Transcription Factor OpWRKY3 Is Involved in the Development and Biosynthesis of Camptothecin and Its Precursors in Ophiorrhiza pumila Hairy Roots

 and
and 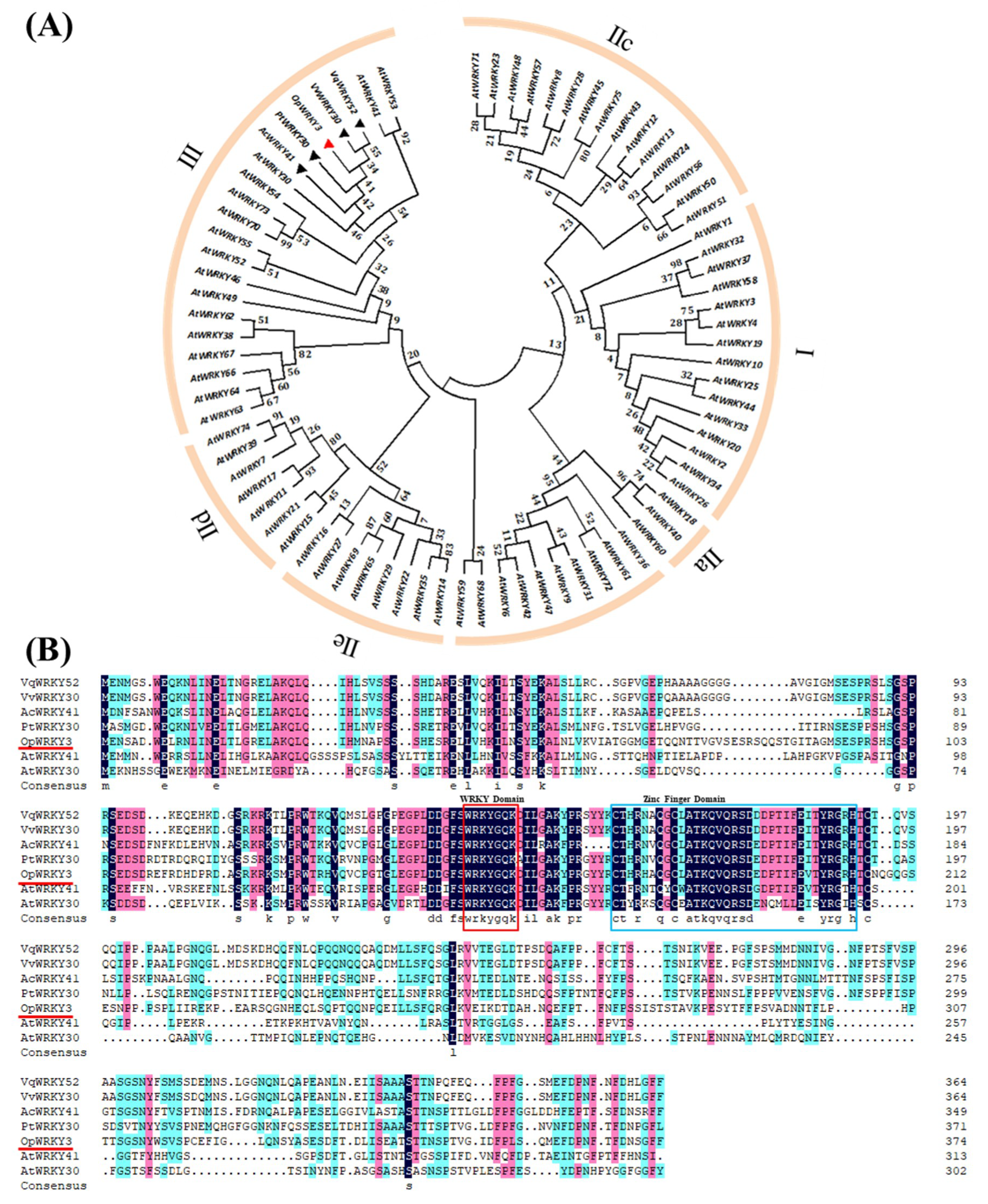{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Bioinformatics and Molecular Characteristics of OpWRKY3
2.2. Tissue and Induction Expression Profiles of OpWRKY3
2.3. Subcellular Localization of OpWRKY3
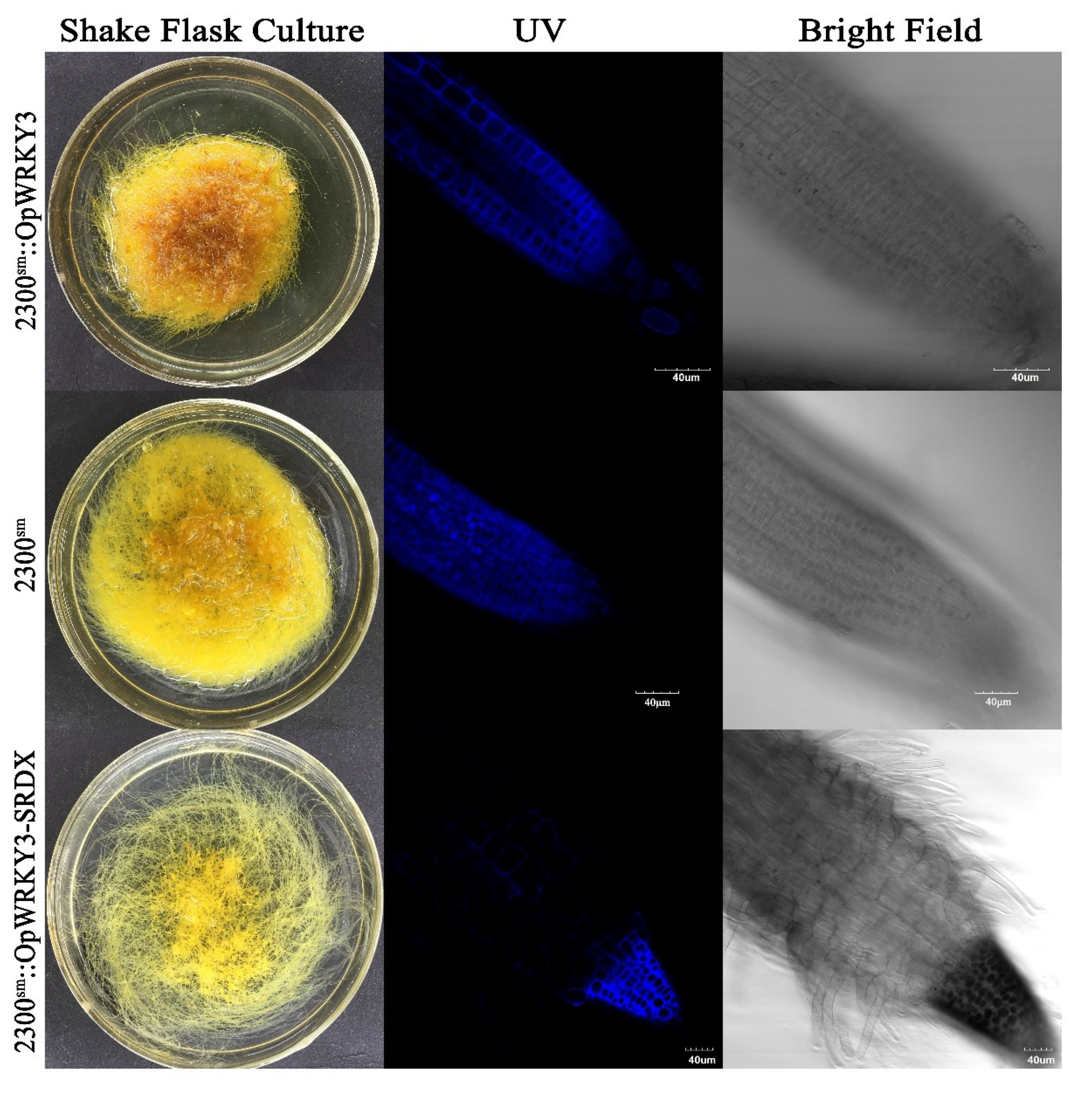
2.4. Acquisition of OpWRKY3 Transgenic Hairy Roots
2.5. Autofluorescent Analysis of Camptothecin in Transgenic Hairy Roots
2.6. Metabolism of Camptothecin and Intermediate Compound in Hairy Roots Overexpressing or Repressing OpWRKY3
2.7. Expression of Camptothecin Biosynthetic Genes in OpWRKY3 Transgenic Hairy Roots
2.8. Activation of the Expression of OpCPR by OpWRKY3
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Elicitor Treatments
4.2. The Gene Clone and Bioinformatics Analyses of OpWRKY3
4.3. Subcellular Localization of OpWRKY3
4.4. Construction of Plant Expression Vectors
4.5. Acquisition of Transgenic Hairy Root Lines
4.6. Quantitative Real-Time PCR
4.7. Confocal Microscope Analysis
4.8. Camptothecin Content Measurement
4.9. Extraction and Analysis of Tryptamine, Loganin and Secologanin
4.10. Yeast-One Hybrid Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| O. pumila | Ophiorrhiza pumila |
| CPT | Camptothecin |
| TIAs | Terpene indole alkaloids |
| GA3 | Gibberellin |
| MJ | Methyl jasmonate |
| ASA | Acetylsalicylic acid |
| SA | Salicylic acid |
| ABA | Abscisic acid |
| TSB | Tryptophan synthase beta |
| TDC | Tryptophan decarboxylase |
| G10H | Geraniol-10-hydroxylase |
| 10-HGO | 10-hydroxy-geraniol oxidoreductase |
| SLS | Secologanin synthase |
| CPR | NADPH:cytochrome P450 reductase |
| STR | Strictosidine synthase |
| SGD | Strictosidine beta-glucosidase |
| TF | Transcription factors |
References
- Lorence, A.; Nessler, C.L. Camptothecin, over four decades of surprising findings. Phytochemistry 2004, 65, 2735–2749. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.; Kumaravadivel, N.; Gnanam, R.; Vellaikumar, S. Rp-HPLC analysis for camptothecin content in Nothapodytes nimmoniana, an endangered medicinal plant. J. Med. Plants Res. 2010, 4, 255–259. [Google Scholar] [CrossRef]
- Saito, K.; Sudo, H.; Yamazaki, M.; Koseki-Nakamura, M.; Kitajima, M.; Takayama, H.; Aimi, N. Feasible production of camptothecin by hairy root culture of Ophiorrhiza pumila. Plant Cell Rep. 2001, 20, 267–271. [Google Scholar] [CrossRef]
- Shaanker, R.U.; Ramesha, B.T.; Ravikanth, G.; Gunaga, R.P.; Vasudeva, R.; Ganeshaiah, K.N. Chemical Profiling of, Nothapodytes nimmoniana for Camptothecin, an Important Anticancer Alkaloid: Towards the Development of a Sustainable Production System, Bioactive Molecules and Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2008; Chapter 10; pp. 198–213. [Google Scholar] [CrossRef]
- Kai, G.Y.; Wu, C.; Gen, L.Y.; Zhang, L.Q.; Cui, L.J.; Ni, X.L. Biosynthesis and biotechnological production of anti-cancer drug camptothecin. Phytochem. Rev. 2015, 14, 525–539. [Google Scholar] [CrossRef]
- Martino, E.; Della Volpe, S.; Terribile, E.; Benetti, E.; Sakaj, M.; Centamore, A.; Sala, A.; Collina, S. The long story of camptothecin: From traditional medicine to drugs. Bioorg. Med. Chem. Lett. 2017, 27, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Beretta, G.L.; Petrangolini, G.; De-Cesare, M.; Pratesi, G.; Perego, P.; Tinelli, S.; Tortoreto, M.; Zucchetti, M.; Frapolli, R.; Bello, E.; et al. Biological properties of IDN5174, a new synthetic camptothecin with the open lactone ring. Cancer Res. 2006, 66, 10976–10982. [Google Scholar] [CrossRef] [PubMed]
- Kai, G.Y.; Dai, L.M.; Mei, X.Y.; Zheng, J.G.; Wang, W.; Lu, Y. In vitro plant regeneration from leaf explants of Ophiorrhiza japonica. Biol. Plant. 2008, 52, 557–560. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, H.; Wang, W.; Qian, Z.; Li, L.; Wang, J.; Zhou, G.; Kai, G. Molecular characterization and expression analysis of a new cDNA encoding strictosidine synthase from Ophiorrhiza japonica. Mol. Biol. Rep. 2009, 36, 1845–1852. [Google Scholar] [CrossRef] [PubMed]
- Sadre, R.; Magallanes-Lundback, M.; Pradhan, S.; Salim, V.; Mesberg, A.; Jones, A.D.; DellaPenna, D. Metabolite diversity in alkaloid biosynthesis: A multilane (Diastereomer) highway for camptothecin synthesis in Camptotheca acuminata. Plant Cell 2016, 28, 1926–1944. [Google Scholar] [CrossRef] [PubMed]
- Sirikantaramas, S.; Yamazaki, M.; Saito, K. Camptothecin: Biosynthesis, Biotechnological Production and Resistance Mechanism(s). Adv. Bot. Res. 2013, 68, 139–161. [Google Scholar]
- Huang, F.C.; Sung, P.H.; Do, Y.Y.; Huang, P.L. Differential expression and functional characterization of the NADPH cytochrome P450 reductase genes from Nothapodytes foetida. Plant Sci. 2012, 190, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Liu, Z.; Wang, Y.; Tang, Z.; Yu, F. A bZIP transcription factor, CaLMF, mediated light-regulated camptothecin biosynthesis in Camptotheca acuminata. Tree Physiol. 2018, 39, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Asano, T.; Yamazaki, Y.; Sirikantaramas, S.; Sudo, H.; Saito, K. Biosynthetic system of camptothecin: An anticancer plant product. Pure Appl. Chem. 2010, 82, 213–218. [Google Scholar] [CrossRef]
- Kai, G.Y.; Li, S.S.; Wang, W.; Lu, Y.; Wang, J.; Liao, P.; Cui, L.J. Molecular cloning and expression analysis of a new gene encoding 3-hydroxy-3-methylglutaryl-CoA synthase from Camptotheca acuminata. Russ. J. Plant Physiol. 2013, 60, 131–138. [Google Scholar] [CrossRef]
- Kai, G.Y.; Teng, X.J.; Cui, L.J.; Li, S.S.; Hao, X.L.; Shi, M.; Yan, B. Effect of three plant hormone elicitors on the camptothecin accumulation and gene transcript profiling in Camptotheca acuminata seedlings. Int. J. Sci. 2014, 3, 86–95. [Google Scholar]
- Asano, T.; Kobayashi, K.; Kashihara, E.; Sudo, H.; Sasaki, R.; Iijima, Y.; Aoki, K.; Shibata, D.; Saito, K.; Yamazaki, M. Suppression of camptothecin biosynthetic genes results in metabolic modification of secondary products in hairy roots of Ophiorrhiza pumila. Phytochemistry 2013, 91, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Ni, X.; Ji, Q.; Teng, X.; Yang, Y.; Wu, C.; Zekria, D.; Zhang, D.; Kai, G. Co-overexpression of geraniol-10-hydroxylase and strictosidine synthase improves anti-cancer drug camptothecin accumulation in Ophiorrhiza pumila. Sci. Rep. 2015, 5, 8227. [Google Scholar] [CrossRef]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Lu, M.; Wang, L.F.; Du, X.H.; Yu, Y.K.; Pan, J.B.; Nan, Z.J.; Han, J.; Wang, W.X.; Zhang, Q.Z.; Sun, Q.P. Molecular cloning and expression analysis of jasmonic acid dependent but salicylic acid independent LeWRKY1. Genet. Mol. Res. 2015, 14, 15390–15398. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Rinerson, C.I.; Rabara, R.C.; Tripathi, P.; Shen, Q.J.; Rushton, P.J. The evolution of WRKY transcription factors. BMC Plant Biol. 2015, 15, 66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [Google Scholar] [CrossRef]
- Bakshi, M.; Oelmuller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 2014, 9, 27700. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Song, Y.; Xing, F.; Wang, N.; Wen, F.; Zhu, C. GhWRKY25, a group I WRKY gene from cotton, confers differential tolerance to abiotic and biotic stresses in transgenic Nicotiana benthamiana. Protoplasma 2016, 253, 1265–1281. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY transcription factors in plant responses to stresses. J. Integr. Plant Biol. 2017, 59, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Hu, Y.; Vannozzi, A.; Wu, K.; Cai, H.; Qin, Y.; Mullis, A.; Lin, Z.; Zhang, L. The WRKY transcription factor family in model plants and crops. Crit. Rev. Plant Sci. 2017, 36, 311–335. [Google Scholar] [CrossRef]
- Viana, V.E.; Busanello, C.; da Maia, L.C.; Pegoraro, C.; Costa de Oliveira, A. Activation of rice WRKY transcription factors: An army of stress fighting soldiers? Curr. Opin. Plant Biol. 2018, 45, 268–275. [Google Scholar] [CrossRef]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: Key components in abscisic acid signalling. Plant Biotechnol. J. 2012, 10, 2–11. [Google Scholar] [CrossRef]
- Devaiah, B.N.; Karthikeyan, A.S.; Raghothama, K.G. WRKY75 transcription factor Is a modulator of phosphate acquisition and root development in Arabidopsis. Plant Physiol. 2007, 143, 1789–1801. [Google Scholar] [CrossRef]
- Ding, Z.; Yan, J.; Li, G.; Wu, Z.; Zhang, S.; Zheng, S. WRKY41 controls Arabidopsis seed dormancy via direct regulation of ABI3 transcript levels not downstream of ABA. Plant J. 2014, 79, 810–823. [Google Scholar] [CrossRef]
- Wei, W.; Cheng, M.N.; Ba, L.J.; Zeng, R.X.; Luo, D.L.; Qin, Y.H.; Liu, Z.L.; Kuang, J.F.; Lu, W.J.; Chen, J.Y.; et al. Pitaya HpWRKY3 Is Associated with Fruit Sugar Accumulation by Transcriptionally Modulating Sucrose Metabolic Genes HpINV2 and HpSuSy1. Int. J. Mol. Sci. 2019, 20, 1890. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Wang, Y.; Shi, M.; Hao, X.; Zhao, W.; Wang, Y.; Ren, J.; Kai, G. Transcription factor SmWRKY1 positively promotes the biosynthesis of tanshinones in Salvia miltiorrhiza. Front. Plant Sci. 2018, 9, 554. [Google Scholar] [CrossRef] [PubMed]
- Chinnapandi, B.; Bucki, P.; Miyara, S.B. SlWRKY45, nematode-responsive tomato WRKY gene, enhances susceptibility to the root knot nematode; M. javanica infection. Plant Signal. Behav. 2017, 12, e1356530. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, T.; Lin, Z.; Gu, B.; Xing, C.; Zhao, L.; Dong, H.; Gao, J.; Xie, Z.; Zhang, S.; et al. A WRKY transcription factor PbrWRKY53 from Pyrus betulaefolia is involved in drought tolerance and AsA accumulation. Plant Biotechnol. J. 2019, 9, 1770–1787. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought responses. Plant Cell. 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yin, Y. WRKY transcription factors are involved in brassinosteroid signaling and mediate the crosstalk between plant growth and drought tolerance. Plant Signal. Behav. 2017, 12, e1365212. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Ma, L.; Wang, S.; Li, L.; Wang, Q.; Yang, R.; Li, Y. Identification and analysis of a candidate WRKY transcription factor gene affecting adventitious root formation using association mapping in Catalpa Scop. DNA Cell Biol. 2019, 38, 297–306. [Google Scholar] [CrossRef]
- Sun, W.J.; Zhan, J.Y.; Zheng, T.R.; Sun, R.; Wang, T.; Tang, Z.Z.; Bu, T.L.; Li, C.L.; Wu, Q.; Chen, H. The jasmonate-responsive transcription factor CbWRKY24 regulates terpenoid biosynthetic genes to promote saponin biosynthesis in Conyzablinii H. Lév. J. Genet. 2018, 97, 1379–1388. [Google Scholar] [CrossRef]
- Suttipanta, N.; Pattanaik, S.; Kulshrestha, M.; Patra, B.; Singh, S.K.; Yuan, L. The transcription factor CrWRKY1 positively regulates the terpenoid indole alkaloid biosynthesis in catharanthus roseus. Plant Physiol. 2011, 157, 2081–2093. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2012, 18, 267–276. [Google Scholar] [CrossRef]
- Deng, C.; Hao, X.; Shi, M.; Fu, R.; Wang, Y.; Zhang, Y.; Zhou, W.; Feng, Y.; Makungac, N.; Kai, G. Tanshinone production could be increased by the expression of SmWRKY2 in Salvia miltiorrhiza hairy roots. Plant Sci. 2019, 284, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.M.; Liu, S.B.; Cai, Q.H.; Tian, X.R. Microscopic fluorescence histological localization and variation of camptothecin in Nothapodytes pittosporoides. Chin. Tradit. Herb. Drugs 2010, 9, 1530–1534. [Google Scholar]
- Sirikantaramas, S.; Sudo, H.; Asano, T.; Yamazaki, M.; Saito, K. Transport of camptothecin in hairy roots of Ophiorrhiza pumila. Phytochemistry 2007, 68, 2881–2886. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Z. Accumulation and localization of camptothecin in young shoot of Camptotheca acuminata. Acta Photophysiol. Sin. 2004, 30, 405–412. [Google Scholar] [CrossRef]
- Pasqua, G.; Monacelli, B.; Valletta, A. Cellular localisation of the anti-cancer drug camptothecin in Camptotheca acuminata Decne (Nyssaceae). Eur. J. Histochem. 2004, 48, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zhou, W.; Zhang, J.; Huang, S.; Wang, H.; Kai, G. Methyl jasmonate induction of tanshinone biosynthesis in Salvia miltiorrhiza hairy roots is mediated by JASMONATE ZIM-DOMAIN repressor proteins. Sci. Rep. 2016, 6, 20919. [Google Scholar] [CrossRef]
- Liu, K.; Li, F.; Zhang, X.; Liu, C.; Zhang, C.; Wu, Z. Progresses and application of researches on EAR-type transcriptional repressor and Chimeric Repressor Gene-Silencing Technology. Mol. Plant Breed. 2010, 8, 1–9. [Google Scholar] [CrossRef]
- Hao, X.; Shi, M.; Cui, L.; Xu, C.; Zhang, Y.; Kai, G. Effects of methyl jasmonate and salicylic acid on the tanshinone production and biosynthetic genes expression in transgenic Salvia miltiorrhiza hairy roots. Biotechnol. Appl. Biochem. 2015, 62, 24–31. [Google Scholar] [CrossRef]
- Huang, Q.; Sun, M.; Yuan, T.; Wang, Y.; Shi, M.; Lu, S.; Tang, B.; Pan, J.; Wang, Y.; Kai, G. The AP2/ERF transcription factor SmERF1L1 regulates the biosynthesis of tanshinones and phenolic acids in Salvia miltiorrhiza. Food Chem. 2019, 274, 368–375. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Wu, C.; Wang, Y.; Xie, C.; Shi, M.; Nile, S.; Zhou, Z.; Kai, G. Transcription Factor OpWRKY3 Is Involved in the Development and Biosynthesis of Camptothecin and Its Precursors in Ophiorrhiza pumila Hairy Roots. Int. J. Mol. Sci. 2019, 20, 3996. https://doi.org/10.3390/ijms20163996
Wang C, Wu C, Wang Y, Xie C, Shi M, Nile S, Zhou Z, Kai G. Transcription Factor OpWRKY3 Is Involved in the Development and Biosynthesis of Camptothecin and Its Precursors in Ophiorrhiza pumila Hairy Roots. International Journal of Molecular Sciences. 2019; 20(16):3996. https://doi.org/10.3390/ijms20163996
Chicago/Turabian StyleWang, Can, Chao Wu, Yao Wang, Chenhong Xie, Min Shi, Shivraj Nile, Zhigang Zhou, and Guoyin Kai. 2019. "Transcription Factor OpWRKY3 Is Involved in the Development and Biosynthesis of Camptothecin and Its Precursors in Ophiorrhiza pumila Hairy Roots" International Journal of Molecular Sciences 20, no. 16: 3996. https://doi.org/10.3390/ijms20163996
APA StyleWang, C., Wu, C., Wang, Y., Xie, C., Shi, M., Nile, S., Zhou, Z., & Kai, G. (2019). Transcription Factor OpWRKY3 Is Involved in the Development and Biosynthesis of Camptothecin and Its Precursors in Ophiorrhiza pumila Hairy Roots. International Journal of Molecular Sciences, 20(16), 3996. https://doi.org/10.3390/ijms20163996