhu, H.H.; Zhou, L.F.; Li, J.; Zhao, L.Y.; Wu, S.S. Genes related to the very early stage of cona-induced fulminant hepatitis: A gene-chip-based study in a mouse model. BMC Genom. 2010, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Yang, Z.R.; Li, Z.; Zhang, F.L.; Hao, L. De novo transcriptome assembly and co-expression network analysis of Cynanchum thesioides: Identification of genes involved in resistance to drought stress. Gene 2019, 710, 375–386. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Guo, R.; Guo, C.; Hou, H.; Wang, X.; Gao, H. Evolutionary and expression analyses of the apple basic leucine zipper transcription factor family. Front. Plant Sci. 2016, 7, 376. [Google Scholar] [CrossRef]
- Gao, M.; Zhang, H.; Guoetal, C. Evolutionary and expression analyses of basic zipper transcription factors in the highly homozygous model grape PN40024 (Vitis vinifera L.). Plant Mol. Biol. Rep. 2014, 32, 1085–1102. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mizoi, J.; Todaka, D.; Shinozaki, K. Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. 2015, 38, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.S.; Dadalto, S.P.; Goncalves, A.B.; De Souza, G.B.; Barros, V.A.; Fietto, L.G. Plant bZIP transcription factors responsive to pathogens: A Review. Int. J. Mol. Sci. 2013, 14, 7815–7828. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, Y.; Cai, H.; Bai, X.; Ji, W.; Ding, X.; Zhu, Y. The Arabidopsis AtbZIP1 transcription factor is a positive regulator of plant tolerance to salt, osmotic and drought stresses. J. Plant Res. 2012, 125, 429–438. [Google Scholar] [CrossRef]
- Chung, E.; Seong, E.; Kim, Y.C.; Chung, E.J.; Oh, S.K.; Lee, S.; Park, J.M.; Joung, Y.H.; Choi, D. A method of high frequency virus induced gene silencing in Chili pepper (Capsicum annuum L. Bukang). Mol. Cells 2004, 17, 377–380. [Google Scholar] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | ID Gene Identifier | Gene Names Description | Protein Size (aa) | Chromsome | 5′ End | 3′ End | Molecular Weight (kDa) | Theoretical pI | Instability Index | GRAVY |
|---|---|---|---|---|---|---|---|---|---|---|
| ZmbZIP1 | GRMZM2G000171 | Basic leucine zipper 19 | 332 | Chr5 | 61724734 | 61728541 | 35.43 | 6.16 | 44.93 | −0.34 |
| ZmbZIP2 | GRMZM2G000842 | TGACG motif-binding transcription factor 6 | 542 | Chr6 | 149721944 | 149728050 | 60.34 | 8.6 | 61.64 | −0.451 |
| ZmbZIP3 | GRMZM2G007063 | Basic leucine zipper 25 | 410 | Chr5 | 4253958 | 4257719 | 42.88 | 5.23 | 55.72 | −0.467 |
| ZmbZIP4 | GRMZM2G008166 | ABA-INSENSITIVE 5 | 194 | Chr6 | 111882466 | 111885951 | 21.58 | 9.85 | 31.95 | −0.303 |
| ZmbZIP5 | GRMZM2G011932 | G-box binding factor 1 | 377 | Chr6 | 165543429 | 165548373 | 40.12 | 7.11 | 64.81 | −0.678 |
| ZmbZIP6 | GRMZM2G019106 | Transcription factor HBP-1a | 361 | Chr4 | 239534487 | 239538429 | 38.21 | 5.31 | 58.4 | −1.017 |
| ZmbZIP7 | GRMZM2G020799 | Basic leucine-zipper 58 | 176 | Chr5 | 204603643 | 204604321 | 20.27 | 6.52 | 78.7 | −0.882 |
| ZmbZIP8 | GRMZM2G024851 | DNA binding protein | 348 | Chr3 | 138906763 | 138910797 | 39.14 | 8.8 | 70.2 | −0.966 |
| ZmbZIP9 | GRMZM2G045236 | DNA binding protein | 654 | Chr6 | 147087480 | 147092015 | 69.66 | 9.03 | 43.59 | −0.49 |
| ZmbZIP10 | GRMZM2G050912 | Putative bZIP transcription factor | 272 | Chr9 | 93665910 | 93671589 | 28.53 | 8.94 | 74.35 | −0.616 |
| ZmbZIP11 | GRMZM2G060109 | Putative bZIP transcription factor | 563 | Chr2 | 214613200 | 214616521 | 59.82 | 5.48 | 53.55 | −0.37 |
| ZmbZIP12 | GRMZM2G062391 | Basic leucine-zipper 52 | 346 | Chr1 | 56217970 | 56223418 | 37.38 | 6.43 | 50.64 | −0.635 |
| ZmbZIP13 | GRMZM2G066734 | bZIP transcription factor 53 | 129 | Chr9 | 135645489 | 135646430 | 14.48 | 10.89 | 68.73 | −0.417 |
| ZmbZIP14 | GRMZM2G073427 | Basic leucine zipper 25 | 310 | Chr1 | 170592733 | 170597621 | 33 | 5.19 | 45.97 | −0.525 |
| ZmbZIP15 | GRMZM2G073892 | bZIP transcription factor | 171 | Chr5 | 217591763 | 217592969 | 18.62 | 9.22 | 83.06 | −0.743 |
| ZmbZIP16 | GRMZM2G092137 | bZIP transcription factor | 156 | Chr7 | 83788712 | 83791125 | 17.57 | 9.24 | 82.49 | −0.835 |
| ZmbZIP17 | GRMZM2G093020 | common plant regulatory factor | 106 | Chr1 | 143352528 | 143354301 | 11.89 | 10.7 | 67.32 | −1.169 |
| ZmbZIP18 | GRMZM2G094352 | Transcription factor TGA4 | 384 | Chr10 | 49989443 | 49992484 | 42.53 | 8.9 | 56.89 | −0.376 |
| ZmbZIP19 | GRMZM2G095078 | EM binding protein 1-like protein | 386 | Chr7 | 19265565 | 19270520 | 40.28 | 6.28 | 69.32 | −0.69 |
| ZmbZIP20 | GRMZM2G098904 | Basic leucine zipper 9 | 324 | Chr5 | 87150348 | 87153356 | 34.29 | 5.37 | 57.99 | −0.373 |
| ZmbZIP21 | GRMZM2G103647 | Basic leucine zipper 9 | 282 | Chr9 | 109162612 | 109164995 | 30.14 | 5.2 | 54.59 | −0.526 |
| ZmbZIP22 | GRMZM2G120167 | DNA binding protein | 349 | Chr4 | 16152931 | 16158076 | 38.56 | 9.49 | 66.14 | −0.936 |
| ZmbZIP23 | GRMZM2G122846 | Putative bZIP transcription factor | 150 | Chr4 | 165667701 | 165668420 | 16.79 | 9.6 | 71.41 | −0.551 |
| ZmbZIP24 | GRMZM2G129247 | G-box-binding factor 4 | 254 | Chr6 | 150274418 | 150276924 | 27.09 | 7.76 | 68.69 | −0.585 |
| ZmbZIP25 | GRMZM2G131961 | TGACG motif-binding transcription factor 4 | 405 | Chr2 | 6719168 | 6724854 | 45.34 | 7.18 | 53.72 | −0.394 |
| ZmbZIP26 | GRMZM2G132868 | G-box-binding factor 4 | 257 | Chr8 | 100221799 | 100225360 | 27.04 | 6.79 | 66.88 | −0.532 |
| ZmbZIP27 | GRMZM2G136266 | Transcription factor PosF21 | 398 | Chr10 | 7921801 | 7925637 | 42.99 | 6.72 | 60.01 | −0.813 |
| ZmbZIP28 | GRMZM2G137046 | Transcription factor HY5 | 170 | Chr5 | 112670143 | 112675317 | 18.72 | 9.89 | 58.5 | −1.118 |
| ZmbZIP29 | GRMZM2G138340 | ABRE-binding factor Embp-2 | 140 | Chr1 | 291173937 | 291177741 | 14.85 | 4.77 | 51.14 | −0.714 |
| ZmbZIP30 | GRMZM2G146020 | Transcription factor PosF21 | 321 | Chr3 | 142765244 | 142769768 | 34.63 | 6.42 | 62.81 | −0.698 |
| ZmbZIP31 | GRMZM2G151295 | Basic leucine-zipper 52 | 353 | Chr9 | 131353502 | 131359160 | 38.05 | 6.14 | 55.83 | −0.645 |
| ZmbZIP32 | GRMZM2G153144 | Transcription factor HBP-1a | 350 | Chr5 | 74180653 | 74185259 | 37.32 | 5.42 | 57.41 | −1.019 |
| ZmbZIP33 | GRMZM2G157722 | ABA-INSENSITIVE 5 | 356 | Chr4 | 57698284 | 57703555 | 37.69 | 5.44 | 49.79 | −0.379 |
| ZmbZIP34 | GRMZM2G159134 | ABA-INSENSITIVE 5 | 333 | Chr3 | 186805980 | 186810898 | 36.25 | 6.69 | 59.57 | −0.71 |
| ZmbZIP35 | GRMZM2G160136 | putative bZIP transcription factor | 251 | Chr3 | 47233315 | 47236145 | 27.59 | 9.36 | 60.85 | −0.369 |
| ZmbZIP36 | GRMZM2G160902 | bZIP transcription factor 53 | 152 | Chr3 | 128957708 | 128959418 | 16.71 | 9.09 | 78.39 | −0.611 |
| ZmbZIP37 | GRMZM2G161009 | ABA-INSENSITIVE 5 | 324 | Chr8 | 119246143 | 119249647 | 35.33 | 6.38 | 52.91 | −0.657 |
| ZmbZIP38 | GRMZM2G171370 | G-box-binding factor 1 | 435 | Chr2 | 161590575 | 161596682 | 45.47 | 6.75 | 63.95 | −0.809 |
| ZmbZIP39 | GRMZM2G174284 | TGACG motif-binding transcription factor 6 | 335 | Chr1 | 51753616 | 51759590 | 37.43 | 8.59 | 56.99 | −0.524 |
| ZmbZIP40 | GRMZM2G175280 | DNA binding protein | 465 | Chr1 | 197089134 | 197092935 | 49.71 | 7.15 | 73.46 | −0.736 |
| ZmbZIP41 | GRMZM2G175870 | DNA binding protein | 229 | Chr6 | 156230705 | 156237177 | 24.84 | 5.36 | 48.29 | −0.49 |
| ZmbZIP42 | GRMZM2G177046 | Ocs element-binding factor 1 | 185 | Chr1 | 281401644 | 281402596 | 20.19 | 10.5 | 71.82 | −0.443 |
| ZmbZIP43 | GRMZM2G180847 | Putative transcription factor PosF21 | 371 | Chr2 | 194071950 | 194077869 | 39.35 | 5.96 | 59.58 | −0.655 |
| ZmbZIP44 | GRMZM2G361611 | Ocs element-binding factor 1 | 162 | Chr4 | 239887667 | 239890292 | 17.13 | 6.29 | 52.56 | −0.107 |
| ZmbZIP45 | GRMZM2G361847 | TGACG motif-binding transcription factor 6 | 333 | Chr7 | 174267403 | 174273062 | 37.15 | 8.9 | 55.08 | −0.521 |
| ZmbZIP46 | GRMZM2G368491 | Common plant regulatory factor 7 | 142 | Chr3 | 227566001 | 227567569 | 15.53 | 9.75 | 50.25 | −0.523 |
| ZmbZIP47 | GRMZM2G425920 | Transcription factor HY5-like | 215 | Chr10 | 122253987 | 122259270 | 23.6 | 10.38 | 69.63 | −1.171 |
| ZmbZIP48 | GRMZM2G438293 | ABA-INSENSITIVE 5 | 238 | Chr8 | 165990148 | 165991775 | 24.92 | 5.73 | 68.24 | −0.524 |
| ZmbZIP49 | GRMZM2G438652 | Basic leucine zipper 25-like | 214 | Chr7 | 3077372 | 3078435 | 22.3 | 11.35 | 59.78 | −0.129 |
| ZmbZIP50 | GRMZM2G448607 | Ocs element-binding factor 1 | 135 | Chr6 | 24280540 | 24281904 | 15.01 | 10.14 | 71.91 | −0.733 |
| ZmbZIP51 | GRMZM2G043600 | Transcription factor RF2a-like | 275 | Chr2 | 216709894 | 216711451 | 30.53 | 6.88 | 66.56 | −0.907 |
| ZmbZIP52 | GRMZM2G479760 | ABA-INSENSITIVE 5 | 377 | Chr5 | 210099032 | 210100747 | 40.16 | 10.03 | 57.18 | −0.659 |
| ZmbZIP53 | GRMZM2G479885 | Ocs element-binding factor 1 | 151 | Chr1 | 147170752 | 147172309 | 16.86 | 6.93 | 73.52 | −0.716 |
| ZmbZIP54 | GRMZM5G858197 | G-box-binding factor 4 | 260 | Chr3 | 175672623 | 175675787 | 27.55 | 8.95 | 74.98 | −0.673 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.; Lu, X.; Zhang, P.; Wang, G.; Wei, L.; Wang, T. Systematic Analysis of Differentially Expressed Maize ZmbZIP Genes between Drought and Rewatering Transcriptome Reveals bZIP Family Members Involved in Abiotic Stress Responses. Int. J. Mol. Sci. 2019, 20, 4103. https://doi.org/10.3390/ijms20174103
Cao L, Lu X, Zhang P, Wang G, Wei L, Wang T. Systematic Analysis of Differentially Expressed Maize ZmbZIP Genes between Drought and Rewatering Transcriptome Reveals bZIP Family Members Involved in Abiotic Stress Responses. International Journal of Molecular Sciences. 2019; 20(17):4103. https://doi.org/10.3390/ijms20174103
Chicago/Turabian StyleCao, Liru, Xiaomin Lu, Pengyu Zhang, Guorui Wang, Li Wei, and Tongchao Wang. 2019. "Systematic Analysis of Differentially Expressed Maize ZmbZIP Genes between Drought and Rewatering Transcriptome Reveals bZIP Family Members Involved in Abiotic Stress Responses" International Journal of Molecular Sciences 20, no. 17: 4103. https://doi.org/10.3390/ijms20174103