Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood–Brain Barrier Alterations after Cerebral Photothrombosis in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. MLR-HFS Does Not Change BBB Leakage and Albumin Extravasation
2.2. MLR-HFS Does Not Influence Degradation of Tight Junction Molecules
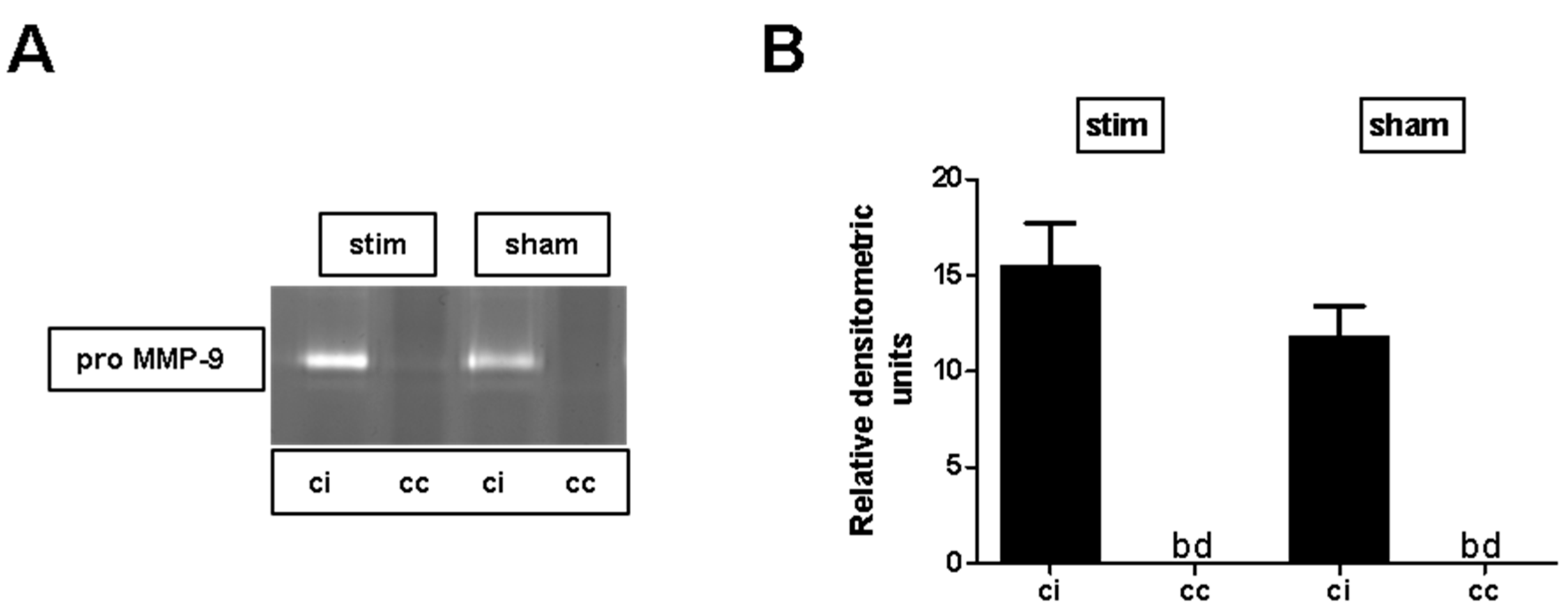
2.3. MLR-HFS Does Not Affect Matrix Metalloproteinase-9 Expression after Photothrombosis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of Photothrombotic Stroke
4.3. Microelectrode Implantation
4.4. High-Frequency Stimulation of the Mesencephalic Locomotor Region
4.5. Collection of Cerebral Tissue
4.6. Immunohistochemistry
4.7. Western Blot Assays
4.8. Zymography
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lo, E.H.; Dalkara, T.; Moskowitz, M.A. Neurological diseases: Mechanisms, challenges and opportunities in stroke. Nat. Rev. Neurosci. 2003, 4, 399. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Rosenberg, G.A. Blood-Brain Barrier Breakdown in Acute and Chronic Cerebrovascular Disease. Stroke 2011, 42, 3323–3328. [Google Scholar] [CrossRef] [PubMed]
- Hjort, N.; Wu, O.; Ashkanian, M.; Solling, C.; Mouridsen, K.; Christensen, S.; Gyldensted, C.; Andersen, G.; Ostergaard, L. MRI Detection of Early Blood-Brain Barrier Disruption: Parenchymal Enhancement Predicts Focal Hemorrhagic Transformation After Thrombolysis. Stroke 2008, 39, 1025–1028. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The Blood-Brain Barrier in Health and Chronic Neurodegenerative Disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Stoll, G.; Kleinschnitz, C.; Meuth, S.G.; Braeuninger, S.; Ip, C.W.; Wessig, C.; Nölte, I.; Bendszus, M. Transient widespread blood–brain barrier alterations after cerebral photothrombosis as revealed by gadofluorine M-enhanced magnetic resonance imaging. J. Cereb. Blood Flow Metab. 2008, 29, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Simpkins, A.N.; Dias, C.; Leigh, R. Identification of Reversible Disruption of the Human Blood–Brain Barrier Following Acute Ischemia. Stroke 2016, 47, 2405–2408. [Google Scholar] [CrossRef]
- Levi, H.; Schoknecht, K.; Prager, O.; Chassidim, Y.; Weissberg, I.; Serlin, Y.; Friedman, A. Stimulation of the Sphenopalatine Ganglion Induces Reperfusion and Blood-Brain Barrier Protection in the Photothrombotic Stroke Model. PLoS ONE 2012, 7, e39636. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Zhu, G.-Y.; Wang, X.; Shi, L.; Du, T.-T.; Liu, D.-F.; Liu, Y.-Y.; Jiang, Y.; Zhang, X.; Zhang, J.-G. Anterior thalamic nuclei deep brain stimulation reduces disruption of the blood–brain barrier, albumin extravasation, inflammation and apoptosis in kainic acid-induced epileptic rats. Neurol. Res. 2017, 39, 1103–1113. [Google Scholar] [CrossRef]
- Fluri, F.; Malzahn, U.; Homola, G.A.; Schuhmann, M.K.; Kleinschnitz, C.; Volkmann, J. Stimulation of the mesencephalic locomotor region for gait recovery after stroke: DBS for Gait Recovery. Ann. Neurol. 2017, 82, 828–840. [Google Scholar] [CrossRef]
- Schuhmann, M.K.; Stoll, G.; Bohr, A.; Volkmann, J.; Fluri, F. Electrical Stimulation of the Mesencephalic Locomotor Region Attenuates Neuronal Loss and Cytokine Expression in the Perifocal Region of Photothrombotic Stroke in Rats. Int. J. Mol. Sci. 2019, 20, 2341. [Google Scholar] [CrossRef]
- Ryczko, D.; Dubuc, R. The Multifunctional Mesencephalic Locomotor Region. Curr. Pharm. Des. 2013, 19, 4448–4470. [Google Scholar] [CrossRef] [PubMed]
- Rayasam, A.; Hsu, M.; Hernández, G.; Kijak, J.; Lindstedt, A.; Gerhart, C.; Sandor, M.; Fabry, Z. Contrasting roles of immune cells in tissue injury and repair in stroke: The dark and bright side of immunity in the brain. Neurochem. Int. 2017, 107, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Petrovic-Djergovic, D.; Goonewardena, S.N.; Pinsky, D.J. Inflammatory Disequilibrium in Stroke. Circ. Res. 2016, 119, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Kooij, G.; Kopplin, K.; Blasig, R.; Stuiver, M.; Koning, N.; Goverse, G.; van der Pol, S.M.A.; van Het Hof, B.; Gollasch, M.; Drexhage, J.A.R.; et al. Disturbed function of the blood-cerebrospinal fluid barrier aggravates neuro-inflammation. Acta Neuropathol. 2014, 128, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Castro Dias, M.; Coisne, C.; Lazarevic, I.; Baden, P.; Hata, M.; Iwamoto, N.; Francisco, D.M.F.; Vanlandewijck, M.; He, L.; Baier, F.A.; et al. Claudin-3-deficient C57BL/6J mice display intact brain barriers. Sci. Rep. 2019, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Pinsky, D.J.; Naka, Y.; Liao, H.; Oz, M.C.; Wagner, D.D.; Mayadas, T.N.; Johnson, R.C.; Hynes, R.O.; Heath, M.; Lawson, C.A.; et al. Hypoxia-induced exocytosis of endothelial cell Weibel-Palade bodies. A mechanism for rapid neutrophil recruitment after cardiac preservation. J. Clin. Invest. 1996, 97, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Mojiri, A.; Nakhaii-Nejad, M.; Phan, W.-L.; Kulak, S.; Radziwon-Balicka, A.; Jurasz, P.; Michelakis, E.; Jahroudi, N. Hypoxia Results in Upregulation and De Novo Activation of Von Willebrand Factor Expression in Lung Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Yarnitsky, D.; Gross, Y.; Lorian, A.; Shalev, A.; Lamensdorf, I.; Bornstein, R.; Shorer, S.; Mayevsky, A.; Patel, K.P.; Abbott, N.J.; et al. Blood—brain barrier opened by stimulation of the parasympathetic sphenopalatine ganglion: A new method for macromolecule delivery to the brain. J. Neurosurg. 2004, 101, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Hardebo, J.E.; Skagerberg, G.; Owman, C. Central origins of preganglionic fibers to the sphenopalatine ganglion in the rat. A fluorescent retrograde tracer study with special reference to its relation to central catecholaminergic systems. J. Auton. Nerv. Syst. 1990, 30, 101–109. [Google Scholar] [CrossRef]
- Skinner, R.D.; Garcia-Rill, E. The mesencephalic locomotor region (MLR) in the rat. Brain Res. 1984, 323, 385–389. [Google Scholar] [CrossRef]
- Liang, N.; Mitchell, J.H.; Smith, S.A.; Mizuno, M. Exaggerated sympathetic and cardiovascular responses to stimulation of the mesencephalic locomotor region in spontaneously hypertensive rats. Am. J. Physiol.-Heart Circ. Physiol. 2016, 310, H123–H131. [Google Scholar] [CrossRef] [PubMed]
- Schuhmann, M.K.; Kraft, P.; Stoll, G.; Lorenz, K.; Meuth, S.G.; Wiendl, H.; Nieswandt, B.; Sparwasser, T.; Beyersdorf, N.; Kerkau, T.; et al. CD28 superagonist-mediated boost of regulatory T cells increases thrombo-inflammation and ischemic neurodegeneration during the acute phase of experimental stroke. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2015, 35, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Schuhmann, M.K.; Fluri, F. Effects of Fullerenols on Mouse Brain Microvascular Endothelial Cells. Int. J. Mol. Sci. 2017, 18, 1783. [Google Scholar] [CrossRef] [PubMed]
- Schuhmann, M.; Bittner, S.; Meuth, S.; Kleinschnitz, C.; Fluri, F. Fingolimod (FTY720-P) Does Not Stabilize the Blood–Brain Barrier under Inflammatory Conditions in an in Vitro Model. Int. J. Mol. Sci. 2015, 16, 29454–29466. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuhmann, M.K.; Stoll, G.; Papp, L.; Bohr, A.; Volkmann, J.; Fluri, F. Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood–Brain Barrier Alterations after Cerebral Photothrombosis in Rats. Int. J. Mol. Sci. 2019, 20, 4036. https://doi.org/10.3390/ijms20164036
Schuhmann MK, Stoll G, Papp L, Bohr A, Volkmann J, Fluri F. Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood–Brain Barrier Alterations after Cerebral Photothrombosis in Rats. International Journal of Molecular Sciences. 2019; 20(16):4036. https://doi.org/10.3390/ijms20164036
Chicago/Turabian StyleSchuhmann, Michael K., Guido Stoll, Lena Papp, Arne Bohr, Jens Volkmann, and Felix Fluri. 2019. "Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood–Brain Barrier Alterations after Cerebral Photothrombosis in Rats" International Journal of Molecular Sciences 20, no. 16: 4036. https://doi.org/10.3390/ijms20164036
APA StyleSchuhmann, M. K., Stoll, G., Papp, L., Bohr, A., Volkmann, J., & Fluri, F. (2019). Electrical Stimulation of the Mesencephalic Locomotor Region Has No Impact on Blood–Brain Barrier Alterations after Cerebral Photothrombosis in Rats. International Journal of Molecular Sciences, 20(16), 4036. https://doi.org/10.3390/ijms20164036