Feeding Stimulates Sphingosine-1-Phosphate Mobilization in Mouse Hypothalamus


Abstract
1. Introduction
2. Results
2.1. Feeding Increases SA1P and S1P Levels in Mouse Hypothalamus
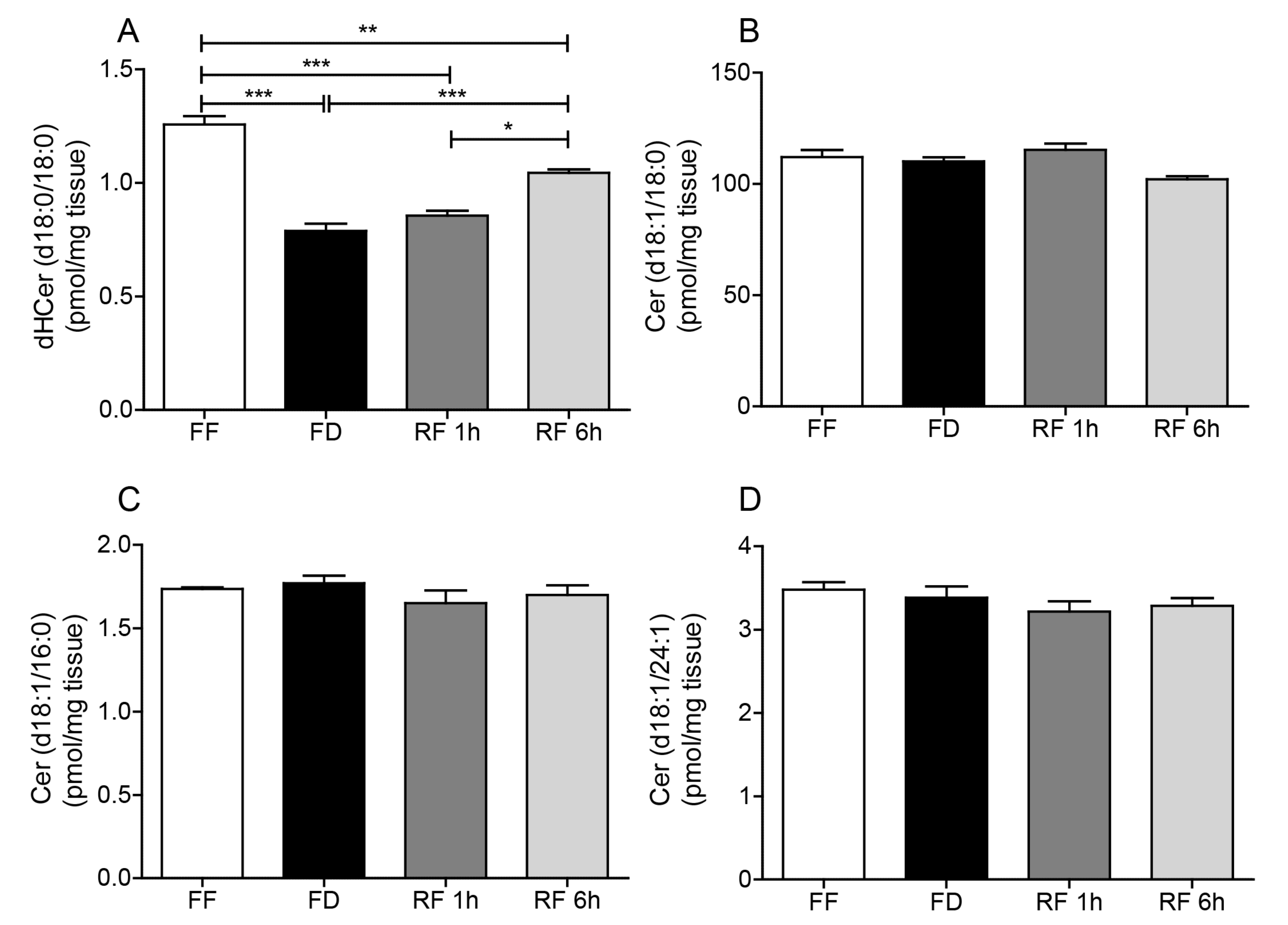
2.2. Effects of Feeding On Hypothalamic Sphingolipid Metabolism
2.3. Effects of Feeding Status on Expression of Sphingolipid-Metabolizing Enzymes
2.4. Feeding Regulates S1pr1 Transcription in Hypothalamus
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Diet
4.3. Chemicals
4.4. Experimental Design: Effects of Food Deprivation and Refeeding
4.5. Tissue Collection
4.6. Lipid Extraction
4.7. LC/MS-MS Analyses
4.8. mRNA Isolation, cDNA Synthesis, and Quantitative Real-time PCR
4.9. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| S1P | sphingosine-1-phosphate |
| SA1P | sphinganine-1-phosphate |
| S1PR1 | sphingosine-1-phosphate receptor 1 |
| SO | sphingosine |
| SA | sphinganine |
| Sptlc2 | serine palmitoyltransferase long chain base subunit 2 |
| Lass1 | longevity assurance gene 1 |
| Sphk1 | sphingosine kinase 1 |
| Sphk2 | sphingosine kinase 2 |
| AgRP | agouti-related protein |
| α-MSH | α-melanocyte-stimulating hormone |
| LC/MS | liquid chromatography/mass spectrometry |
| FF | free feeding |
| FD | food deprivation |
| RF 1h | 1 h refeeding |
| RF 6h | 6 h refeeding |
| TFA | trifluoroacetic acid |
| MRM | multiple reaction monitoring |
| SEM | standard error of the mean |
| Gapdh | glyceraldehyde 3-phosphate dehydrogenase |
| Hprt | hypoxanthine phosphoribosyltransferase |
References
- Morton, G.J.; Cummings, D.E.; Baskin, D.G.; Barsh, G.S.; Schwartz, M.W. Central nervous system control of food intake and body weight. Nature 2006, 443, 289. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Lee, D.K.; Jo, Y.H. Cholinergic neurons in the dorsomedial hypothalamus regulate food intake. Mol. Metab. 2017, 6, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Koch, M.; Horvath, T.L. Molecular and cellular regulation of hypothalamic melanocortin neurons controlling food intake and energy metabolism. Mol. Psychiatry 2014, 19, 752. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.H.; Lee, D.K.; Liu, S.M.; Chua, S.C.; Schwartz, G.J.; Jo, Y.H. Activation of temperature-sensitive TRPV1-like receptors in ARC POMC neurons reduces food intake. PLoS Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Crespo, C.S.; Cachero, A.P.; Jiménez, L.P.; Barrios, V.; Ferreiro, E.A. Peptides and food intake. Front. Endocrinol. (Lausanne). 2014, 16, e2004399. [Google Scholar]
- Krashes, M.J.; Koda, S.; Ye, C.P.; Rogan, S.C.; Adams, A.C.; Cusher, D.S.; Maratos-Flier, E.; Roth, B.L.; Lowell, B.B. Rapid, reversible activation of AgRP neurons drives feeding behavior in mice. J. Clin. Invest. 2011, 121, 1424–1428. [Google Scholar] [CrossRef] [PubMed]
- D’agostino, G.; Diano, S. Alpha-melanocyte stimulating hormone: Production and degradation. J. Mol. Med. 2010, 88, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175. [Google Scholar] [CrossRef]
- Silva, V.R.R.; Micheletti, T.O.; Pimentel, G.D.; Katashima, C.K.; Lenhare, L.; Morari, J.; Mendes, M.C.S.; Razolli, D.S.; Rocha, G.Z.; de Souza, C.T.; et al. Hypothalamic S1P/S1PR1 axis controls energy homeostasis. Nat. Commun. 2014, 5, 4859. [Google Scholar] [CrossRef]
- Hanada, K. Serine palmitoyltransferase, a key enzyme of sphingolipid metabolism. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2003, 1632, 16–30. [Google Scholar] [CrossRef]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef]
- Liu, H.; Sugiura, M.; Nava, V.E.; Edsall, L.C.; Kono, K.; Poulton, S.; Milstien, S.; Kohama, T.; Spiegel, S. Molecular cloning and functional characterization of a novel mammalian sphingosine kinase type 2 isoform. J. Biol. Chem. 2000, 275, 19513–19520. [Google Scholar] [CrossRef]
- Bryan, L.; Kordula, T.; Spiegel, S.; Milstien, S. Regulation and functions of sphingosine kinases in the brain. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2008, 1781, 459–466. [Google Scholar] [CrossRef]
- Park, S.W.; Kim, M.; Chen, S.W.C.; Brown, K.M.; D’Agati, V.D.; Lee, H.T. Sphinganine-1-phosphate protects kidney and liver after hepatic ischemia and reperfusion in mice through S1P 1 receptor activation. Lab. Investig. 2010, 90, 1209. [Google Scholar] [CrossRef]
- Delgado, A.; Martínez-Cartro, M. Therapeutic Potential of the Modulation of Sphingosine-1-Phosphate Receptors. Curr. Med. Chem. 2016, 23, 242–264. [Google Scholar] [CrossRef]
- Gillard, B.K.; Clement, R.G.; Marcus, D.M. Variations among cell lines in the synthesis of sphingolipids in de novo and recycling pathways. Glycobiology 1998, 8, 885–890. [Google Scholar] [CrossRef]
- Marchesini, N.; Hannun, Y.A. Acid and neutral sphingomyelinases: Roles and mechanisms of regulation. Biochem. Cell Biol. 2004, 82, 27–44. [Google Scholar] [CrossRef]
- Tettamanti, G. Ganglioside/glycosphingolipid turnover: New concepts. Glycoconj. J. 2003, 20, 301–317. [Google Scholar] [CrossRef]
- Picard, A.; Moullé, V.S.; Le Foll, C.; Cansell, C.; Véret, J.; Coant, N.; Le Stunff, H.; Migrenne, S.; Luquet, S.; Cruciani-Guglielmacci, C.; et al. Physiological and pathophysiological implications of lipid sensing in the brain. Diabetes Obes. Metab. 2014, 16, 49–55. [Google Scholar] [CrossRef]
- Le Stunff, H.; Coant, N.; Migrenne, S.; Magnan, C. Targeting lipid sensing in the central nervous system: New therapy against the development of obesity and type 2 diabetes. Expert Opin. Ther. Targets 2013, 17, 545–555. [Google Scholar] [CrossRef]
- Cruciani-Guglielmacci, C.; López, M.; Campana, M.; le Stunff, H. Brain ceramide metabolism in the control of energy balance. Front. Physiol. 2017, 8, 787. [Google Scholar] [CrossRef]
- Campana, M.; Bellini, L.; Rouch, C.; Rachdi, L.; Coant, N.; Butin, N.; Bandet, C.L.; Philippe, E.; Meneyrol, K.; Kassis, N.; et al. Inhibition of central de novo ceramide synthesis restores insulin signaling in hypothalamus and enhances β-cell function of obese Zucker rats. Mol. Metab. 2018, 8, 23–36. [Google Scholar] [CrossRef]
- Contreras, C.; González-García, I.; Martínez-Sánchez, N.; Seoane-Collazo, P.; Jacas, J.; Morgan, D.A.; Serra, D.; Gallego, R.; Gonzalez, F.; Casals, N.; et al. Central ceramide-induced hypothalamic lipotoxicity and ER stress regulate energy balance. Cell Rep. 2014, 9, 366–377. [Google Scholar] [CrossRef]
- Borg, M.L.; Omran, S.F.; Weir, J.; Meikle, P.J.; Watt, M.J. Consumption of a high-fat diet, but not regular endurance exercise training, regulates hypothalamic lipid accumulation in mice. J. Physiol. 2012, 590, 4377–4389. [Google Scholar] [CrossRef]
- Car, H.; Zendzian-Piotrowska, M.; Prokopiuk, S.; Fiedorowicz, A.; Sadowska, A.; Kurek, K.; Sawicka, D. Ceramide profiles in the brain of rats with diabetes induced by streptozotocin. FEBS J. 2012, 279, 1943–1952. [Google Scholar] [CrossRef]
- Basit, A.; Piomelli, D.; Armirotti, A. Rapid evaluation of 25 key sphingolipids and phosphosphingolipids in human plasma by LC-MS/MS. Anal. Bioanal. Chem. 2015, 407, 5189–5198. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FF | FD | RF 1h | RF 6h | |
|---|---|---|---|---|
| (pmol/mg tissue) | (pmol/mg tissue) | (pmol/mg tissue) | (pmol/mg tissue) | |
| Ceramide (d18:1/20:0) | 4.20 ± 0.37 | 4.37 ± 0.46 | 4.01 ± 0.27 | 3.98 ± 0.36 |
| Ceramide (d18:1/22:0) | 2.65 ± 0.16 | 2.74 ± 0.17 | 2.51 ± 0.12 | 2.51 ± 0.14 |
| Ceramide (d18:1/24:0) | 1.24 ± 0.04 | 1.23 ± 0.01 | 1.13 ± 0.04 | 1.25 ± 0.06 |
| Dihydroceramide (d18:0/16:0) | 0.13 ± 0.012 | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.01 |
| Dihydroceramide (d18:1/24:0) | 0.19 ± 0.01 | 0.18 ± 0.01 | 0.17 ± 0.01 | 0.19 ± 0.03 |
| Dihydroceramide (d18:0/24:1) | 0.31 ± 0.02 | 0.27 ± 0.01 | 0.27 ± 0.03 | 0.26 ± 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vozella, V.; Realini, N.; Misto, A.; Piomelli, D. Feeding Stimulates Sphingosine-1-Phosphate Mobilization in Mouse Hypothalamus. Int. J. Mol. Sci. 2019, 20, 4008. https://doi.org/10.3390/ijms20164008
Vozella V, Realini N, Misto A, Piomelli D. Feeding Stimulates Sphingosine-1-Phosphate Mobilization in Mouse Hypothalamus. International Journal of Molecular Sciences. 2019; 20(16):4008. https://doi.org/10.3390/ijms20164008
Chicago/Turabian StyleVozella, Valentina, Natalia Realini, Alessandra Misto, and Daniele Piomelli. 2019. "Feeding Stimulates Sphingosine-1-Phosphate Mobilization in Mouse Hypothalamus" International Journal of Molecular Sciences 20, no. 16: 4008. https://doi.org/10.3390/ijms20164008
APA StyleVozella, V., Realini, N., Misto, A., & Piomelli, D. (2019). Feeding Stimulates Sphingosine-1-Phosphate Mobilization in Mouse Hypothalamus. International Journal of Molecular Sciences, 20(16), 4008. https://doi.org/10.3390/ijms20164008