De Novo Transcriptome Analysis Reveals Abundant Gonad-specific Genes in the Ovary and Testis of Henosepilachna vigintioctopunctata

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
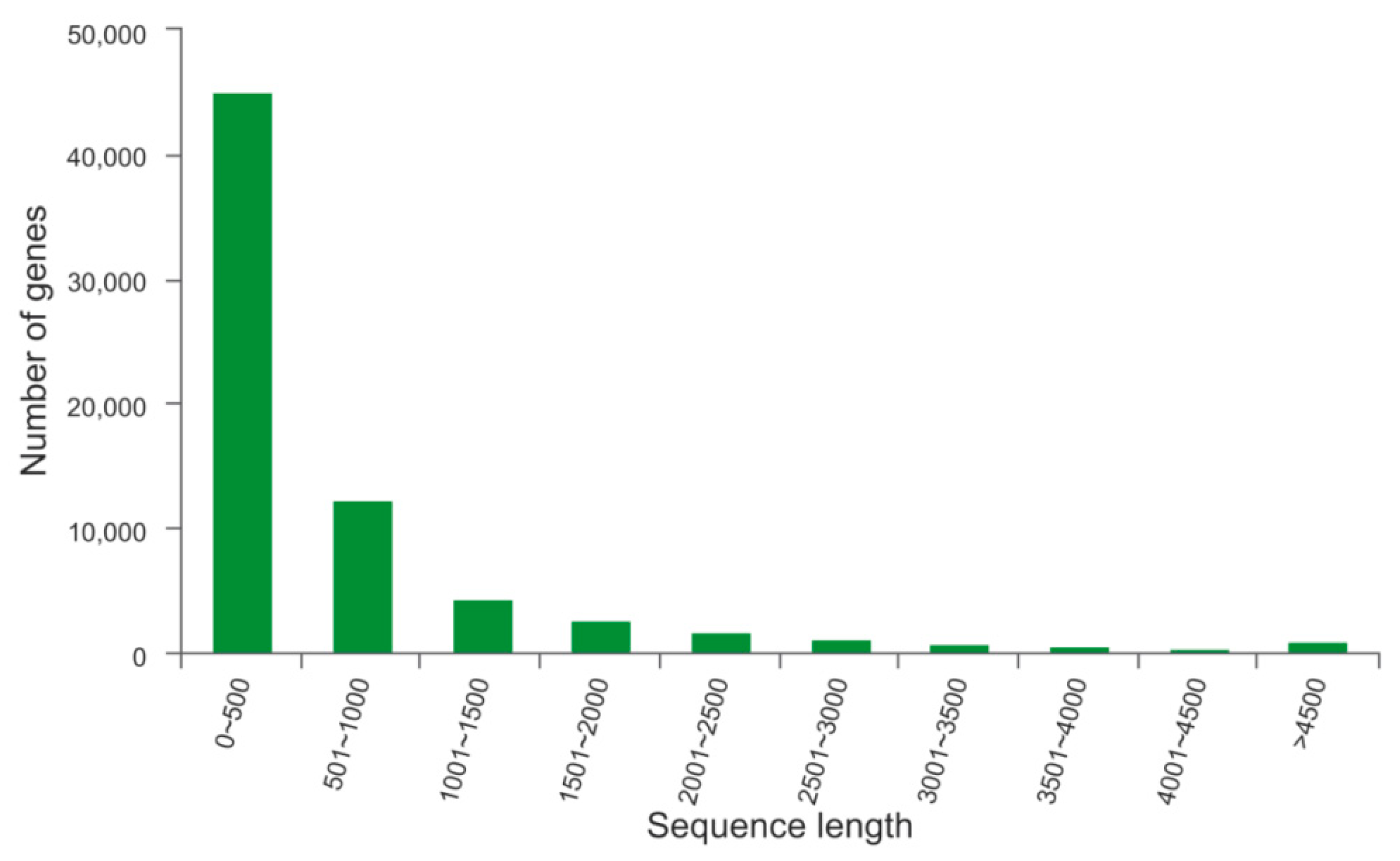
2.1. Illumina Sequencing and Assembly
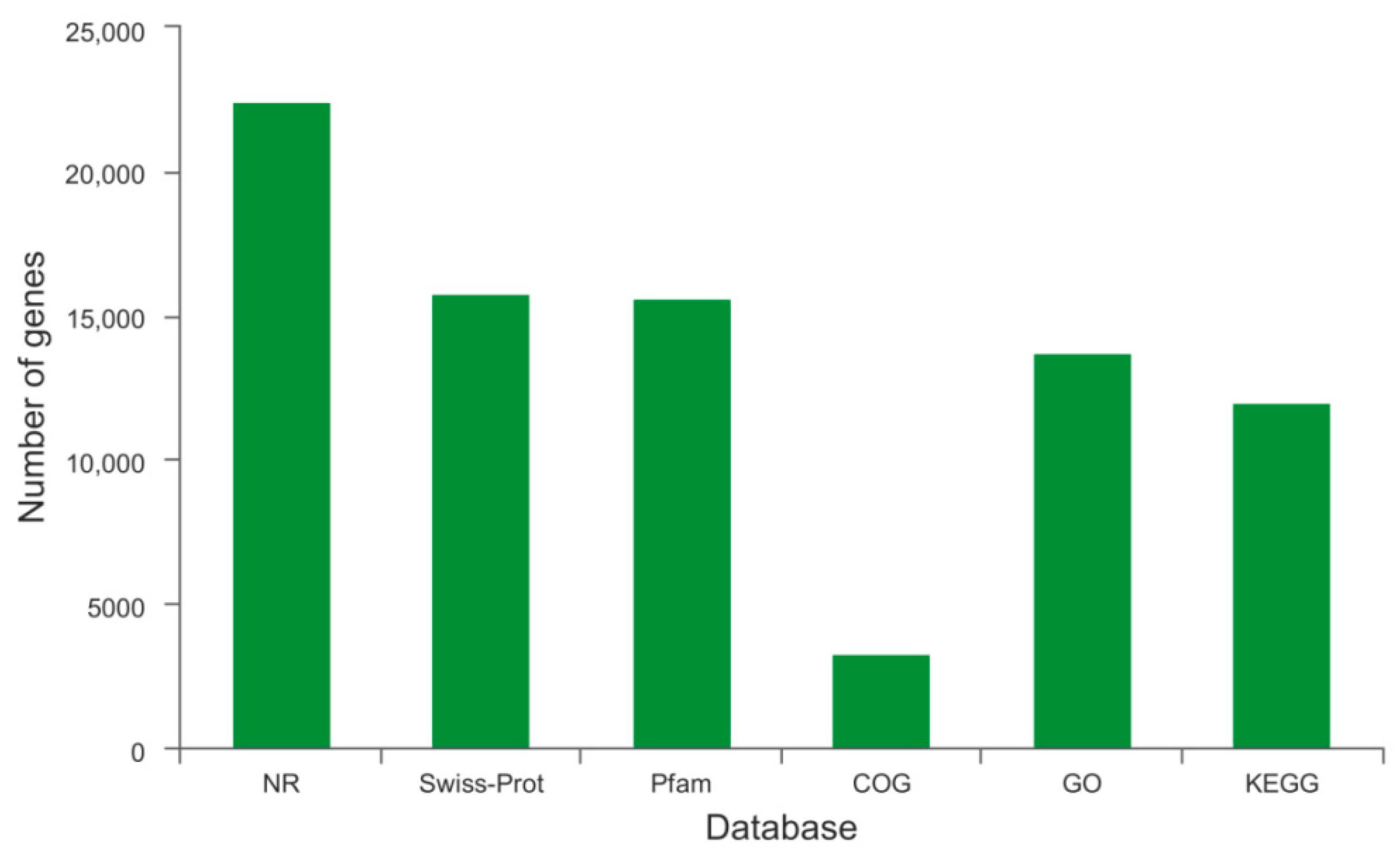
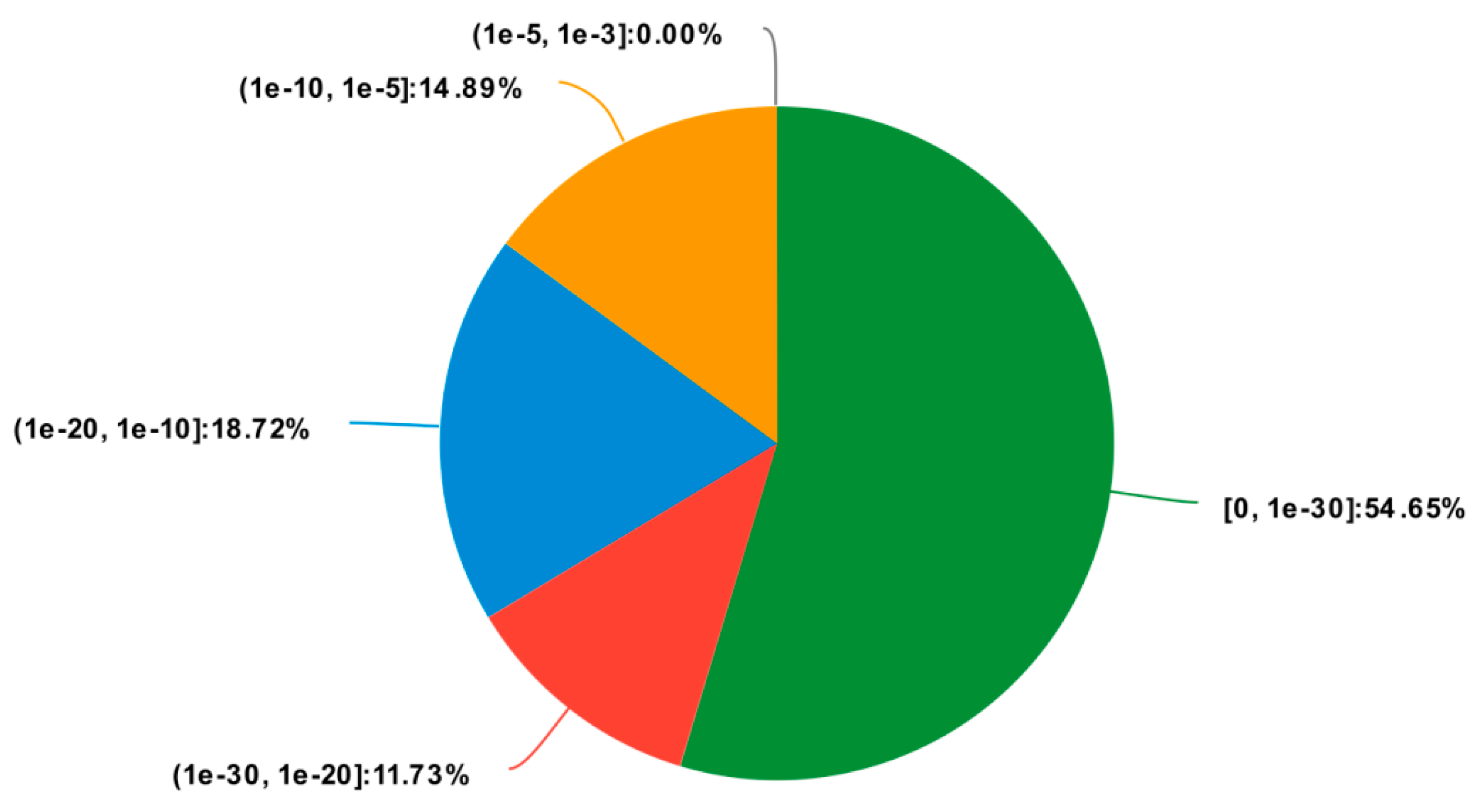
2.2. Annotation and Functional Classification
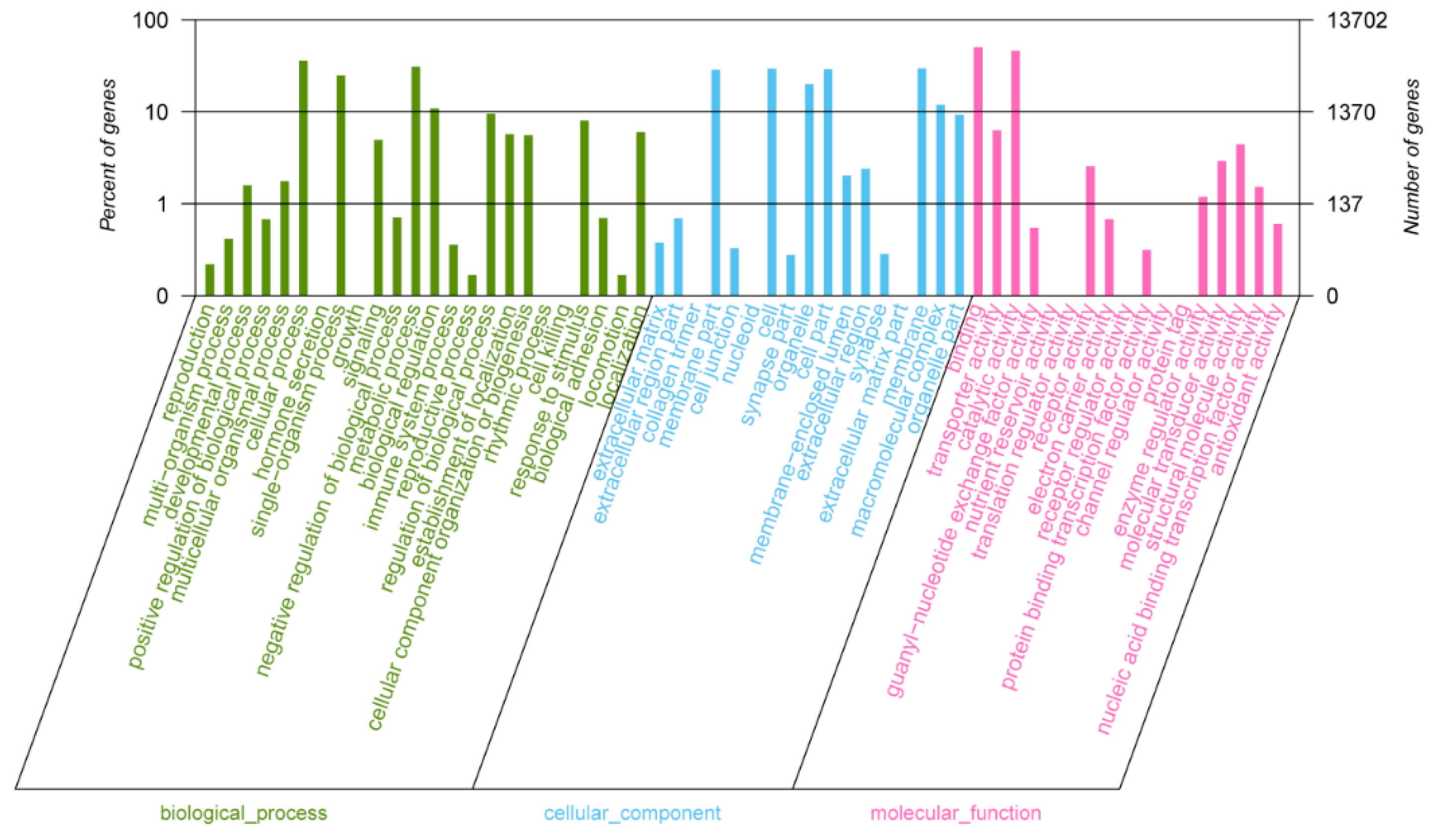
2.3. Gene Ontology Analysis
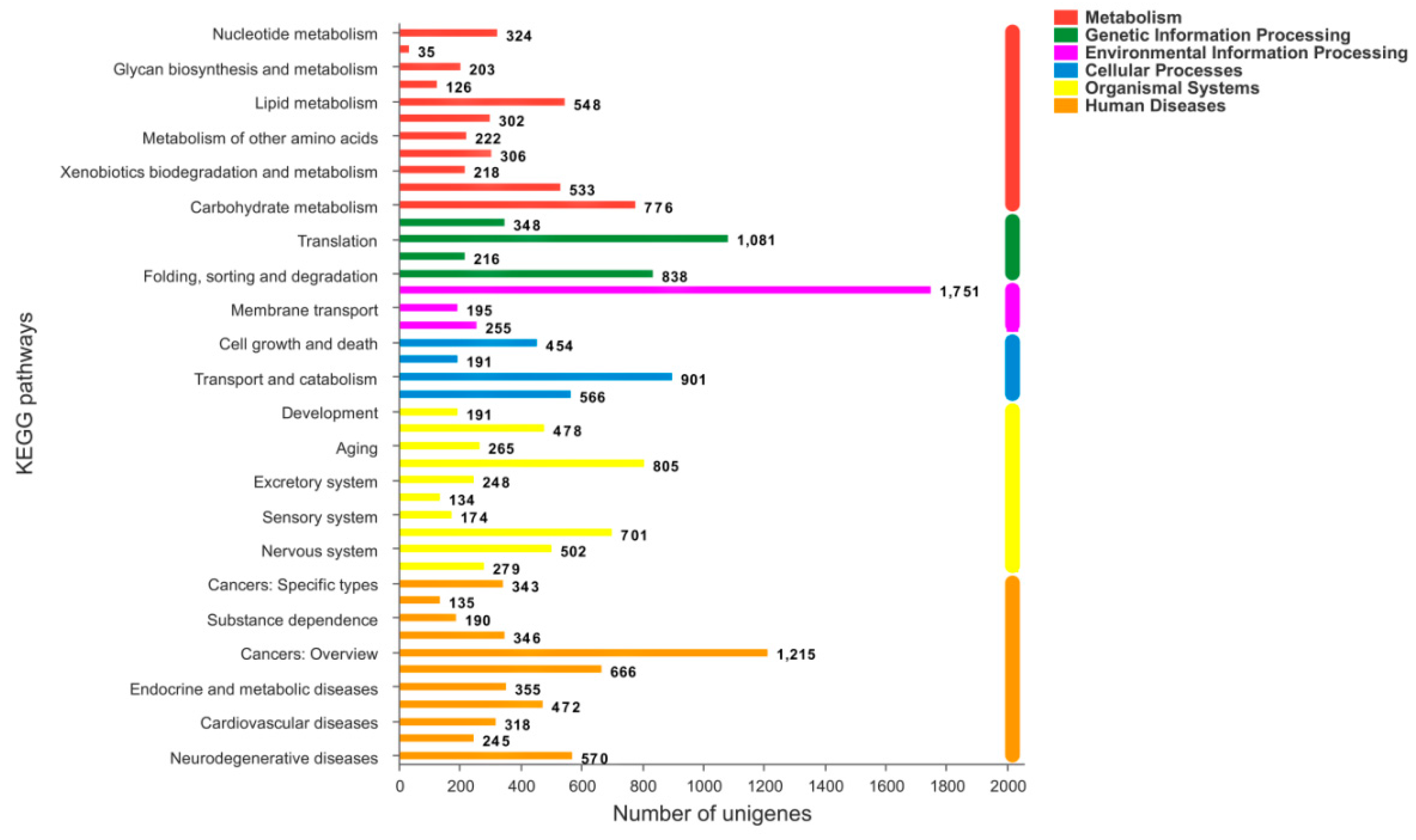
2.4. KEGG Analysis
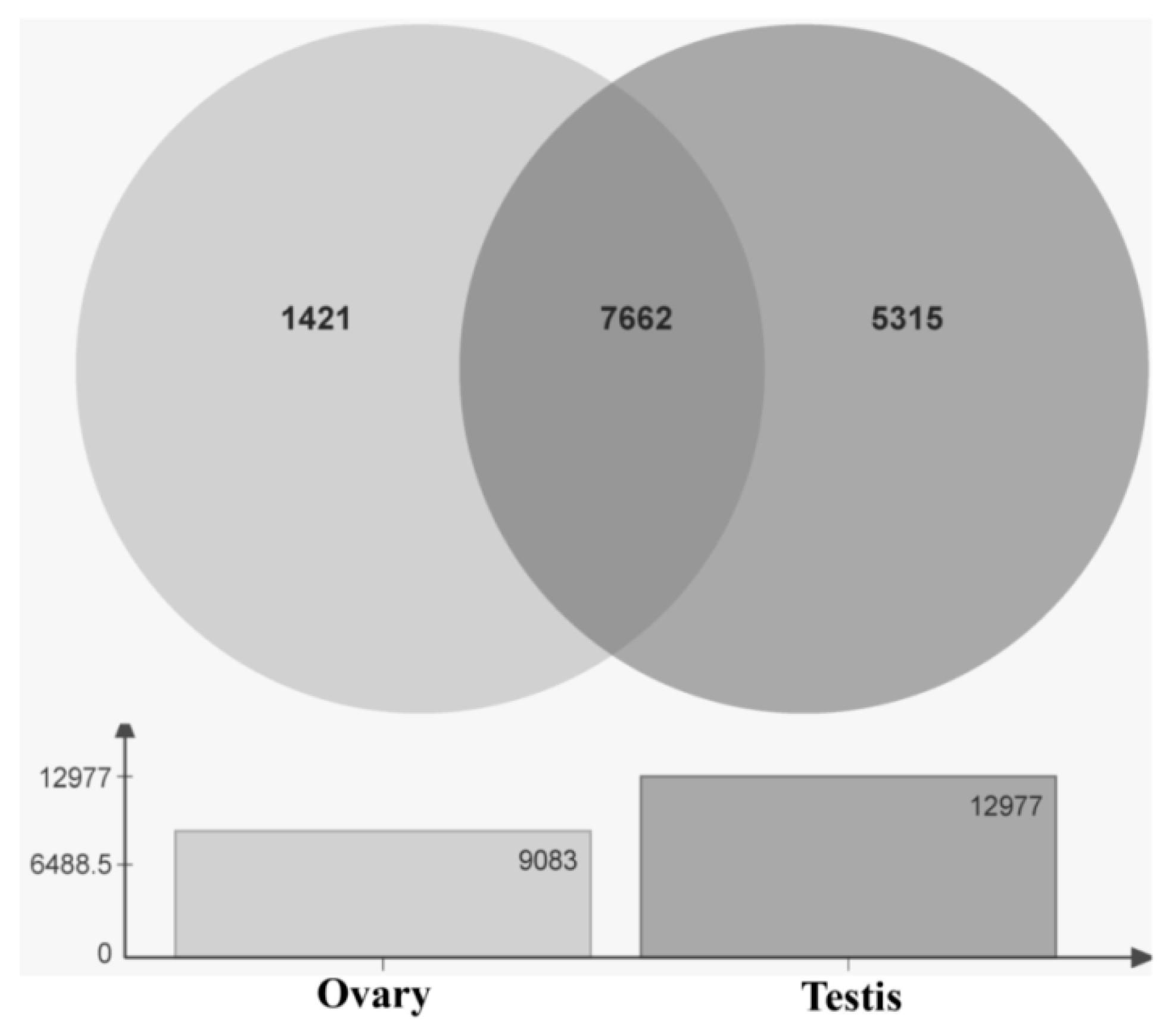
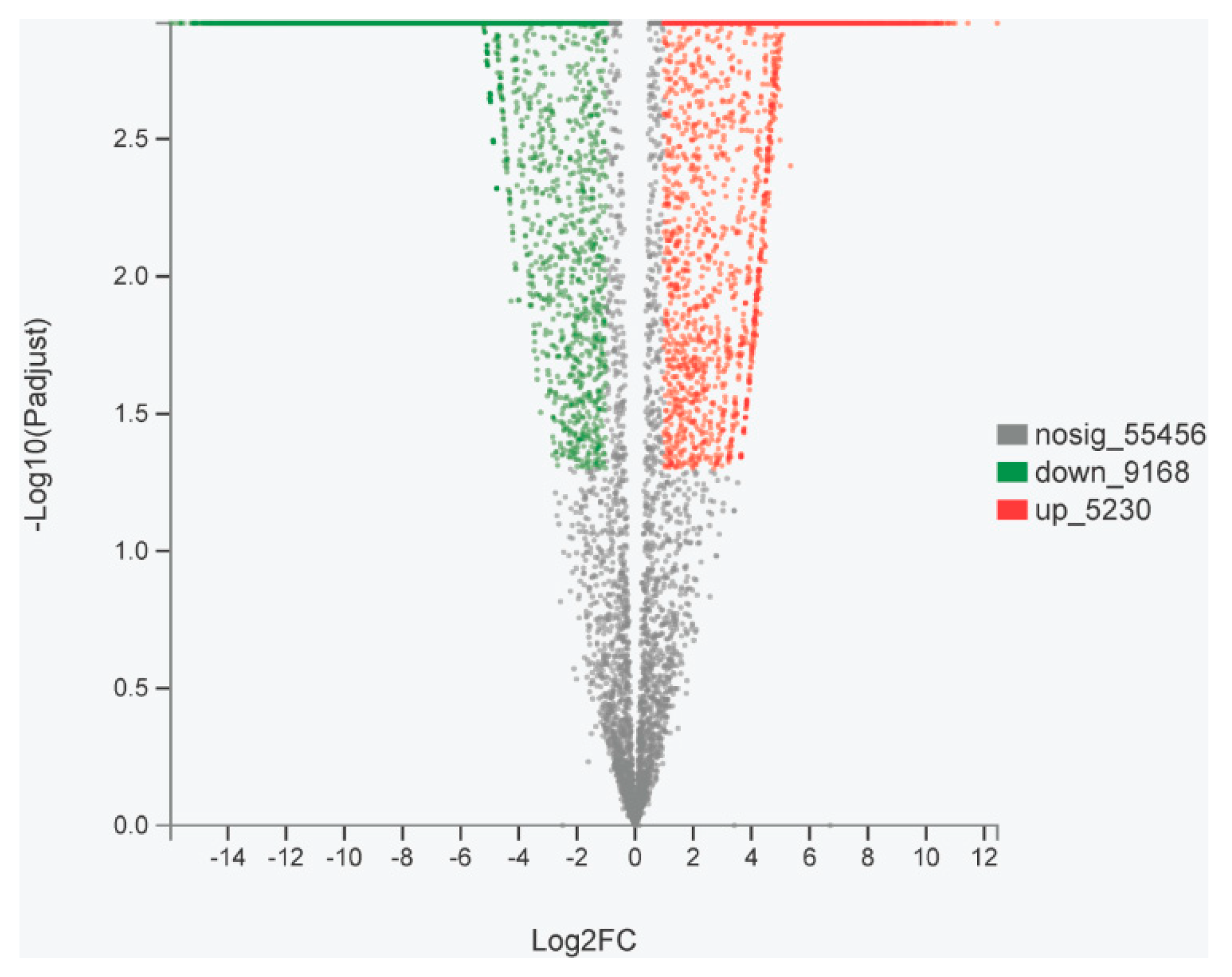
2.5. Analysis of Differentially Expressed Genes in Ovary and Testis
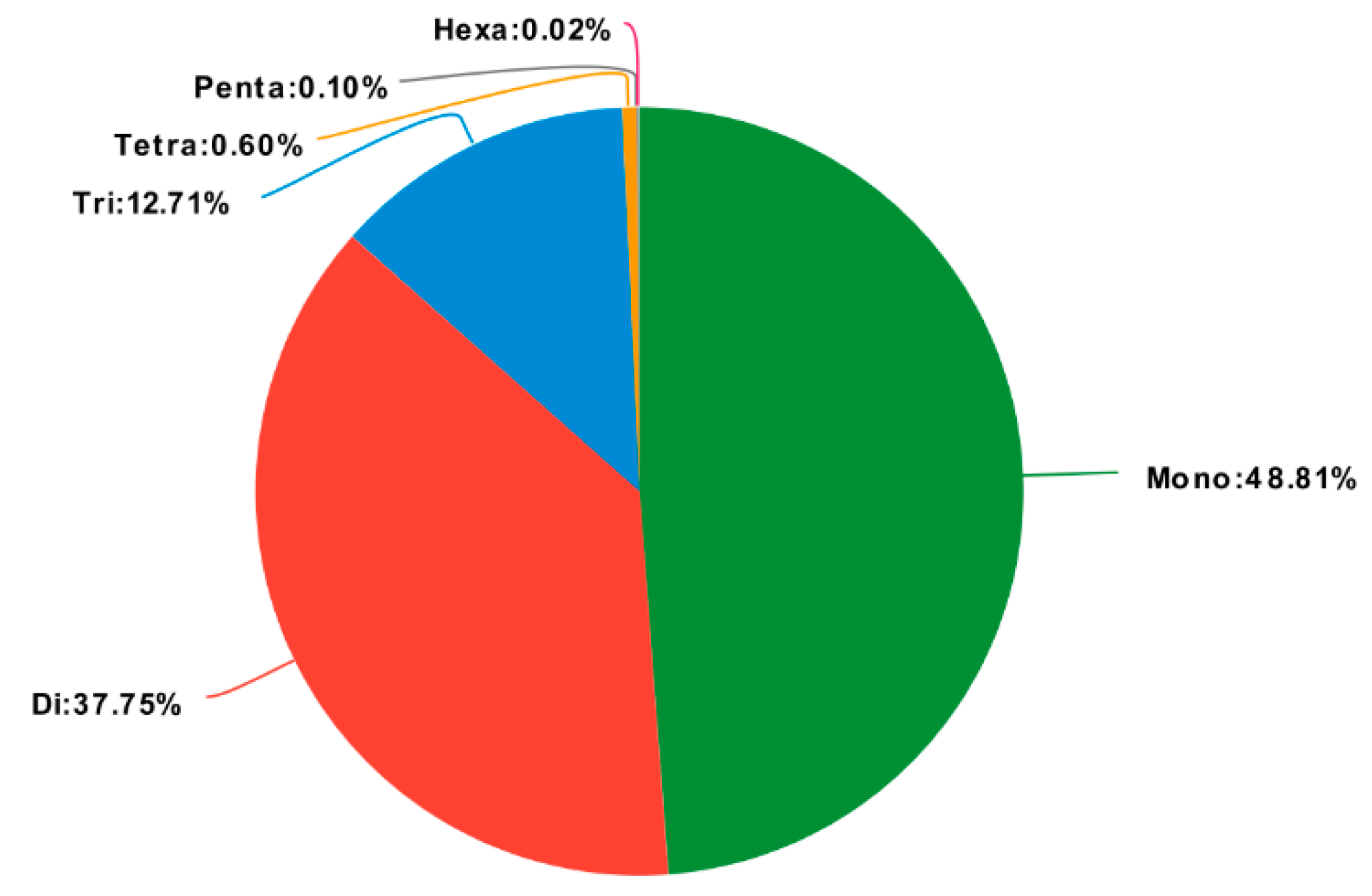
2.6. Discovery of Molecular Markers
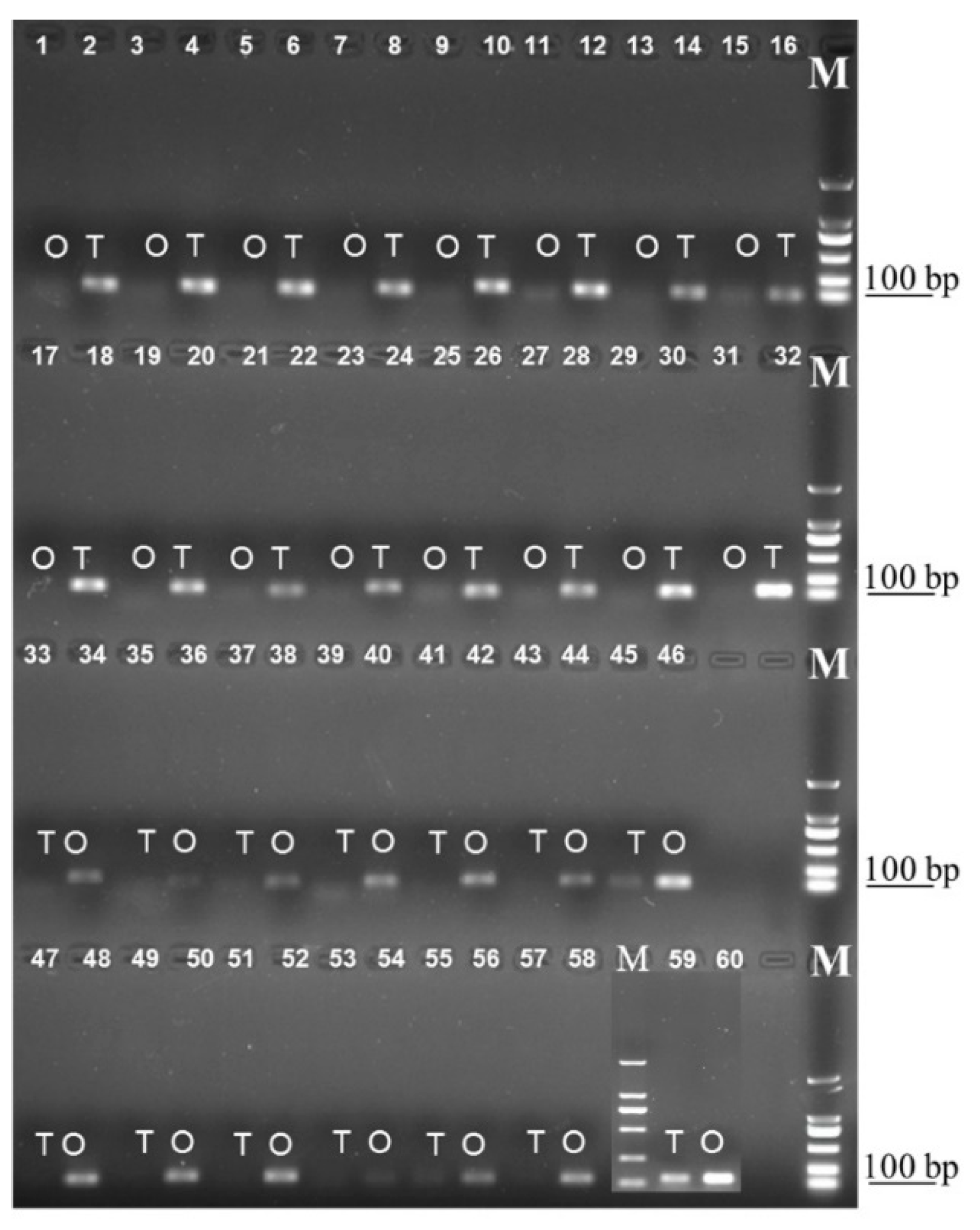
2.7. Semi-RT-qPCR and RT-qPCR Confirmation of Gonad-biased Genes
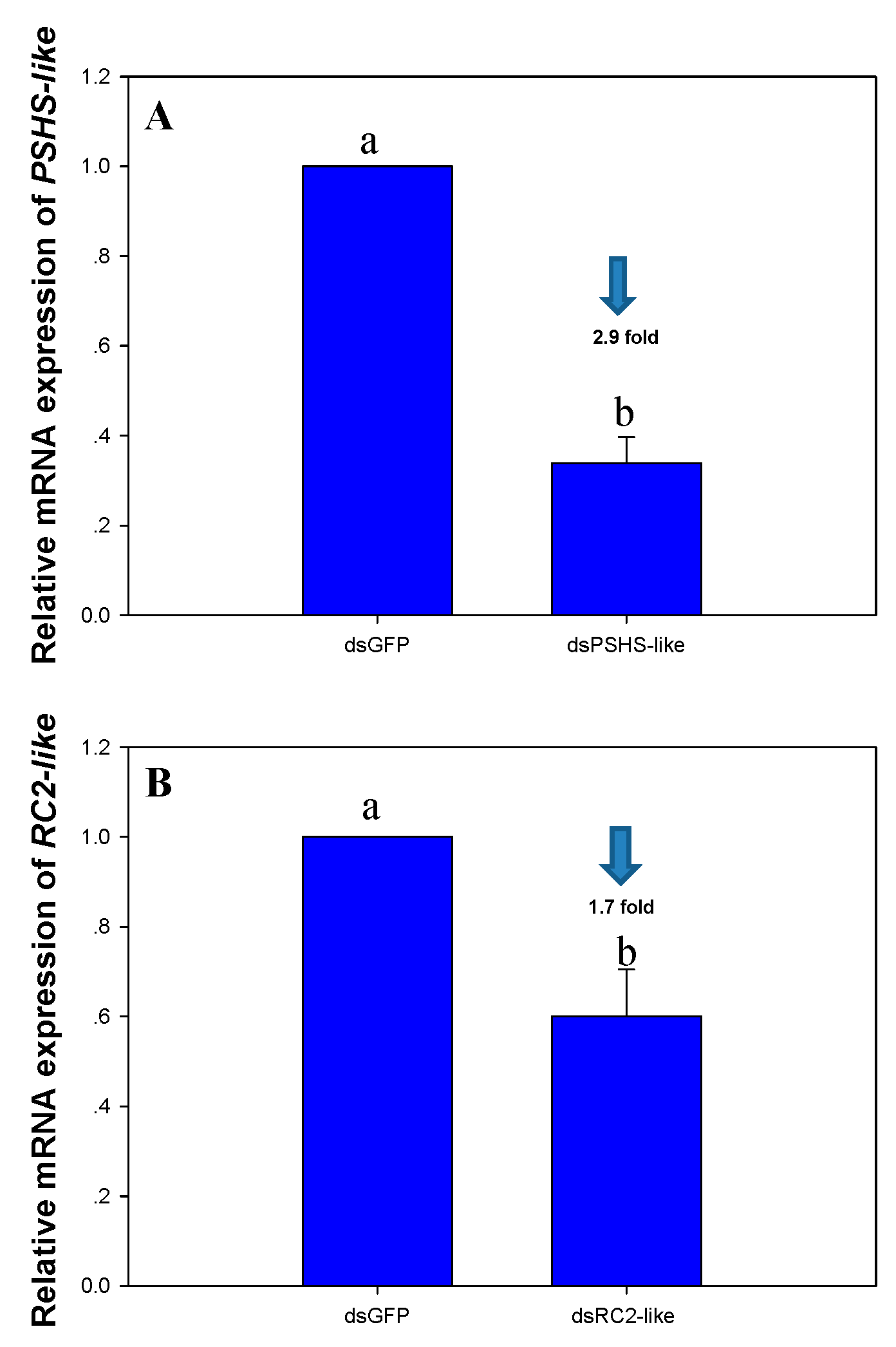
2.8. The Effects of Dietary RNAi of dsPSHS-like and dsRC2-like on PSHS-like and RC2-like Gene Expression
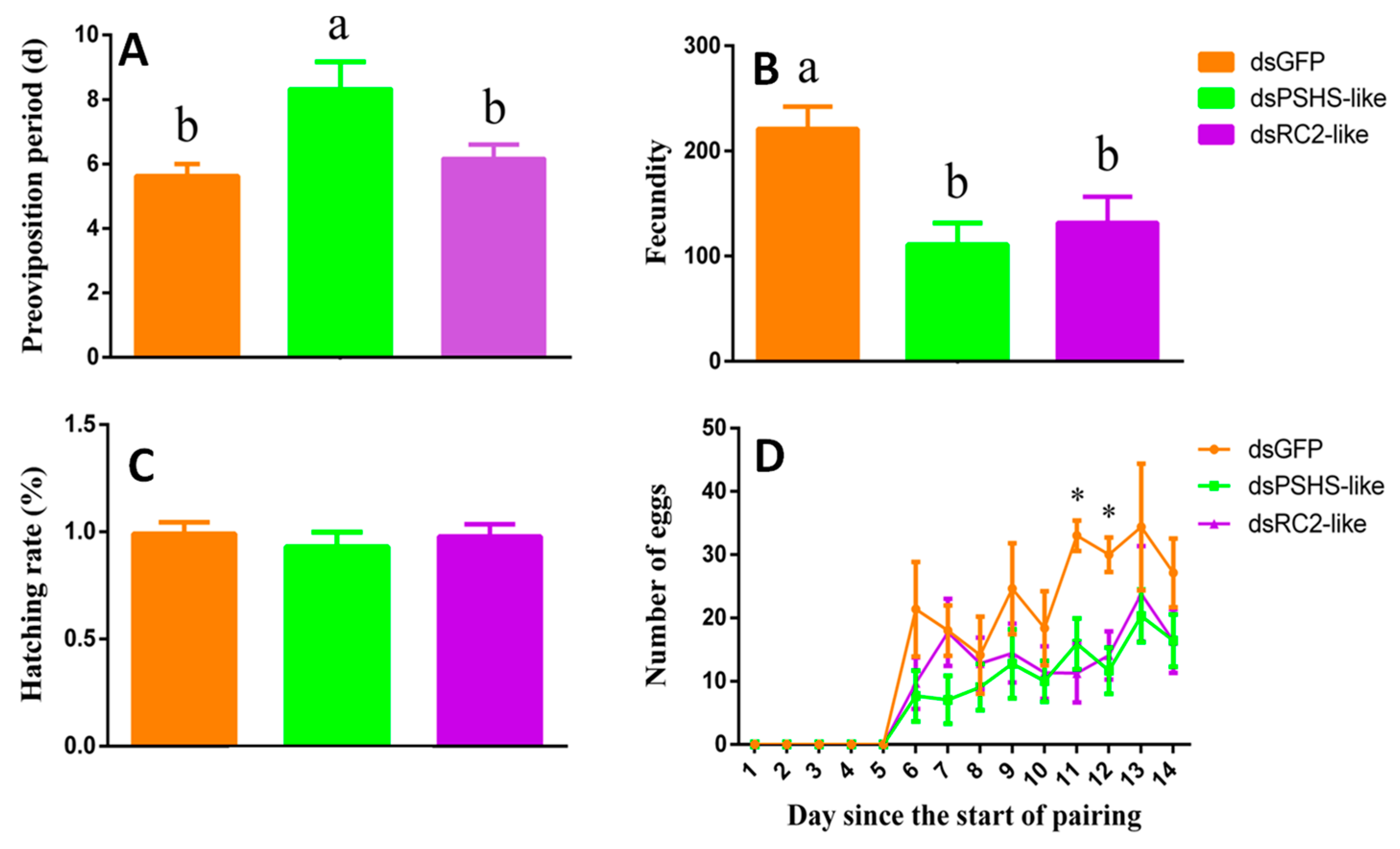
2.9. The Effects of Silencing dsPSHS-like and dsRC2-like on the Phenotype of H. vigintioctopunctata
3. Discussion
4. Materials and Methods
4.1. Insect Rearing and Sample Collection
4.2. RNA Isolation, Library Preparation, and Sequencing
4.3. Sequence Assembly and Functional Annotation
4.4. Identification of Differentially Expressed Genes in Ovary and Testis
4.5. Molecular Marker Detection
4.6. Semi-RT-qPCR and RT-qPCR Confirmation of Gonad-biased Genes
4.7. Dietary RNAi Toxicity Assay
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Nr | Non-redundant |
| COG | Clusters of Orthologous Groups of proteins |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| RT-qPCR | reverse transcriptase-quantitative polymerase chain reaction |
| SSRs | simple sequence repeats |
References
- Ghosh, S.K.; Senapati, S.K. Biology and seasonal fluctuation of Henosepilachna vigintioctopunctata Fabr. on brinjal under Terai region of West Bengal. Indian J. Agric. Res. 2001, 35, 149–154. [Google Scholar]
- Zhou, L.; Xie, B.G.; Wang, X.P. Population dynamic of Henosepilachna vigintioctopunctata in different host plants in Jianghan plain. China J. North. Hort. 2015, 11, 103–105. [Google Scholar]
- Shinogi, T.; Hamanishi, Y.; Otsu, Y.; Wang, Y.Q.; Nonomura, T.; Matsuda, Y.; Toyoda, H.; Narusaka, Y.; Tosaa, Y.; Mayama, S. Role of induced resistance in interactions of Epilachna vigintioctopunctata with host and non-host plant species. Plant Sci. 2005, 168, 1477–1485. [Google Scholar] [CrossRef]
- Venkatesha, M.G. Seasonal occurrence of Henosepilachna vigintioctopunctata (F.) (Coleoptera: Coccinellidae) and its parasitoid on Ashwagandha in India. J. Asia-Pac. Entomol. 2006, 9, 265–268. [Google Scholar] [CrossRef]
- Pang, X.F.; Mao, J.L. Two new species of Vibidia luliangensis are described. Econ. Insects China 1979, 22, 468–470. [Google Scholar]
- Sharma, A.; Thakur, A.; Kaur, S.; Pati, P.K. Effect of Alternaria alternata on the coccinellid pest Henosepilachna vigintioctopunctata and its implications for biological pest management. J. Pest Sci. 2012, 85, 513–518. [Google Scholar] [CrossRef]
- Kawazu, K. Rearing the 28-spotted ladybird beetle, Henosepilachna vigintioctopunctata (Coleoptera: Coccinelidae), with a switchover from host plant leaves to artificial diet. Appl. Entomol. Zool. 2014, 49, 359–362. [Google Scholar] [CrossRef]
- Gui, L.Y.; Meng, G.L.; Gong, X.W. Resistance identification of eggplant varieties in field area. China J. Hubei Agric. Sci. 1995, 5, 34–35. [Google Scholar]
- Fu, Z.; Hu, B.; Tu, X.Y. Observation on the oviposition of Henosepilachna vigintioctopunctata (Fabricius). J. Anhui Agri. Sci. 2009, 37, 16423–16499. [Google Scholar]
- Farhadi, R.; Allahyari, H.; Chi, H. Life table and predation capacity of Hippodamia variegata (Coleoptera: Coccinellidae) feeding on Aphis fabae (Hemiptera: Aphididae). Biol. Control 2011, 59, 83–89. [Google Scholar] [CrossRef]
- Wang, G.H.; Cao, B.; Huang, C.L.; Ge, F. Effects of Henosepilachna vigintioctopunctata Fabricius infestation on resistant enzyme activity in the leaves of tomato genotypes with different JA pathways. China J. Appl. Entomol. 2015, 52, 1236–1243. [Google Scholar]
- Wang, Z.L.; Li, C.R.; Yuan, J.J.; Li, S.X.; Wang, X.P.; Chi, H. Demographic comparison of Henosepilachna vigintioctopunctata (F.) (Coleoptera: Coccinellidae) reared on three cultivars of Solanum melongena L. and a wild host plant Solanum nigrum L. J. Econ. Entomol. 2017, 110, 2084–2091. [Google Scholar] [CrossRef] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Bellés, X. Beyond Drosophila: RNAi in vivo and functional genomics in insects. Annu. Rev. Entomol. 2010, 55, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Burand, J.P.; Hunter, W.B. RNAi: Future in insect management. J. Invertebr. Pathol. 2013, 112, S68–S74. [Google Scholar] [CrossRef] [PubMed]
- Katoch, R.; Sethi, A.; Thakur, N.; Murdock, L.L. RNAi for insect control: Current perspective and future challenges. Appl. Biochem. Biotech. 2013, 171, 847–873. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Khan, S.A.; Heckel, D.G.; Bock, R. Next-generation insect-resistant plants: RNAi-mediated crop protection. Trends Biotechnol. 2017, 35, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Mogilicherla, K.; Gurusamy, D.; Chen, X.; Chereddy, S.C.R.R.; Palli, S.R. Double-stranded RNA binding protein, Staufen, is required for the initiation of RNAi in coleopteran insects. Proc. Natl. Acad. Sci. USA 2018, 115, 8334–8339. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Xu, J.; Palli, R.; Ferguson, J.; Palli, S.R. Ingested RNA interference for managing the populations of the Colorado potato beetle, Leptinotarsa decemlineata. Pest Manag. Sci. 2011, 67, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.D.; Killiny, N. Effect of parental RNA interference of a transformer-2 homologue on female reproduction and offspring sex determination in Asian citrus psyllid. Physiol. Entomol. 2018, 43, 42–50. [Google Scholar] [CrossRef]
- Moriyama, M.; Hosokawa, T.; Tanahashi, M.; Nikoh, N.; Fukatsu, T. Suppression of bedbug’s reproduction by RNA interference of vitellogenin. PLoS ONE 2016, 11, e0153984. [Google Scholar] [CrossRef] [PubMed]
- Basnet, S.; Kamble, S.T. Knockdown of the chromatin remodeling gene brahma by RNA interference reduces reproductive fitness and lifespan in common bed bug (Hemiptera: Cimicidae). J. Med. Entomol. 2018, 55, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Basnet, S.; Kamble, S.T. RNAi-mediated knockdown of vATPase subunits affects survival and reproduction of bed bugs (Hemiptera: Cimicidae). J. Med. Entomol. 2018, 55, 540–546. [Google Scholar] [CrossRef]
- Xu, L.J.; Hou, Q.L.; Zhao, Y.J.; Lu, L.H.; Li, B.Y.; Ni, Z.F.; Liang, R.Q. Silencing of a lipase maturation factor 2-like gene by wheat-mediated RNAi reduces the survivability and reproductive capacity of the grain aphid, Sitobion avenae. Arch. Insect Biochem. 2017, 95, e21392. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.P.; Kassa, A.; Hu, X.; Robeson, J.; McMahon, M.; Richtman, N.M.; Steimel, J.P.; Kernodle, B.M.; Crane, V.C.; Sandahl, G.; et al. Control of western corn rootworm (Diabrotica virgifera virgifera) reproduction through plant-mediated RNA interference. Sci. Rep. 2017, 7, 12591. [Google Scholar] [CrossRef]
- Li, K.L.; Wan, P.J.; Wang, W.X.; Lai, F.X.; Fu, Q. Ran involved in the development and reproduction is a potential target for RNA-interference-based pest management in Nilaparvata lugens. PLoS ONE 2015, 10, e0142142. [Google Scholar] [CrossRef]
- Luo, J.; Liang, S.; Li, J.; Xu, Z.; Li, L.; Zhu, B.; Li, Z.; Lei, C.; Lindsey, K.; Chen, L.; et al. A transgenic strategy for controlling plant bugs (Adelphocoris suturalis) through expression of double-stranded RNA homologous to fatty acyl-coenzyme A reductase in cotton. NewPhytol. 2017, 215, 1173–1185. [Google Scholar] [CrossRef]
- Ali, M.W.; Zheng, W.; Sohail, S.; Li, Q.; Zheng, W.; Zhang, H. A genetically enhanced sterile insect technique against the fruit fly, Bactrocera dorsalis (Hendel) by feeding adult double-stranded RNAs. Sci. Rep. 2017, 7, 4063. [Google Scholar] [CrossRef]
- Su, Y.L.; Li, J.M.; Li, M.; Luan, J.B.; Ye, X.D.; Wang, X.W.; Liu, S.S. Transcriptomic analysis of the salivary glands of an invasive whitefly. PLoS ONE 2012, 7, e39303. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.D.; Su, Y.L.; Zhao, Q.Y.; Xia, W.Q.; Liu, S.S.; Wang, X.W. Transcriptomic analyses reveal the adaptive features and biological differences of guts from two invasive whitefly species. BMC Genomics 2014, 15, 370. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.Y.; Wang, Y.; Wu, W.J.; Zhao, D.; Xue, J.; Zhang, B.Q.; Shen, Z.C.; Zhang, C.X. De novo intestine-specific transcriptome of the brown planthopper Nilaparvata lugens revealed potential functions in digestion, detoxification and immune response. Genomics 2012, 99, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ji, R.; Ye, W.; Chen, H.; Lai, W.; Fu, Q.; Lou, Y. Transcriptome analysis of fat bodies from two brown planthopper (Nilaparvata lugens) populations with different virulence levels in rice. PLoS ONE 2014, 9, e88528. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Fang, Q.; Zhao, L.; Xia, H.; Zhou, Y.; Xiao, J.; Li, K.; Ye, G. De novo assembly and developmental transcriptome analysis of the small white butterfly Pieris rapae. PLoS ONE 2016, 11, e0159258. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, Y.X.; Jiang, G.F. Corrigendum: De novo assembly and characterization of the testis transcriptome and development of EST-SSR markers in the cockroach Periplaneta americana. Sci. Rep. 2015, 5, 16547. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, J.; Cui, M.; Tao, J.; Luo, Y. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 2016, 6, 26652. [Google Scholar] [CrossRef]
- Wei, D.; Li, H.M.; Yang, W.J.; Wei, D.D.; Dou, W.; Huang, Y.; Wang, J.J. Transcriptome profiling of the testis reveals genes involved in spermatogenesis and marker discovery in the oriental fruit fly, Bactrocera dorsalis. Insect Mol. Biol. 2015, 24, 41–57. [Google Scholar] [CrossRef]
- Parisi, M.; Nuttall, R.; Naiman, D.; Bouffard, G.; Malley, J.; Andrews, J.; Eastman, S.; Oliver, B. Paucity of genes on the Drosophila X chromosome showing male-biased expression. Science 2003, 299, 697–700. [Google Scholar] [CrossRef]
- Goldman, T.D.; Arbeitman, M.N. Genomic and functional studies of Drosophila sex hierarchy regulated gene expression in adult head and nervous system tissues. PLoS Genet. 2007, 3, e216. [Google Scholar] [CrossRef]
- Catalán, A.; Hutter, S.; Parsch, J. Population and sex differences in Drosophila melanogaster brain gene expression. BMC Genomics 2012, 13, 654. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.W.; Wright, A.E.; Zimmer, F.; Dean, R.; Montgomery, S.H.; Pointer, M.A.; Mank, J.E. Sexual selection drives evolution and rapid turnover of male gene expression. Proc. Natl. Acad. Sci. USA 2015, 112, 4393–4398. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, X.; Zou, Z.; Jia, X.; Wang, Y.; Zhang, Z. Transcriptome analysis of the differences in gene expression between testis and ovary in green mud crab (Scylla paramamosain). BMC Genomics 2014, 15, 585. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.L.; Liu, P.; Jia, F.L.; Li, J.; Gao, B.Q. De novo transcriptome analysis of Portunus trituberculatus ovary and testis by RNA-Seq: Identification of genes involved in gonadal development. PLoS ONE 2015, 10, e0128659. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.X.; Wei, P.Y.; Zhang, B.; Zhao, Y.Z.; Zeng, D.G.; Chen, X.L.; Li, M.; Chen, X.H. Gonadal transcriptomic analysis and differentially expressed genes in the testis and ovary of the Pacific white shrimp (Litopenaeus vannamei). BMC Genomics 2015, 16, 1006. [Google Scholar] [CrossRef] [PubMed]
- Lü, J.; Chen, S.M.; Guo, M.J.; Ye, C.Y.; Qiu, B.L.; Wu, J.H.; Yang, C.X.; Pan, H.P. Correction: Selection and validation of reference genes for RT-qPCR analysis of the ladybird beetle Henosepilachna vigintioctopunctata. Front. Physiol. 2019, 10, 981. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- MISA-MIcroSAtellite Identification Tool. Available online: http://pgrc.ipk-gatersleben.de/misa/misa.html (accessed on 22 July 2019).
- E-RNAi-Design of RNAi Constructs. Available online: https://www.dkfz.de/signaling/e-rnai3// (accessed on 22 July 2019).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Results | |
| Number of ovary raw reads | 133,674,920 |
| Number of testis raw reads | 137,273,168 |
| Number of total raw reads | 270,948,088 |
| Number of ovary clean reads | 132,895,862 |
| Number of testis clean reads | 136,381,100 |
| Number of total clean reads | 269,276,962 |
| Assembly results | |
| Number of transcripts | 99,718 |
| Average length of transcripts (bp) | 780 |
| Minimum transcript size (bp) | 201 |
| Maximum transcript size (bp) | 16,278 |
| N50 | 1284 |
| Database | Nr | Swiss-Prot | Pfam | COG | GO | KEGG | Total |
|---|---|---|---|---|---|---|---|
| Unigenes | 22,472 | 15,765 | 15,617 | 3261 | 13,702 | 11,970 | 24,317 |
| Species | Unigenes Number | Percentage (%) |
|---|---|---|
| Tribolium castaneum | 5069 | 27.73 |
| Anoplophora glabripennis | 3490 | 15.65 |
| Aethina tumida | 1655 | 7.42 |
| Leptinotarsa decemlineata | 1455 | 6.52 |
| Onthophagus taurus | 535 | 2.40 |
| Cryptotermes secundus | 452 | 2.03 |
| Dendroctonus ponderosae | 446 | 2.00 |
| Nicrophorus vespilloides | 364 | 1.63 |
| Agrilus planipennis | 358 | 1.61 |
| Branchiostoma belcheri | 303 | 1.36 |
| Amyelois transitella | 274 | 1.23 |
| Lasius niger | 255 | 1.14 |
| Centruroides sculpturatus | 185 | 0.83 |
| Spodoptera litura | 178 | 0.80 |
| others | 7285 | 32.66 |
| SSR Information | Number |
|---|---|
| Total number of identified SSRs | 4964 |
| Number of unigenes containing SSRs | 4213 |
| Number of unigenes containing more than 1 SSR | 303 |
| Mono-nucleotide repeats | 2423 |
| Di-nucleotide repeats | 1874 |
| Tri-nucleotide repeats | 631 |
| Tetra-nucleotide repeats | 30 |
| Penta-nucleotide repeats | 5 |
| Hexa-nucleotide repeats | 1 |
| Database | Software | Format | Parameter |
|---|---|---|---|
| NR | DIAMOND | v0.8.37.99 | 1e−5 |
| Swiss-Prot | DIAMOND | v0.8.37.99 | 1e−5 |
| Pfam | HMMER3 | 3.1b2 | default parameters |
| COG | DIAMOND | v0.8.37.99 | 1e−5 |
| GO | BLAST2GO | 2.5.0 | default parameters |
| KEGG | KOBAS | 2.1.1 | default parameters |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, W.; Lü, J.; Guo, M.; Chen, S.; Qiu, B.; Sang, W.; Yang, C.; Zhang, Y.; Pan, H. De Novo Transcriptome Analysis Reveals Abundant Gonad-specific Genes in the Ovary and Testis of Henosepilachna vigintioctopunctata. Int. J. Mol. Sci. 2019, 20, 4084. https://doi.org/10.3390/ijms20174084
Guo W, Lü J, Guo M, Chen S, Qiu B, Sang W, Yang C, Zhang Y, Pan H. De Novo Transcriptome Analysis Reveals Abundant Gonad-specific Genes in the Ovary and Testis of Henosepilachna vigintioctopunctata. International Journal of Molecular Sciences. 2019; 20(17):4084. https://doi.org/10.3390/ijms20174084
Chicago/Turabian StyleGuo, Wei, Jing Lü, Mujuan Guo, Shimin Chen, Baoli Qiu, Wen Sang, Chunxiao Yang, Youjun Zhang, and Huipeng Pan. 2019. "De Novo Transcriptome Analysis Reveals Abundant Gonad-specific Genes in the Ovary and Testis of Henosepilachna vigintioctopunctata" International Journal of Molecular Sciences 20, no. 17: 4084. https://doi.org/10.3390/ijms20174084
APA StyleGuo, W., Lü, J., Guo, M., Chen, S., Qiu, B., Sang, W., Yang, C., Zhang, Y., & Pan, H. (2019). De Novo Transcriptome Analysis Reveals Abundant Gonad-specific Genes in the Ovary and Testis of Henosepilachna vigintioctopunctata. International Journal of Molecular Sciences, 20(17), 4084. https://doi.org/10.3390/ijms20174084