Agrobacterium-Mediated Gene Transient Overexpression and Tobacco Rattle Virus (TRV)-Based Gene Silencing in Cassava

Abstract
1. Introduction
2. Results
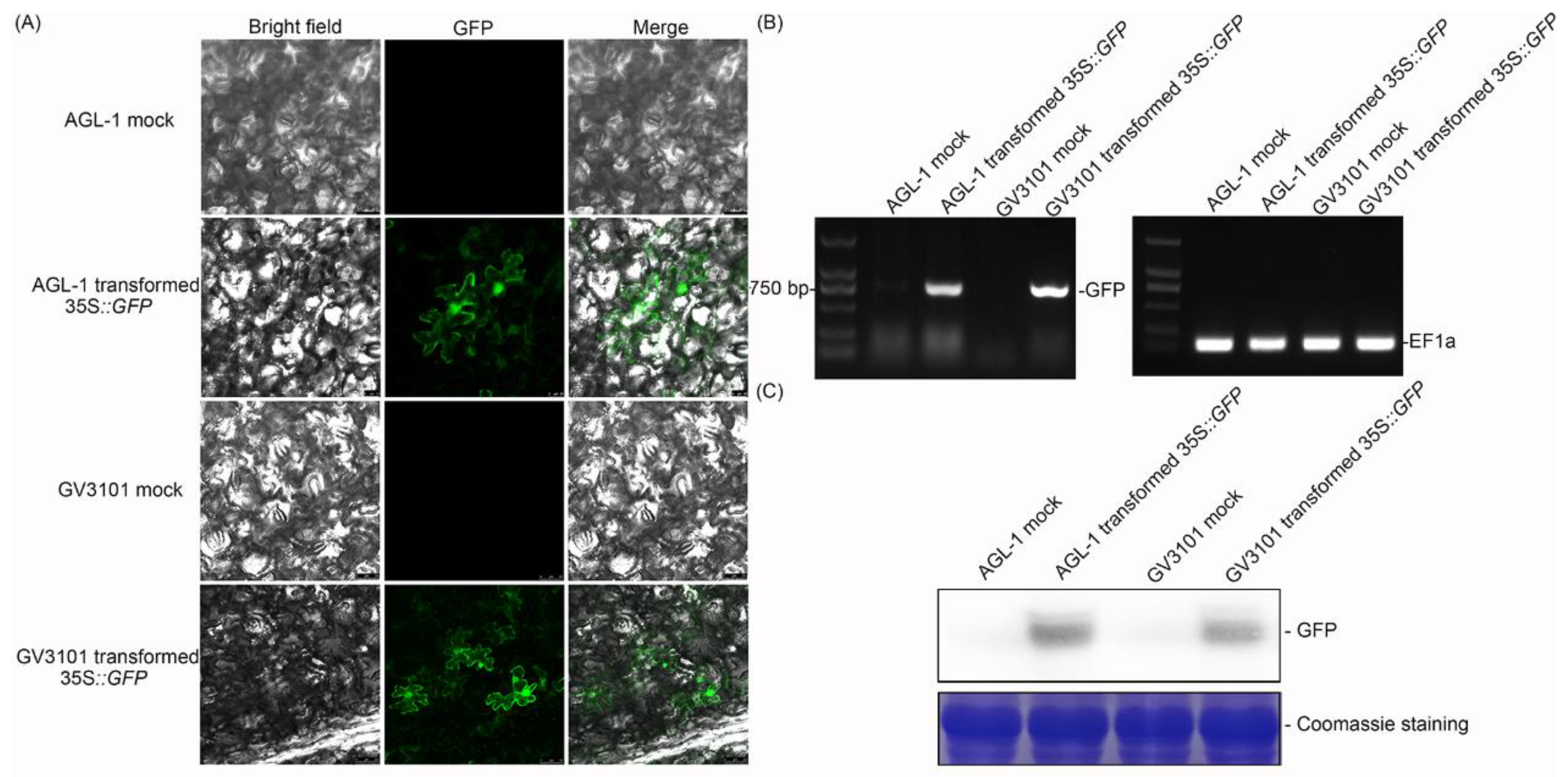
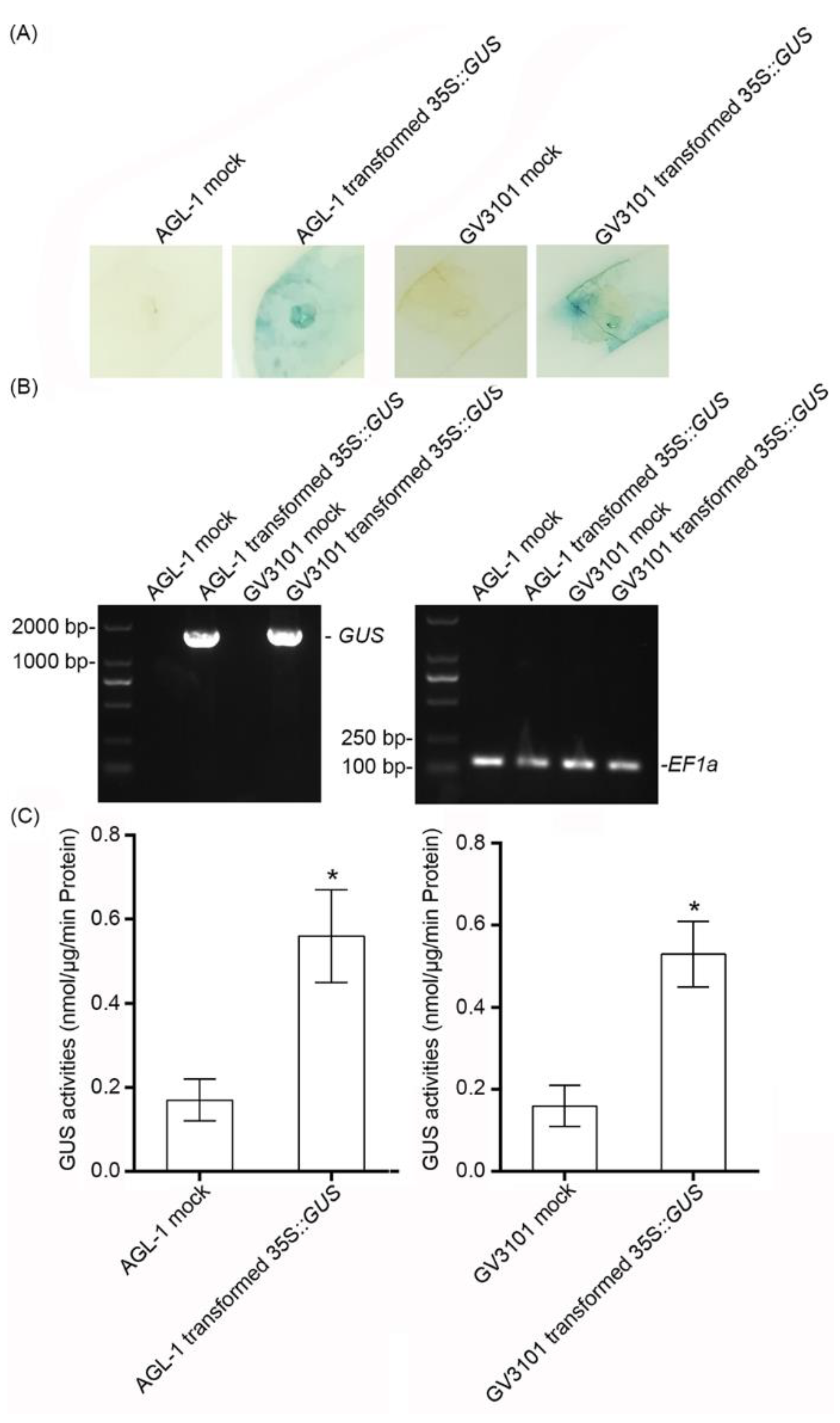
2.1. The Effect of the Transient Expression Assay in Cassava Leaves
2.2. Silencing of MePDS in Cassava
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Vectors and Vector Construction
4.3. Agrobacterium Infiltration of Cassava
4.4. DNA and RNA Extraction
4.5. Reverse-Transcription PCR and Quantitative Real-Time PCR
4.6. Confocal Microscopy Scanning
4.7. Protein Extraction and Western Blot
4.8. GUS Staining and Activity Detection
4.9. PCR Detection of Cassava Plants
4.10. Chlorophyll Content of Cassava Leaves
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, M.C.; Mutka, A.M.; Hummel, A.W.; Berry, J.; Chauhan, R.D.; Vijayaraghavan, A.; Taylor, N.J.; Voytas, D.F.; Chitwood, D.H.; Bart, R.S. Gene expression atlas for the food security crop cassava. New Phytol. 2017, 213, 1632–1641. [Google Scholar]
- Nyaboga, E.; Njiru, J.; Nguu, E.; Gruissem, W.; Vanderschuren, H. Unlocking the potential of tropical root crop biotechnology in east Africa by establishing a genetic transformation platform for local farmer-preferred cassava cultivars. Front. Plant SCI. 2013, 4, 526. [Google Scholar] [CrossRef]
- Wydro, M.; Kozubek, E.; Lehmann, P. Optimization of transient Agrobacterium-mediated gene expression system in leaves of Nicotiana benthamiana. Acta. Biochimica. Pol. 2006, 53, 289–298. [Google Scholar]
- Kim, M.J.; Baek, K.; Park, C.M. Optimization of conditions for transient Agrobacterium-mediated gene expression assays in Arabidopsis. Plant Cell Rep. 2009, 28, 1159–1167. [Google Scholar]
- Ma, L.; Lukasik, E.; Gawehns, F.; Takken, F.L. The Use of Agroinfiltration for Transient Expression of Plant Resistance and Fungal Effector Proteins in Nicotiana benthamiana Leaves. Methods Mol. Biol. 2012, 835, 61. [Google Scholar]
- Ding, B.Q.; Yuan, Y.W. Testing the utility of fluorescent proteins in Mimulus lewisii by an Agrobacterium-mediated transient assay. Plant Cell Rep. 2016, 35, 771–777. [Google Scholar]
- Mani, T.; Manjula, S. Optimization of Agrobacterium-mediated transient gene expression and endogenous gene silencing in Piper colubrinum by vacuum infiltration. Plant Cell Tiss. Org. 2011, 105, 113–119. [Google Scholar]
- Rakouský, S.; Kocábek, T.; Vincenciová, R.; Ondřej, M. Transient β-glucuronidase activity after infiltration of Arabidopsis thaliana by Agrobacterium tumefaciens. Biol. Plant. 1997, 40, 33–41. [Google Scholar] [CrossRef]
- Lu, J.; Bai, M.; Ren, H. An efficient transient expression system for gene function analysis in rose. Plant Methods. 2017, 13, 116. [Google Scholar] [CrossRef]
- Yang, X.F.; Yu, X.Q.; Zhou, Z.; Ma, W.J.; Tang, G.X. High-efficiency Agrobacterium tumefaciens mediated transformation system using cotyledonary node as explants in soybean (Glycine max L.). Acta. Physiol. Plant. 2016, 38, 60. [Google Scholar]
- Fister, A.S.; Shi, Z.; Zhang, Y.F.; Helliwell, E.E.; Maximova, S.N.; Guiltinan, M.J. Protocol: Transient expression system for functional genomics in the tropical tree Theobroma cacao L. Plant Methods. 2016, 12, 19. [Google Scholar] [PubMed]
- Li, H.P.; Li, K.; Guo, Y.T.; Guo, J.D.; Miao, K.T.; Botella, J.R.; Song, C.P.; Miao, Y.C. A transient transformation system for gene characterization in upland cotton (Gossypium hirsutum). Plant Methods 2018, 14, 50. [Google Scholar] [PubMed]
- Kamboj, D.; Yadav, R.C.; Singh, A.; Yadav, N.R.; Singh, D. Plant regeneration and Agrobacterium-mediated transformation in Indian mustard (Brassica juncea). J. Oilseed Brassica. 2016, 6, 191–197. [Google Scholar]
- Bhaskar, P.B.; Venkateshwaran, M.; Wu, L.; Ané, J.M.; Jiang, J. Agrobacterium-Mediated Transient Gene Expression and Silencing: A Rapid Tool for Functional Gene Assay in Potato. PLoS ONE. 2009, 4, e5812. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, T.; Tomczak, A.; Michelmore, R. Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol. J. 2005, 3, 259–273. [Google Scholar] [PubMed]
- Lange, M.; Yellina, A.L.; Orashakova, S.; Becker, A. Virus-induced gene silencing (VIGS) in plants: An overview of target species and the virus-derived vector systems. Methods Mol. Biol. 2013, 975, 1–14. [Google Scholar] [PubMed]
- Hiriart, J.B.; Aro, E.M.; Lehto, K. Dynamics of the VIGS-mediated chimeric silencing of the Nicotiana benthamiana ChlH gene and of the tobacco mosaic virus vector. Mol. Plant Microbe Interact. 2003, 16, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Lacomme, C.; Chapman, S. Use of potato virus X (PVX)-based vectors for gene expression and virus-induced gene silencing (VIGS). Curr. Protoc. Microbiol. 2008, 16, 1. [Google Scholar]
- Zhao, D.; Zhao, J.R.; Huang, X.; Li, N.; Liu, Y.; Huang, Z.J.; Zhang, Z.Y. Functional Analysis of TNBL1 Gene in Wheat Defense Response to Barley yellow dwarf virus Using BSMV-VIGS Technique. Acta. Agronomica. Sinica. 2011, 37, 2106–2110. [Google Scholar] [CrossRef]
- Zhou, T.; Liu, X.; Fan, Z. Use of a Virus Gene Silencing Vector for Maize Functional Genomics Research. Methods Mol. Biol. 2018, 1676, 141–150. [Google Scholar]
- Liu, X.B.; Liu, N.; Li, F.K.; Wu, L.Z.; Zhang, J.; Wang, D.M. Establishment of TRV-mediated Transient Gene-Silencing System in Soybean. Sci. Agri. Sin. 2015, 1212. [Google Scholar]
- Fu, D.Q.; Zhu, B.Z.; Zhu, H.L.; Jiang, W.B.; Luo, Y.B. The application of TRV-mediated VIGS technique in the study of gene function in fruits and vegetables. Plant Physiol. J. 2017, 43, 299–308. [Google Scholar]
- Unver, T.; Budak, H. Virus-Induced Gene Silencing, a Post Transcriptional Gene Silencing Method. Int. J. Plant Genomics. 2009, 198680. [Google Scholar]
- Ratcliff, F.; Martin-Hernandez, A.M.; Baulcombe, D.C. Technical Advance. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [PubMed]
- Zhou, X.F.; Sun, J.D.; Zhao, Z.; Lv, J.; Wei, X.W.; Cai, R.; Xu, H.W. The Feasibility Analysis of PVX and TRV Vectors as the VIGS Tool for Studying the Gene Function. Physics. Procedia. 2002, 23, 46–54. [Google Scholar] [CrossRef][Green Version]
- Wang, C.C.; Cai, X.Z.; Wang, X.M.; Zheng, Z. Optimisation of tobacco rattle virus-induced gene silencing in Arabidopsis. Funct. Plant Biol. 2006, 33, 347–355. [Google Scholar]
- Tian, J.; Pei, H.X.; Zhang, S.; Chen, J.W.; Chen, W.; Yang, R.Y.; Meng, Y.; You, J.; Gao, J.; Ma, N. TRV-GFP: A modified Tobacco rattle virus vector for efficient and visualizable analysis of gene function. J. Exp. Bot. 2014, 65, 311–322. [Google Scholar] [PubMed]
- Tian, J.; Cheng, L.; Han, Z.Y.; Yao, Y.C. Tobacco rattle virus mediated gene silencing in strawberry plants. Plant Cell Tiss. Org. 2015, 120, 1131–1138. [Google Scholar]
- Wang, X.Y.; LÜ, K.; Cai, C.P.; Xu, J.; Guo, W.Z. Establishment and Application of TRV-Mediated Virus-Induced Gene Silencing in Cotton. Acta. Agronomica. Sin. 2014, 40, 1356. [Google Scholar] [CrossRef]
- Anu, K.; Jessymol, K.K.; Chidambareswaren, M.; Gayathri, G.S.; Manjula, S. Down-regulation of osmotin (PR5) gene by virus-induced gene silencing (VIGS) leads to susceptibility of resistant Piper colubrinum to the oomycete pathogen Phytophthora capsici Leonian. Indian, J. Exp. Biol. 2015, 53, 329–334. [Google Scholar]
- Zhang, J.; Yu, D.; Zhang, Y.; Liu, K.; Xu, K.; Zhang, F.; Wang, J.; Tan, G.X.; Nie, X.H.; Ji, Q.H.; et al. Vacuum and Co-cultivation Agroinfiltration of (Germinated) Seeds Results in Tobacco Rattle Virus (TRV) Mediated Whole-Plant Virus-Induced Gene Silencing (VIGS) in Wheat and Maize. Front. Plant Sci. 2017, 8, 393. [Google Scholar] [PubMed]
- Hartl, M.; Merker, H.; Schmidt, D.D.; Baldwin, I.T. Optimized virus-induced gene silencing in Solanum nigrum reveals the defensive function of leucine aminopeptidase against herbivores and the shortcomings of empty vector controls. New Phytol. 2010, 179, 356–365. [Google Scholar] [CrossRef]
- Cutler, S.R.; Ehrhardt, D.W.; Griffitts, J.S.; Somerville, C.R. Random GFP::cDNA fusions enable visualization of subcellular structures in cells of arabidopsis at a high frequency. Proc. Natl. Acad. Sci. USA 2000, 97, 3718–3723. [Google Scholar] [CrossRef] [PubMed]
- Apio, H.B. Production of friable embryogenic callus and regeneration of Ugandan farmer-preferred cassava genotypes. Afr. J. Biotechnol. 2015, 14, 1854–1864. [Google Scholar]
- Tsuda, K.; Qi, Y.; Nguyen, L.V.; Bethke, G.; Tsuda, Y.; Glazebrook, J.; Katagiri, F. An efficient Agrobacterium-mediated transient transformation of Arabidopsis. Plant J. 2012, 69, 713–719. [Google Scholar] [PubMed]
- Díaz, T.P.; Bernal, G.A.; Camilo, L.C. Transient gus gene expression in cassava (manihot esculenta crantz) using agrobacterium tumefaciens leaf infiltration. Revista. Mvz. Cordoba 2014, 19, 4338–4349. [Google Scholar]
- Olivier, V.; Susana, R.; Pere, M.; David, B. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003, 33, 949–956. [Google Scholar]
- Johansen, L.K.; Carrington, J.C. Silencing on the Spot. Induction and Suppression of RNA Silencing in the Agrobacterium-Mediated Transient Expression System. Plant Physiol. 2001, 126, 930–938. [Google Scholar] [CrossRef]
- Andrieu, A.; Breitler, J.C.; Siré, C.; Meynard, D.; Gantet, P.; Guiderdoni, E. Anin planta, agrobacterium-mediated transient gene expression method for inducing gene silencing in rice (oryza sativa L.) leaves. Rice 2012, 5, 23. [Google Scholar] [CrossRef]
- Wege, S.; Scholz, A.; Gleissberg, S.; Becker, A. Highly Efficient Virus-induced Gene Silencing (VIGS) in California Poppy (Eschscholzia californica): An Evaluation of VIGS as a Strategy to Obtain Functional Data from Non-model Plants. Ann. Bot. 2007, 100, 641–649. [Google Scholar]
- Xu, H.; Xu, L.F.; Yang, P.; Cao, Y.; Tang, Y.; He, G.; Yuan, S.X.; Ming, J. Tobacco rattle virus-induced PHYTOENE DESATURASE (PDS) and Mg-chelatase H subunit (ChlH) gene silencing in Solanum pseudocapsicum L. PeerJ. 2018, 6, e4424. [Google Scholar] [PubMed]
- Dai, W.; Wang, M.; Gong, X.; Liu, J.H. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. New Phytol. 2018, 219, 972–989. [Google Scholar] [PubMed]
- Gould, B.; Kramer, E.M. Virus-induced gene silencing as a tool for functional analyses in the emerging model plant Aquilegia (columbine, Ranunculaceae). Plant Methods 2007, 3, 6. [Google Scholar] [PubMed]
- Bruun-Rasmussen, M.; Madsen, C.T.; Jessing, S.; Albrechtsen, M. Stability of Barley stripe mosaic virus-induced gene silencing in barley. Molecular plant-microbe interactions. MPMI 2007, 20, 1323. [Google Scholar] [CrossRef] [PubMed]
- Rotenberg, D.; Thompson, T.S.; German, T.L.; Willis, D.K. Methods for effective real-time RT-PCR analysis of virus-induced gene silencing. J. Virol. Methods 2006, 138, 49–59. [Google Scholar]
- Uçarlı, C.; Tufan, F.; Gürel, F. Expression and genomic integration of transgenes after Agrobacterium-mediated transformation of mature barley embryos. Genet. Mol. Res. 2015, 14, 1096–1105. [Google Scholar]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA transformation-competent arabidopsis genomic library in Agrobacterium. Bio/Technology 1991, 9, 963–967. [Google Scholar] [CrossRef]
- Chabaud, M.; Carvalho-Niebel, F.D.; Barker, D.G. Efficient transformation of Medicago truncatula cv. jemalong using the hypervirulent Agrobacterium tumefaciens strain AGL1. Plant Cell Rep. 2003, 22, 46–51. [Google Scholar] [CrossRef]
- Fofana, I.B.; Sangaré, A.; Collier, R.; Taylor, C.; Fauquet, C.M. A geminivirus-induced gene silencing system for gene function validation in cassava. Plant Mol Biol. 2004, 56, 613–624. [Google Scholar] [CrossRef]
- Beyene, G.; Chauhan, R.D.; Taylor, N.J. A rapid virus-induced gene silencing (VIGS) method for assessing resistance and susceptibility to cassava mosaic disease. Virol. J. 2017, 14, 47. [Google Scholar]
- Matias, L.E.; Joel-Elias, K. Cassava germinivirus agroclones for virus-induced gene silencing in cassava leaves and roots. Plant Methods 2018. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Qin, G.; Chen, T.; Tian, S. The mode of action of remorin1 in regulating fruit ripening at transcriptional and post-transcriptional levels. New Phytol. 2018, 219, 1406–1420. [Google Scholar] [CrossRef] [PubMed]
- Inskeep, W.P.; Bloom, P.R. Extinction coefficients of chlorophyll a and B in N, N-dimethylformamide and 80% acetone. Plant Physiol. 1985, 77, 483–485. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Report Gene | Bacterial Strain | Number | The Positive Rate of Reporter Gene |
|---|---|---|---|
| GFP | GV3101 | 20 | 60.00% |
| GUS | GV3101 | 20 | 75.00% |
| GFP | AGL-1 | 20 | 65.00% |
| GUS | AGL-1 | 20 | 75.00% |
| Bacterial Strain | Number | The Positive Rate of Relative Transcript Level (<60%) | The Positive Rate of Albino Phenotype |
|---|---|---|---|
| AGL-1 | 8 | 75.00% | 37.50% |
| GV3101 | 8 | 62.50% | 12.50% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, H.; Xie, Y.; Liu, G.; Wei, Y.; Hu, W.; Shi, H. Agrobacterium-Mediated Gene Transient Overexpression and Tobacco Rattle Virus (TRV)-Based Gene Silencing in Cassava. Int. J. Mol. Sci. 2019, 20, 3976. https://doi.org/10.3390/ijms20163976
Zeng H, Xie Y, Liu G, Wei Y, Hu W, Shi H. Agrobacterium-Mediated Gene Transient Overexpression and Tobacco Rattle Virus (TRV)-Based Gene Silencing in Cassava. International Journal of Molecular Sciences. 2019; 20(16):3976. https://doi.org/10.3390/ijms20163976
Chicago/Turabian StyleZeng, Hongqiu, Yanwei Xie, Guoyin Liu, Yunxie Wei, Wei Hu, and Haitao Shi. 2019. "Agrobacterium-Mediated Gene Transient Overexpression and Tobacco Rattle Virus (TRV)-Based Gene Silencing in Cassava" International Journal of Molecular Sciences 20, no. 16: 3976. https://doi.org/10.3390/ijms20163976
APA StyleZeng, H., Xie, Y., Liu, G., Wei, Y., Hu, W., & Shi, H. (2019). Agrobacterium-Mediated Gene Transient Overexpression and Tobacco Rattle Virus (TRV)-Based Gene Silencing in Cassava. International Journal of Molecular Sciences, 20(16), 3976. https://doi.org/10.3390/ijms20163976