Extensive Chromosomal Reorganization in Apistogramma Fishes (Cichlidae, Cichlinae) Fits the Complex Evolutionary Diversification of the Genus


 ,
,
Abstract
1. Introduction
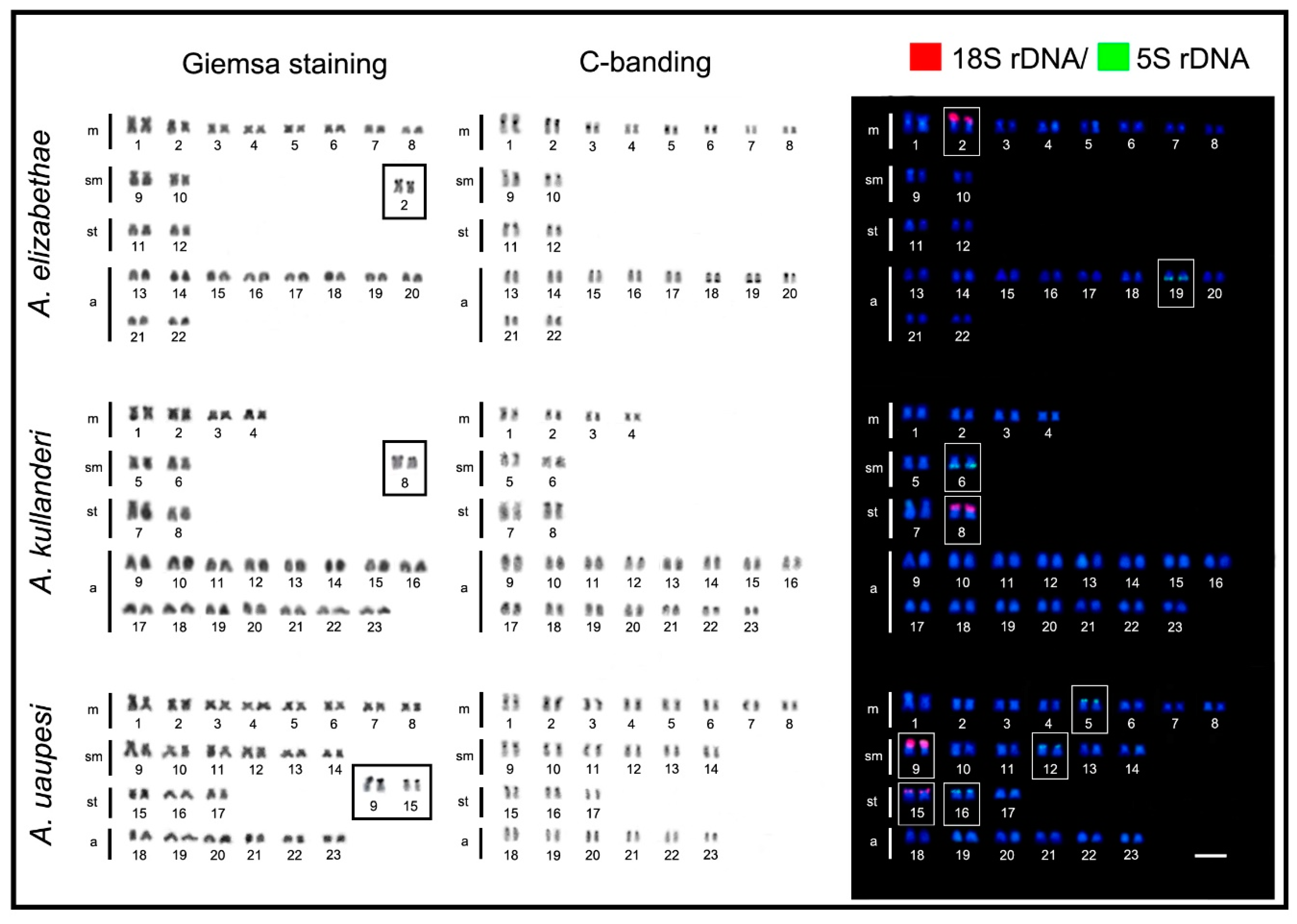
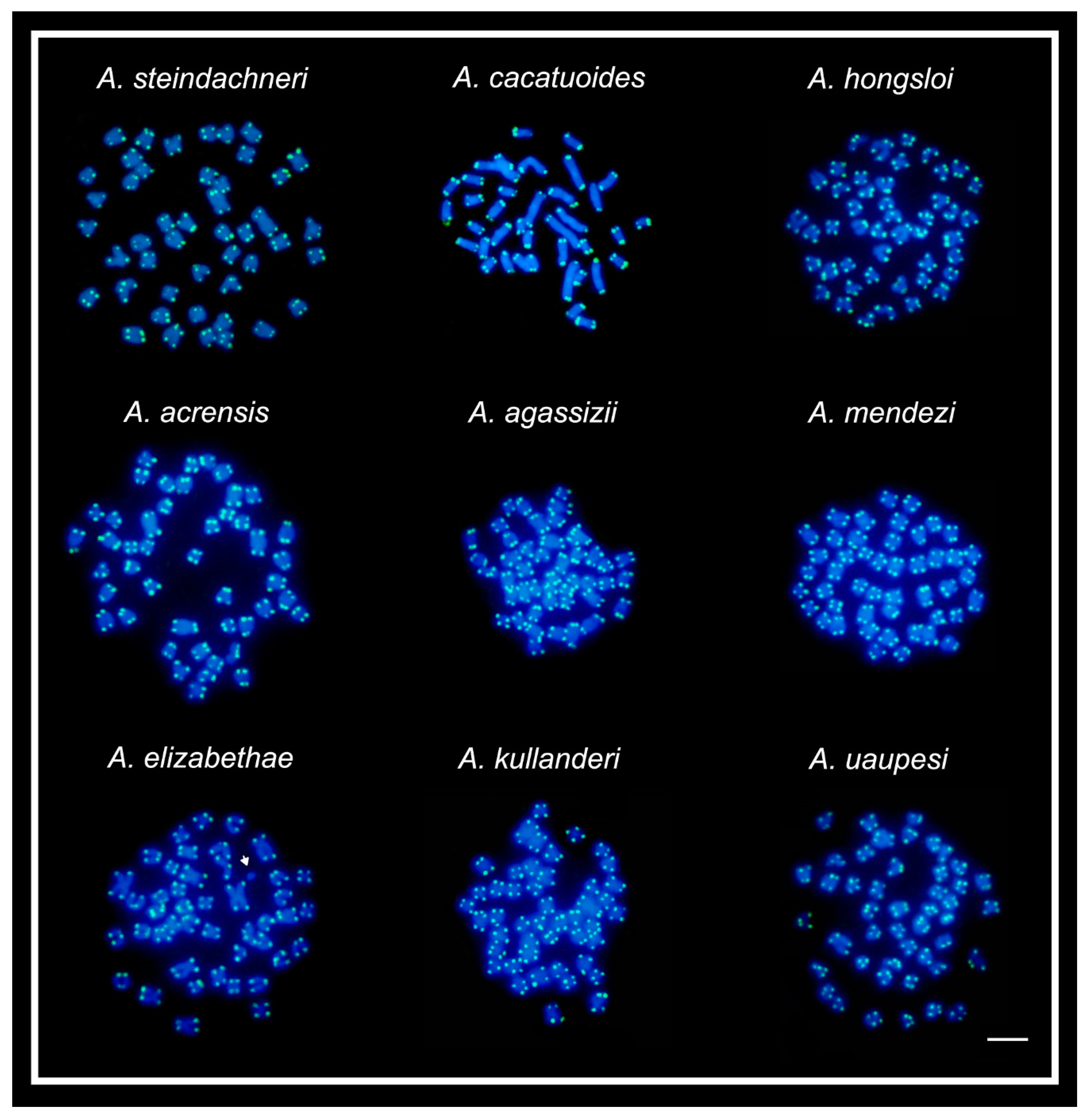
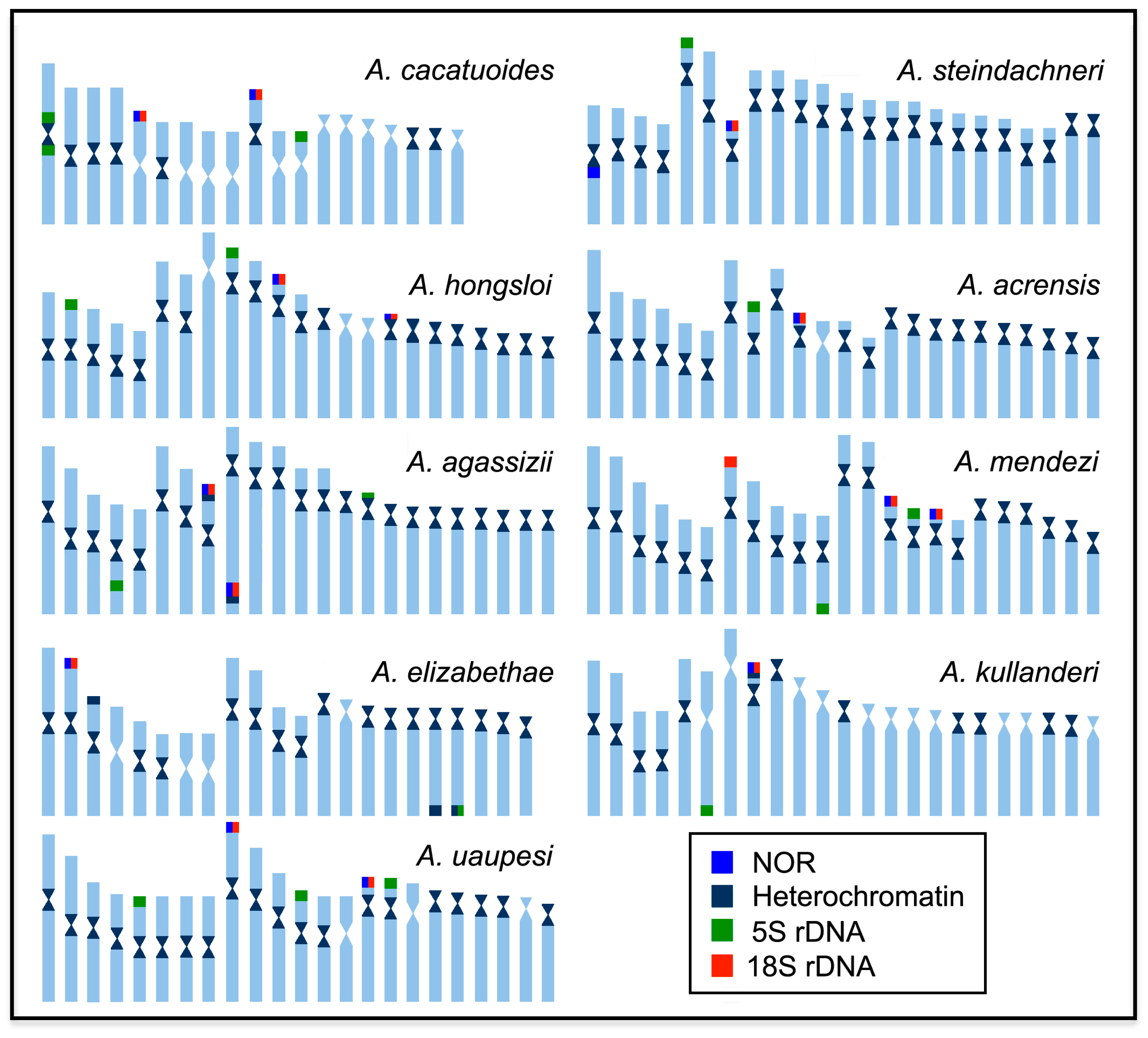
2. Results
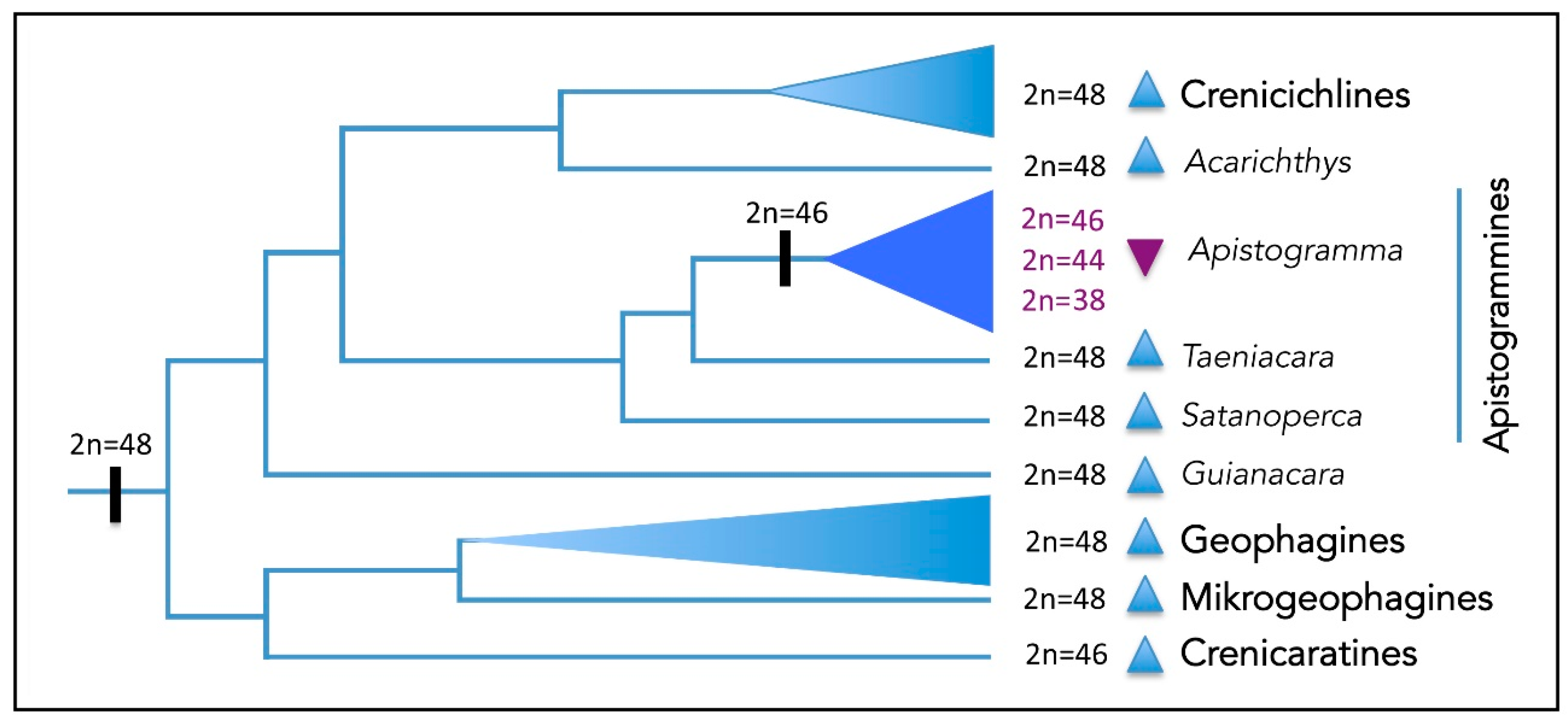
3. Discussion
3.1. Chromosomal Evolutionary Pathways in the Apistogramma genus
3.2. The Complex Chromosomal Diversification in Apistogramma Species Flock
3.3. Evolutionary Divergence in the Apistogramma genus. A General View
4. Materials and Methods
4.1. Individuals, Chromosomal Preparations, DNA Extraction
4.2. Conventional Chromosome Staining
4.3. Fluorescence In Situ Hybridization
4.4. Karyotype Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kocher, T.D. Adaptive evolution and explosive speciation: The cichlid fish model. Nat. Rev. Genet. 2004, 5, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Eschmeyer, W.N.; Fricke, R.; van der Laan, R. Catalog of Fishes: Genera, Species, References. Available online: http://www.calacademy.org/scientists/projects/catalog-of-fishes (accessed on 10 July 2019).
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.M.; Petry, P.L.; Rocha, L.A.R. Fish biodiversity and conservation in South America. J. Fish Biol. 2016, 89, 1–36. [Google Scholar] [CrossRef]
- Römer, U. Cichlid Atlas: Natural History of South American Dwarf Cichlids; Mergus-Publishers: Melle, Germany, 2006. [Google Scholar]
- Turner, G.F. Adaptive radiation of cichlid fish. Curr. Biol. 2007, 17, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Arbour, J.H.; López-Fernández, H. Continental cichlid radiations: Functional diversity reveals the role of changing ecological opportunity in the Neotropics. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160556. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández, H.; Winemiller, K.O.; Honeycutt, R.L. Multilocus phylogeny and rapid radiations in Neotropical cichlid fishes (Perciformes: Cichlidae: Cichlinae). Mol. Phylogenet. Evol. 2010, 55, 1070–1086. [Google Scholar] [CrossRef] [PubMed]
- López-Fernández, H.; Arbour, J.H.; Winemiller, K.O.; Honeycutt, R.L. Testing for ancient adaptive radiations in neotropical cichlid fishes. Evolution 2013, 67, 1321–1337. [Google Scholar] [CrossRef]
- Feldberg, E.; Porto, J.I.R.; Bertollo, L.A.C. Chromosomal changes and adaptation of cichlid fishes during evolution. In Fish Adaptation; Val, A.L., Kapoor, B.G., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2003; pp. 285–308. [Google Scholar]
- Ferreira, I.A.; Poletto, A.B.; Kocher, T.D.; Mota-Velasco, J.C.; Penman, D.J.; Martins, C. Chromosome evolution in African cichlid fish: Contributions from the physical mapping of repeated DNAs. Cytogenet. Genome Res. 2010, 129, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Poletto, A.B.; Ferreira, I.A.; Cabral-de-Mello, D.C.; Nakajima, R.T.; Mazzuchelli, J.; Ribeiro, H.B.; Venere, P.C.; Nirchio, M.; Kocher, T.D.; Martins, C. Chromosome differentiation patterns during cichlid fish evolution. BMC Genet. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Valente, G.T.; Mazzuchelli, J.; Ferreira, I.A.; Poletto, A.B.; Fantinatti, B.E.A.; Martins, C. Cytogenetic mapping of the retroelements Rex1, Rex3 and Rex6 among cichlid fish: New Insights on the chromosomal distribution of transposable elements. Cytogenet. Genome Res. 2011, 133, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Artoni, R.F.; Vicari, M.R.; Martins, C.; Feldberg, E. Chromosomal evolution of neotropical cichlids: The role of repetitive DNA sequences in the organization and structure of karyotype. Rev. Fish Biol. Fish. 2013, 23, 201–214. [Google Scholar] [CrossRef]
- Hodanova, L.; Kalous, L.; Musilova, Z. Comparative cytogenetics of Neotropical cichlid fishes (Nannacara, Ivanacara and Cleithracara) indicates evolutionary reduction of diploid chromosome numbers. Comp. Cytogenet. 2014, 8, 169–183. [Google Scholar] [PubMed]
- Astudillo-Clavijo, V.; Arbour, J.H.; López-Fernández, H. Selection towards different adaptive optima drove the early diversification of locomotor phenotypes in the radiation of Neotropical geophagine cichlids. BMC Evol. Biol. 2015, 15, 77. [Google Scholar] [CrossRef] [PubMed]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Species by Family/Subfamily. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog (accessed on 8 July 2019).
- Staeck, W.; Schindler, I. Apistogramma sororcula, a new dwarf cichlid. (Teleostei: Cichlidae) from the drainage of the rio. Guaporé in Bolivia and Brazil. Vertebr. Zool. 2016, 66, 141–150. [Google Scholar]
- Kullander, S.O. Check List of the Freshwater Fishes of South and Central America; Reis, R.E., Kullander, S.O., Ferraris, J.C.J., Eds.; Edipucrs: Porto Alegre, Brazil, 2003; pp. 605–654. [Google Scholar]
- Britzke, R.; Mehanna, M. Status taxonômico de Apistogramma Regan, 1911 e sua classificação. Bol. Da Soc. Bras. De Ictiol. 2010, 98, 9–16. [Google Scholar]
- Mesa, S.L.M.; Lasso, C.A. Revisión del género Apistogramma Regan 1913 (Perciformes, Cichlidae) en la cuenca del río Orinoco. In Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá D.C., Colombia, 2011; p. 192. [Google Scholar]
- Varella, H.R.; Sabaj Pérez, M.H. A titan among dwarfs–Apistogramma kullanderi, new species (Teleostei: Cichlidae). Ichthyol. Explor. Freshw. 2014, 25, 243–258. [Google Scholar]
- Steele, S.E.; López-Fernández, H. Body size diversity and frequency distributions of Neotropical cichlid fishes (Cichliformes: Cichlidae: Cichlinae). PLoS ONE 2014, 9, e106336. [Google Scholar] [CrossRef] [PubMed]
- Tougard, C.; García Dávila, C.R.; Römer, U.; Duponchelle, F.; Cerqueira, F.; Paradis, E.; Guinand, B.; Chávez, C.A.; Salas, V.; Quérouil, S.; et al. Tempo and rates of diversification in the South American cichlid genus Apistogramma (Teleostei: Perciformes: Cichlidae). PLoS ONE 2017, 12, e0182618. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Barton, N.H. Accumulating postzygotic isolation genes in parapatry: A new twist on chromosomal speciation. Evolution 2003, 57, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; O’Neill, R.J. Chromosomes, conflict, and epigenetics: Chromosomal speciation revisited. Annu. Rev. Genom. Hum. Genet. 2010, 11, 291–316. [Google Scholar] [CrossRef] [PubMed]
- Lukhtanov, V.A.; Dincă, V.; Talavera, G.; Vila, R. Unprecedented within-species chromosome number cline in the Wood White butterfly Leptidea sinapis and its significance for karyotype evolution and speciation. BMC Evol. Biol. 2011, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Rieseberg, L.H. Revisiting the impact of inversions in evolution: From population genetic markers to drivers of adaptive shifts and speciation? Annu. Rev. Ecol. Evol. Syst. 2008, 39, 21–42. [Google Scholar] [CrossRef] [PubMed]
- Römer, U.; Engelking, B.; Beisenherz, W. Genetically determined mate choice can be influenced by learning in Apistogramma cacatuoides Hoedeman, 1951 (Teleostei, Cichlidae). Vertebr. Zool. 2014, 64, 199–206. [Google Scholar]
- IUCN. Available online: https://www.iucnredlist.org (accessed on 8 July 2019).
- Cioffi, M.B.; Molina, W.F.; Artoni, R.F.; Bertollo, L.A.C. Chromosomes as tools for discovering biodiversity–the case of Erythrinidae fish family. In Recent Trends in Cytogenetic Studies–Methodologies and Applications; Tirunilai, P., Ed.; InTech Publisher: Rijeka, Croatia, 2012; pp. 125–146. [Google Scholar]
- Thompson, K.W. Cytotaxonomy of 41 species of Neotropical Cichlidae. Copeia 1979, 4, 679–691. [Google Scholar] [CrossRef]
- Roncati, H.A.; Pastori, M.C.; Fenocchio, A.S. Cytogenetic studies and evolutive considerations on fishes of the family Cichlidae (Perciformes) from Paraná River (Argentina). Cytologia 2007, 72, 379–384. [Google Scholar] [CrossRef][Green Version]
- Gross, M.C.; Schneider, C.H.; Valente, G.T.; Martins, C.; Feldberg, E. Variability of 18S rDNA locus among Symphysodon fishes: Chromosomal rearrangements. J. Fish Biol. 2010, 76, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Benzaquem, D.C.; Feldberg, E.; Porto, J.I.R.; Gross, M.C.; Zuanon, J.A.S. Cytotaxonomy and karyoevolution of the genus Crenicichla (Perciformes, Cichlidae). Genet. Mol. Biol. 2008, 31, 250–255. [Google Scholar] [CrossRef]
- Salzburger, W.; Meyer, A.; Baric, S.; Verheyen, E.; Sturmbauer, C.; Westneat, M.W. Phylogeny of the Lake Tanganyika cichlid species flock and its relationship to the Central and East African Haplochromine cichlid fish faunas. Syst. Biol. 2002, 51, 113–135. [Google Scholar] [CrossRef]
- Piálek, L.; Ríčan, O.; Casciotta, J.; Almirón, A.; Zrzavý, J. Multilocus phylogeny of Crenicichla (Teleostei: Cichlidae), with biogeography of the C. lacustris group: Species flocks as a model for sympatric speciation in rivers. Mol. Phylogenetics Evol. 2012, 62, 46–61. [Google Scholar] [CrossRef]
- Perazzo, G.X.; Noleto, R.B.; Vicari, M.R.; Gava, A.; Cestari, M.M. Trends of karyotypical evolution in the pearl cichlid, Geophagus brasiliensis, from southern Brazil. Zoology 2013, 116, 286–292. [Google Scholar] [CrossRef]
- Getlekha, N.; Molina, W.F.; Cioffi, M.B.; Yano, C.F.; Maneechot, N.; Bertollo, L.A.C.; Supiwong, W.; Tanomtong, A. Repetitive DNAs highlight the role of chromosomal fusions in the karyotype evolution of Dascyllus species (Pomacentridae, Perciformes). Genetica 2016, 144, 203–211. [Google Scholar] [CrossRef]
- Vicari, M.R.; Noleto, R.B.; Artoni, R.F.; Moreira-Filho, O.; Bertollo, L.A.C. Comparative cytogenetics among species of the Astyanax scabripinnis complex: Evolutionary and biogeographical inferences. Genet. Mol. Biol. 2008, 31, 173–179. [Google Scholar] [CrossRef]
- Greenwood, P.H. The cichlid fishes of Lake Victoria, East Africa: The biology and evolution of a species flock. Bull. Br. Mus. 1974, 6, 1–134. [Google Scholar]
- Lecointre, G.; Améziane, N.; Boisselier, M.C.; Bonillo, C.; Busson, F.; Causse, R.; Chenuil, A.; Couloux, A.; Coutanceau, J.P.; Cruaud, C.; et al. Is the species flock concept operational? The Antarctic shelf case. PLoS ONE 2013, 8, e68787. [Google Scholar] [CrossRef] [PubMed]
- Varella, H.R.; Britzke, R. Apistogramma eleutheria and A. flavipedunculata, two new species of dwarf cichlids from the rio Curuá on Serra do Cachimbo, Brazil (Teleostei: Cichlidae). Ichthyol. Explor. Freshw. 2016, 27, 81–95. [Google Scholar]
- Cioffi, M.B.; Kejnovskyý, E.; Marquioni, V.; Poltronieri, J.; Molina, W.F.; Diniz, D.; Bertollo, L.A.C. The key role of repeated DNAs in sex chromosome evolution in two fish species with ZW sex chromosome system. Mol. Cytogenet. 2012, 5, 28. [Google Scholar] [CrossRef]
- Lima-Filho, P.A.; Bertollo, L.A.C.; Cioffi, M.B.; Costa, G.W.W.F.; Molina, W.F. Karyotype divergence and spreading of 5S rDNA sequences between genomes of two species: Darter and emerald gobies (Ctenogobius, Gobiidae). Cytogenet. Genome Res. 2014, 142, 197–203. [Google Scholar] [CrossRef]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [PubMed]
- Supiwong, W.; Pinthong, K.; Seetapan, K.; Saenjundaeng, P.; Bertollo, L.A.C.; de Oliveira, E.A.; Yano, C.F.; Liehr, T.; Phimphan, S.; Tanomtong, A.; et al. Karyotype diversity and evolutionary trends in the Asian swamp eel Monopterus albus (Synbranchiformes, Synbranchidae): A case of chromosomal speciation? BMC Evol. Biol. 2019, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Feldberg, E.; Bertollo, L.A.C. Nucleolar organizing regions in some species of Neotropical cichlid fish (Pisces, Perciformes). Caryologia 1985, 38, 319–324. [Google Scholar] [CrossRef]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Pires, L.B.; Giuliano-Caetano, L.; Dias, A.L. Cytogenetic characterization of Geophagus brasiliensis and two species of Gymnogeophagus (Cichlidae: Geophaginae) from Guaiba Lake, RS, Brazil. Folia Biol. 2010, 58, 29–34. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. 2018. Available online: www.fishbase.org (acessed on 10 February 2019).
- Prieto, J.L.; McStay, B. Pseudo-NORs: A novel model for studying nucleoli. Biochim. Biophys. Acta 2008, 1783, 2116–2123. [Google Scholar] [CrossRef]
- Motta-Neto, C.C.; Marques, A.; Costa, G.W.W.F.; Cioffi, M.B.; Bertollo, L.A.C.; Soares, R.X.; Scortecci, K.C.; Artoni, R.F.; Molina, W.F. Differential hypomethylation of the repetitive Tol2/Alu-rich sequences in the genome of Bodianus species (Labriformes, Labridae). Comp. Cytogenet. 2018, 12, 145–162. [Google Scholar] [CrossRef] [PubMed]
- Robicheau, B.M.; Susko, E.; Harrigan, A.M.; Snyder, M. Ribosomal RNA genes contribute to the formation of pseudogenes and junk DNA in the human genome. Genome Biol. Evol. 2017, 9, 380–397. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Srivastava, R.; Ahn, S.H. The epigenetic pathways to ribosomal DNA silencing. Microbiol. Mol. Biol. Rev. 2016, 80, 545–563. [Google Scholar] [PubMed]
- Martins, I.C.; Portela-Castro, A.L.B.; Júlio Júnior, H.F. Chromosome analysis of 5 species of the Cichlidae family (Pisces-Perciformes) from the Parana River. Cytologia 1995, 60, 223–231. [Google Scholar] [CrossRef]
- Mendonça, M.N.C.; Porto, J.I.R.; Feldberg, E. Ocorrência de três citótipos em Satanoperca aff. jurupari (Perciformes, Cichlidae) no Catalão, Manaus, AM. Genet. Mol. Biol. 1999, 22, 69. [Google Scholar]
- Silva, F.A.; Carvalho, N.D.; Schneider, C.H.; Terencio, M.L.; Feldberg, E.; Gross, M.C. Comparative cytotaxonomy of two species of fish from the genus Satanoperca reveals the presence of a B chromosome. Zebrafish 2016, 13, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Molina, W.F.; Galetti, P.M., Jr. Robertsonian rearrangements in the reef fish Chromis (Perciformes, Pomacentridae) involving chromosomes bearing 5S rRNA genes. Genet. Mol. Biol. 2002, 25, 373–377. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Chromosome spreading of associated transposable elements and ribosomal DNA in the fish Erythrinus erythrinus. Implications for genome change and karyoevolution in fish. BMC Evol. Biol. 2010, 10, 271. [Google Scholar] [CrossRef]
- Oliveira, E.A.; Sember, A.; Bertollo, L.A.C.; Yano, C.F.; Ezaz, T.; Moreira Filho, O.; Hatanaka, T.; Trifonov, V.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Tracking the evolutionary pathway of sex chromosomes among fishes: Characterizing the unique XX/XY1Y2 system in Hoplias malabaricus (Teleostei, Characiformes). Chromosoma 2018, 127, 115. [Google Scholar] [CrossRef] [PubMed]
- Clark, F.E.; Conte, M.A.; Kocher, T.D. Genomic characterization of a B chromosome in Lake Malawi cichlid fishes. Genes 2018, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Perazzo, G.X.; Noleto, R.B.; Vicari, M.R.; Gava, A.; Cestari, M.M. B chromosome polymorphism in South American cichlid. Neotrop. Biodivers. 2018, 4, 3–9. [Google Scholar] [CrossRef]
- Ready, J.S.; Sampaio, I.; Schneider, H.; Vinson, C.; Santos, T.; Turner, G.F. Colour forms of Amazonian cichlid fish represent reproductively isolated species. J. Evol. Biol. 2006, 19, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Engelking, B.; Römer, U.; Beisenherz, W. The intraspecific colour preference in mate choice by female Apistogramma cacatuoides Hoedeman, 1951 (Teleostei: Perciformes: Cichlidae). Vertebr. Zool. 2010, 60, 9–18. [Google Scholar]
- Seehausen, O. Conservation: Losing biodiversity by reverse speciation. Curr. Biol. 2006, 16, 334–337. [Google Scholar] [CrossRef]
- Price, T.D.; Bouvier, M.M. The evolution of F1 postzygotic incompatibilities in birds. Evolution 2002, 56, 2083–2089. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.T. Evolution of intrinsic post-zygotic reproductive isolation in fish. Ann. Zool. Fenn. 2003, 40, 321–329. [Google Scholar]
- Molina, W.F.; Alves, D.E.O.; Araujo, W.C.; Martinez, P.A.; Silva, M.F.M.; Costa, G.W.W.F. Performance of human immunostimulating agents in the improvement of fish cytogenetic preparations. Genet. Mol. Res. 2010, 9, 1807–1814. [Google Scholar] [CrossRef]
- Gold, J.R.; Li, Y.C.; Shipley, N.S.; Powers, P.K. Improved methods for working with fish chromosomes with a review of metaphase chromosome banding. J. Fish Biol. 1990, 37, 563–575. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Howell, W.M.; Black, D.A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef] [PubMed]
- Sumner, A.T. Simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Pinkel, D.; Straume, T.; Gray, J.W. Cytogenetic analysis using quantitative, high-sensitivity, fluorescence hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 2934–2938. [Google Scholar] [CrossRef] [PubMed]
- Pendás, A.M.; Moran, P.; Freije, J.P.; Garcia-Vazquez, E. Chromosomal mapping and nucleotide sequence of two tandem repeats of Atlantic salmon 5S rDNA. Cytogenet. Cell Genet. 1994, 67, 31–36. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
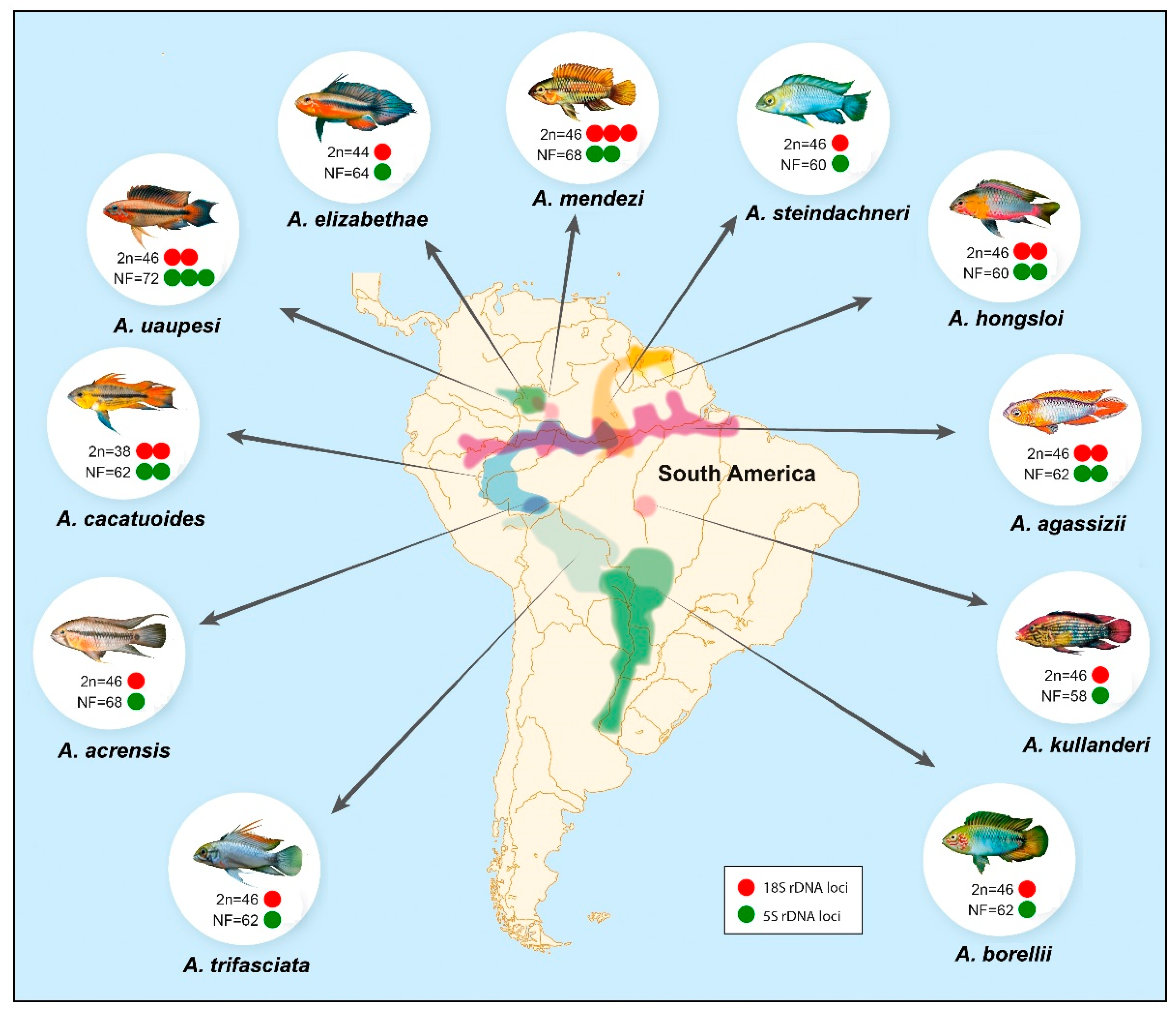{kind=link}
| Species | 2n | Karyotype Formula | NF | 18S rDNA | 5S rDNA | Ref. |
|---|---|---|---|---|---|---|
| A. acrensis | 46 | 12m + 4sm + 6st + 24a | 62 | Unique (st-T) | Unique (sm-T) | * |
| A. agassizii | 46 | 12m + 4sm + 8st + 22a | 62 | Multiple—2 pairs (sm/st-T) | Multiple—2 pairs (m/a-T) | * |
| A. agassizii | 46 | 24m/sm + 22st/a | 70 | - | - | [31] |
| A. borellii | 46 | 16m/sm + 30st/a | 62 | - | - | [11] |
| A. cacatuoides | 38 | 18m + 6sm + 14a | 62 | Multiple—2 pairs (m/sm-T) | Multiple—2 pairs (m-I, st-T) | * |
| A. elizabethae | 44 | 16m + 4sm + 4st + 20a | 64 | Unique (m-T) | Unique—(a-T) | * |
| A. hongsloi | 46 | 10m + 4sm + 10st + 22a | 60 | Multiple—2 pairs (m/sm-T) | Multiple—2 pairs (st/a-T) | * |
| A. kullanderi | 46 | 8m + 4sm + 4st + 30a | 58 | Unique (st-T) | Unique (sm-T) | * |
| A. mendezi | 46 | 12m + 10sm + 10st + 14a | 68 | Multiple—3 pairs (1sm/2a-T) | Multiple—2 pairs (m/sm-T) | * |
| A. ortmanni | 46 | 24m/sm + 22st/a | 70 | - | - | [31] |
| A. steindachneri | 46 | 8m + 6sm + 28st + 4a | 60 | Unique (st-T) | Unique (sm-T) | * |
| A. trifasciata | 46 | 16m/sm + 30st/a | 62 | - | - | [32] |
| A. uaupesi | 46 | 18m + 8sm + 6st + 14a | 72 | Multiple—2 pairs (sm/st-T) | Multiple—3 pairs (m/sm/st-T) | * |
| Species Complex | Species | Geographic Distribution | N | Sex |
|---|---|---|---|---|
| A. cacatuoides | A. cacatuoides Hoedeman, 1951 | E | 6 | 3 ♂, 2 ♀, 1 immature |
| A. steindachneri | A. steindachneri Regan, 1908 | E | 5 | 2 ♂, 3 ♀ |
| A. agassizii | A. agassizii (Steindachner, 1875) | E | 3 | 2 ♂, 1 ♀ |
| A. agassizii | A. elizabethae Kullander, 1980 | R | 2 | 2 ♂ |
| A. agassizii | A. mendezi Römer, 1994 | R | 2 | 1 ♂, 1 ♀ |
| A. regani? | A. kullanderi Varella and Sabaj Pérez, 2014 | VR | 1 | 1 ♀ |
| A. regani | A. acrensis Staeck, 2003 | R | 3 | 2 ♂, 1 ♀ |
| A. pertensis | A. uaupesi Kullander, 1980 | R | 2 | 2 ♂ |
| A. diplotaenia | A. diplotaenia | R | 3 | 2 ♂, 1 ♀ |
| A. macmasteri | A. hongsloi | R | 2 | 2 ♀ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner Werneck Félix da Costa, G.; de Bello Cioffi, M.; Liehr, T.; Feldberg, E.; Antonio Carlos Bertollo, L.; Franco Molina, W. Extensive Chromosomal Reorganization in Apistogramma Fishes (Cichlidae, Cichlinae) Fits the Complex Evolutionary Diversification of the Genus. Int. J. Mol. Sci. 2019, 20, 4077. https://doi.org/10.3390/ijms20174077
Wagner Werneck Félix da Costa G, de Bello Cioffi M, Liehr T, Feldberg E, Antonio Carlos Bertollo L, Franco Molina W. Extensive Chromosomal Reorganization in Apistogramma Fishes (Cichlidae, Cichlinae) Fits the Complex Evolutionary Diversification of the Genus. International Journal of Molecular Sciences. 2019; 20(17):4077. https://doi.org/10.3390/ijms20174077
Chicago/Turabian StyleWagner Werneck Félix da Costa, Gideão, Marcelo de Bello Cioffi, Thomas Liehr, Eliana Feldberg, Luiz Antonio Carlos Bertollo, and Wagner Franco Molina. 2019. "Extensive Chromosomal Reorganization in Apistogramma Fishes (Cichlidae, Cichlinae) Fits the Complex Evolutionary Diversification of the Genus" International Journal of Molecular Sciences 20, no. 17: 4077. https://doi.org/10.3390/ijms20174077
APA StyleWagner Werneck Félix da Costa, G., de Bello Cioffi, M., Liehr, T., Feldberg, E., Antonio Carlos Bertollo, L., & Franco Molina, W. (2019). Extensive Chromosomal Reorganization in Apistogramma Fishes (Cichlidae, Cichlinae) Fits the Complex Evolutionary Diversification of the Genus. International Journal of Molecular Sciences, 20(17), 4077. https://doi.org/10.3390/ijms20174077