Transcriptional Response of Subcutaneous White Adipose Tissue to Acute Cold Exposure in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Acute Cold Exposure Stimulates sWAT Browning in Mice
2.2. Differentially Expressed Gene Screening and Gene Ontology Analysis in Cold-Treated sWAT
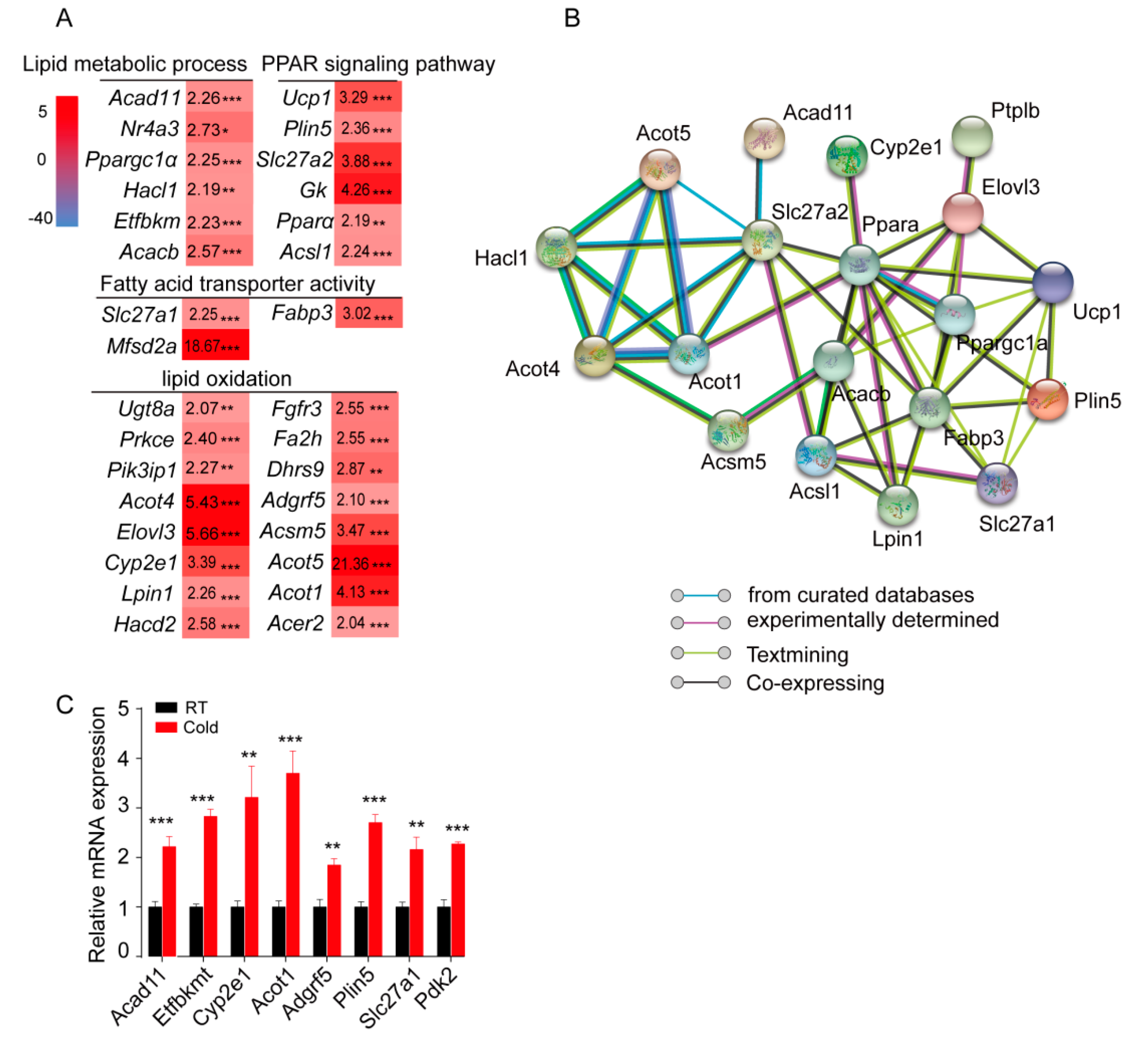
2.3. Cold Exposure Significantly Alters the Expression of Genes Involved in the Fatty Acid Metabolic Process and Lipid Oxidation in sWAT
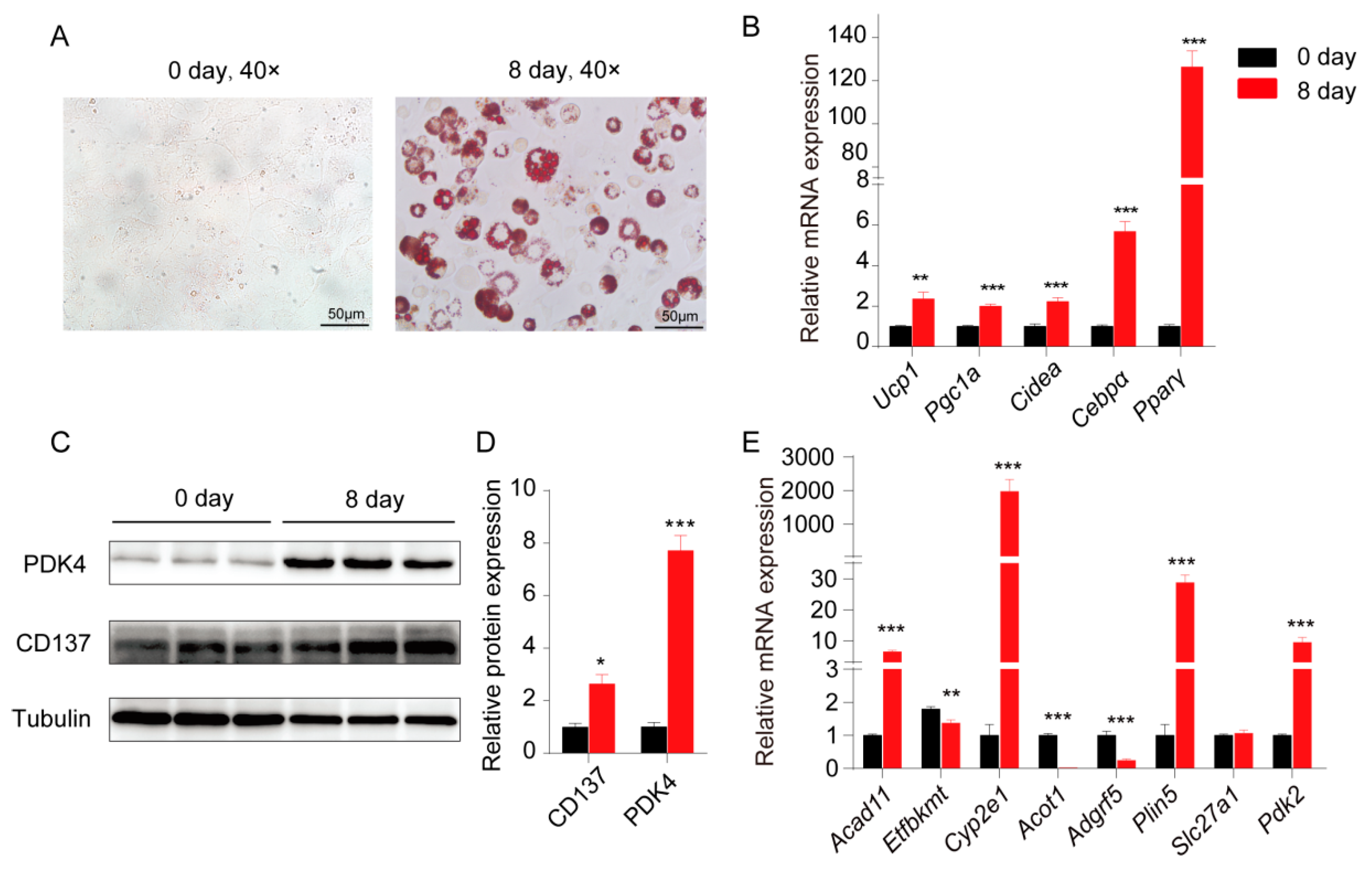
2.4. Validation of the Potential WAT Browning Candidate Genes in 3T3-L1 Differentiation Toward Beige Adipocyte
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Collection
4.2. RNA Preparation and Quantitative Real-Time PCR
4.3. RNA-Seq Analysis
4.4. Gene Ontology (GO) and Signaling Network Analyses
4.5. Hematoxylin and Eosin (H&E) Staining
4.6. Western Blot Analysis
4.7. Cell Culture and in Vitro Browning
4.8. Oil Red O Staining
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| sWAT | subcutaneous white adipose tissues |
| BAT | brown adipose tissue |
| WAT | white adipose tissue |
| DEGs | differentially expressed genes |
| GO | gene ontology |
| RT | room temperature |
| QPCR | quantitative real-time PCR |
| PPARγ | peroxisome proliferator-activated receptor-γ |
References
- Maximilian, T.; Gerdtham, U.G.; Peter, N.; Sanjib, S. Economic Burden of Obesity: A Systematic Literature Review. Int. J. Environ. Res. Public Health 2017, 14, 435. [Google Scholar]
- Hubert, H. Obesity as an independent risk factor for cardiovascular disease: A 26-year follow-up of participants in the Framingham Heart Study. Circulation 1983, 67, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Miroslava, C.; Michaela, K.; Jana, D.; Pavel, P.; Martina, G.; Vaclav, B.; Dana, M.; Jitka, K. Mitochondria in White, Brown, and Beige Adipocytes. Stem Cells Int. 2016, 2016, 6067349. [Google Scholar]
- Wang, W.; Seale, P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 2016, 17, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, B.; Foster, M.T. Beiging of white adipose tissue as a therapeutic strategy for weight loss in humans. Horm. Mol. Biol. Clin. Investig. 2017, 31, undefined. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Levy, J.D.; Zhang, Y.; Frontini, A.; Kolodin, D.P.; Svensson, K.J.; Lo, J.C.; Zeng, X.; Ye, L.; Khandekar, M.J. Ablation of PRDM16 and beige adipose causes metabolic dysfunction and a subcutaneous to visceral fat switch. Cell 2014, 156, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.I.; Middelbeek, R.J.W.; Townsend, K.L.; Lee, M.-Y.; Takahashi, H.; So, K.; Hitchcox, K.M.; Markan, K.R.; Hellbach, K.; Hirshman, M.F. A Novel Role for Subcutaneous Adipose Tissue in Exercise-Induced Improvements in Glucose Homeostasis. Diabetes 2015, 64, 2002–2014. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef]
- Schulz, T.J.; Huang, P.; Huang, T.L.; Xue, R.; Mcdougall, L.E.; Townsend, K.L.; Cypess, A.M.; Mishina, Y.; Gussoni, E.; Tseng, Y.H. Brown-fat paucity due to impaired BMP signalling induces compensatory browning of white fat. Nature 2013, 495, 379–383. [Google Scholar] [CrossRef]
- Garcia, R.A.; Roemmich, J.N.; Claycombe, K.J. Evaluation of markers of beige adipocytes in white adipose tissue of the mouse. Nutr. Metab. 2016, 13, 24. [Google Scholar] [CrossRef]
- Piao, Z.; Zhai, B.; Jiang, X.; Dong, M.; Jin, W. Reduced adiposity by compensatory WAT browning upon iBAT removal in mice. Biochem. Biophys. Res. Commun. 2018, 501, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Tao, C.; Huang, S.; Wang, Y.; Wei, G.; Zhang, Y.; Qi, D.; Wang, Y.; Li, K. Changes in white and brown adipose tissue microRNA expression in cold-induced mice. Biochem. Biophys. Res. Commun. 2015, 463, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Qimuge, N.R.; Qin, J.; Cai, R.; Li, X.; Chu, G.Y.; Pang, W.J.; Yang, G.S. Acute and chronic cold exposure differentially affects the browning of porcine white adipose tissue. Anim. Int. J. Anim. Biosci. 2018, 12, 1435–1441. [Google Scholar] [CrossRef]
- Lin, J.; Cao, C.; Tao, C.; Ye, R.; Zhao, J. Cold adaptation in pigs depends on UCP3 in beige adipocytes. J. Mol. Cell Biol. 2017, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Rosell, M.; Kaforou, M.; Frontini, A.; Okolo, A.; Chan, Y.W.; Nikolopoulou, E.; Millership, S.; Fenech, M.E.; Macintyre, D.; Turner, J.O. Brown and white adipose tissues: Intrinsic differences in gene expression and response to cold exposure in mice. Am. J. Physiol. Endocrinol. Metab. 2014, 306, 945–964. [Google Scholar] [CrossRef]
- Steensels, S.; Ersoy, B.A. Fatty acid activation in thermogenic adipose tissue. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2018, 1864, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.W.; Ozdemir, C.; Kawano, Y.; LeClair, K.B.; Vernochet, C.; Kahn, C.R.; Hagen, S.J.; Cohen, D.E. Thioesterase Superfamily Member 2/Acyl-CoA Thioesterase 13 (Them2/Acot13) Regulates Adaptive Thermogenesis in Mice. J. Biol. Chem. 2013, 288, 33376–33386. [Google Scholar] [CrossRef]
- Ellis, J.M.; Bowman, C.E.; Wolfgang, M.J.; Zane, A. Metabolic and Tissue-Specific Regulation of Acyl-CoA Metabolism. PLoS ONE 2015, 10, e0116587. [Google Scholar] [CrossRef] [PubMed]
- Franklin, M.P.; Sathyanarayan, A.; Mashek, D.G. Acyl-CoA Thioesterase 1 (ACOT1) Regulates PPARα to Couple Fatty Acid Flux with Oxidative Capacity During Fasting. Diabetes 2017, 66, 2112. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bell, M.; Sreenevasan, U.; Hu, H.; Liu, J.; Dalen, K.; Londos, C.; Yamaguchi, T.; Rizzo, M.A.; Coleman, R. Unique Regulation of Adipose Triglyceride Lipase (ATGL) by Perilipin 5, a Lipid Droplet-associated Protein. J. Biol. Chem. 2011, 286, 15707–15715. [Google Scholar] [CrossRef]
- Wang, H.; Sreenivasan, U.; Hu, H.; Saladino, A.; Polster, B.M.; Lund, L.M.; Gong, D.-w.; Stanley, W.C.; Sztalryd, C. Perilipin 5, a lipid droplet-associated protein, provides physical and metabolic linkage to mitochondria. J. Lipid Res. 2011, 52, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Bosma, M.; Minnaard, R.; Sparks, L.M.; Schaart, G.; Losen, M.; Baets, M.H.d.; Duimel, H.; Kersten, S.; Bickel, P.E.; Schrauwen, P. The lipid droplet coat protein perilipin 5 also localizes to muscle mitochondria. Histochem. Cell Biol. 2012, 137, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Laurens, C.; Bourlier, V.; Mairal, A.; Louche, K.; Badin, P.-M.; Mouisel, E.; Montagner, A.; Marette, A.; Tremblay, A.; Weisnagel, J.S. Perilipin 5 fine-tunes lipid oxidation to metabolic demand and protects against lipotoxicity in skeletal muscle. Sci. Rep. 2016, 6, 38310. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Lin, J.; Huang, J.; Zhang, H.; Zhang, R.; Zhang, X.; Cao, C.; Hambly, C.; Qin, G.; Yao, J. Reconstitution of UCP1 using CRISPR/Cas9 in the white adipose tissue of pigs decreases fat deposition and improves thermogenic capacity. Proc. Natl. Acad. Sci. USA 2017, 114, E9474. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Zhang, Z.; Zheng, H.; Xiong, S.; Su, Y.; Ma, X.; Yi, C. Browning of Human Subcutaneous Adipose Tissue after Its Transplantation in Nude Mice. Plast. Reconstr. Surg. 2018, 142, 1. [Google Scholar] [CrossRef] [PubMed]
- Minchul, S.; Tae-Won, G.; Mi, C.; Minhee, B.; Jae-Sam, H.; Mi-Ae, K.; Eun-Young, Y. Tenebrio molitor Larvae Inhibit Adipogenesis through AMPK and MAPKs Signaling in 3T3-L1 Adipocytes and Obesity in High-Fat Diet-Induced Obese Mice. Int. J. Mol. Sci. 2017, 18, 518. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Pan, J.; Cao, C.; Zhang, L.; Zhao, Y.; Fan, Y.; Li, K.; Tao, C.; Wang, Y. Transcriptional Response of Subcutaneous White Adipose Tissue to Acute Cold Exposure in Mice. Int. J. Mol. Sci. 2019, 20, 3968. https://doi.org/10.3390/ijms20163968
Liang X, Pan J, Cao C, Zhang L, Zhao Y, Fan Y, Li K, Tao C, Wang Y. Transcriptional Response of Subcutaneous White Adipose Tissue to Acute Cold Exposure in Mice. International Journal of Molecular Sciences. 2019; 20(16):3968. https://doi.org/10.3390/ijms20163968
Chicago/Turabian StyleLiang, Xiaojuan, Jianfei Pan, Chunwei Cao, Lilan Zhang, Ying Zhao, Yiping Fan, Kui Li, Cong Tao, and Yanfang Wang. 2019. "Transcriptional Response of Subcutaneous White Adipose Tissue to Acute Cold Exposure in Mice" International Journal of Molecular Sciences 20, no. 16: 3968. https://doi.org/10.3390/ijms20163968
APA StyleLiang, X., Pan, J., Cao, C., Zhang, L., Zhao, Y., Fan, Y., Li, K., Tao, C., & Wang, Y. (2019). Transcriptional Response of Subcutaneous White Adipose Tissue to Acute Cold Exposure in Mice. International Journal of Molecular Sciences, 20(16), 3968. https://doi.org/10.3390/ijms20163968