Dental Pulp Stem Cell-Derived Factors Alleviate Subarachnoid Hemorrhage-Induced Neuroinflammation and Ischemic Neurological Deficits


 and
and 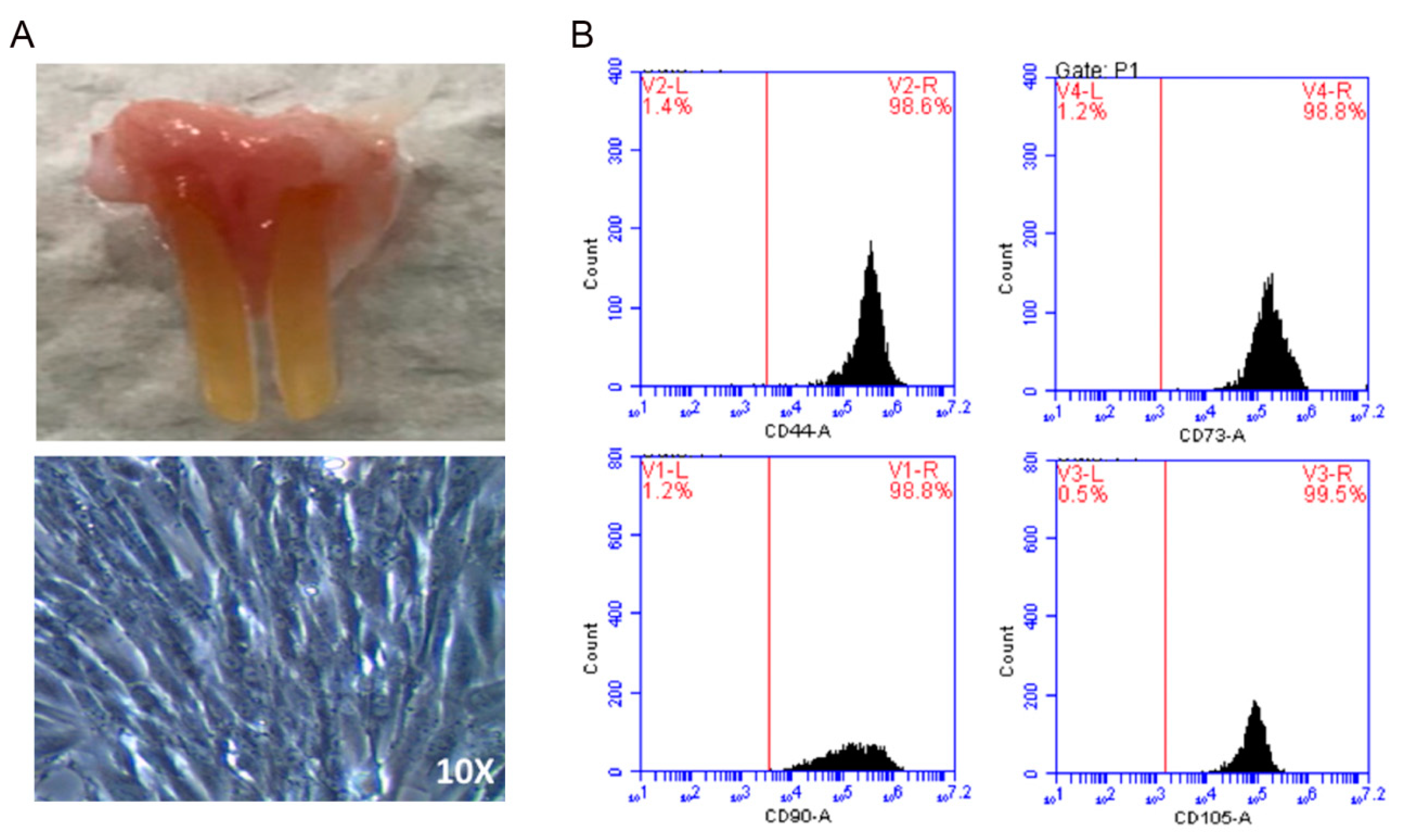{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Preparation of Dental Pulp-Derived Mesenchymal Stem Cells
2.2. The Conditioned Medium of Cultivated DPSCs Ameliorated aSAH in Rats
2.3. IGF-1 as the Major Active Constituent in the DPSC-CM
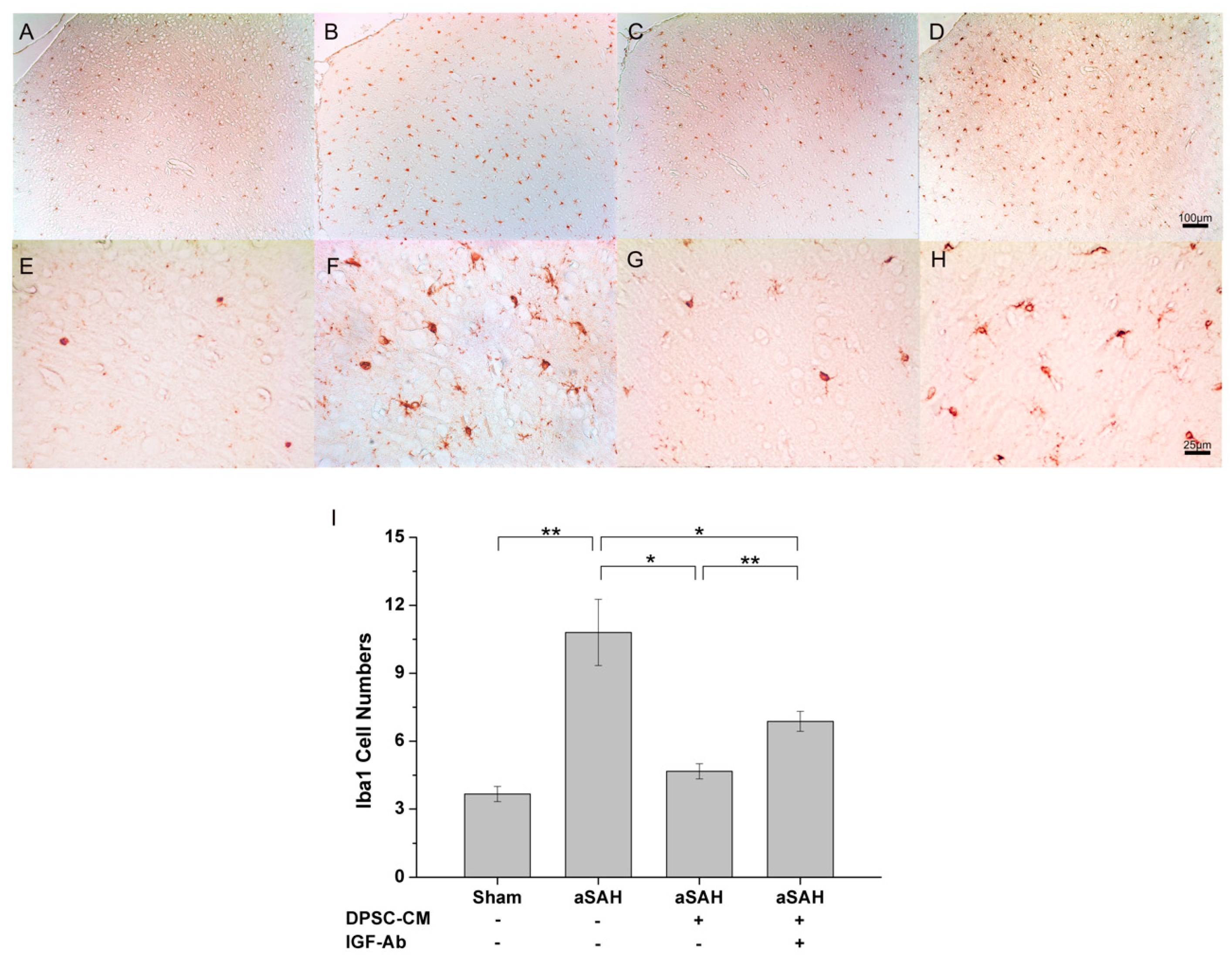
2.4. DPSC-CM Alleviates Neuroinflammation in aSAH Partially via IGF-1
2.5. DPSC-CM Improves Microcirculation in aSAH Partially via IGF-1
2.6. Therapeutic Efficacy of DPSC-CM on Cognitive and Motor Impairments in aSAH-Injured Rats
3. Discussion
4. Materials and Methods
4.1. Experimental Model of aSAH
4.2. Cranial Window for Observation of Microcirculation
4.3. DPSC Isolation and Culture
4.4. Preparation of Conditioned Medium
4.5. Treatment Algorithm
4.6. Brain Preparation for Immunohistochemistry Analysis
4.7. Rotarod Test
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| aSAH | Aneurysmal subarachnoid hemorrhage |
| DPSC | Dental pulp-derived stem cell |
| IGF-1 | Insulin-like growth factor-1 |
| DPSC-CM | DPSC-derived conditioned medium |
| TIMP2 | tissue inhibitor of metalloproteinases 2 |
| TMP1 | tissue inhibitor of metalloproteinases 1 |
| TGF-β | transforming growth factor β |
References
- Suarez, J.I.; Tarr, R.W.; Selman, W.R. Aneurysmal subarachnoid hemorrhage. N. Engl. J. Med. 2006, 354, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Schievink, W.I. Intracranial aneurysms. N. Engl. J. Med. 1997, 336, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Broderick, J.P.; Brott, T.G.; Duldner, J.E.; Tomsick, T.; Leach, A. Initial and recurrent bleeding are the major causes of death following subarachnoid hemorrhage. Stroke 1994, 25, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- Kassell, N.F.; Torner, J.C.; Haley, E.C., Jr.; Jane, J.A.; Adams, H.P.; Kongable, G.L. The international cooperative study on the timing of aneurysm surgery. Part 1: Overall management results. J. Neurosurg. 1990, 73, 18–36. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Manoel, A.L.; Macdonald, R.L. Neuroinflammation as a target for intervention in subarachnoid hemorrhage. Front. Neurol. 2018, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Tae-Hoon, L.; Yoon-Seok, L. Transplantation of mouse embryonic stem cell after middle cerebral artery occlusion. Acta Cir. Bras. 2012, 27, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagisawa, D.; Qi, M.; Kim, D.H.; Kitamura, Y.; Inden, M.; Tsuchiya, D.; Takata, K.; Taniguchi, T.; Yoshimoto, K.; Shimohama, S.; et al. Improvement of focal ischemia-induced rat dopaminergic dysfunction by striatal transplantation of mouse embryonic stem cells. Neurosci. Lett. 2006, 407, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yasuhara, T.; Shingo, T.; Muraoka, K.; Kameda, M.; Takeuchi, A.; Yano, A.; Kurozumi, K.; Agari, T.; Miyoshi, Y.; et al. Embryonic neural stem cells transplanted in middle cerebral artery occlusion model of rats demonstrated potent therapeutic effects, compared to adult neural stem cells. Brain Res. 2008, 1234, 172–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.J.; Chang, C.M.; Tsai, S.K.; Chang, Y.L.; Chou, S.J.; Huang, S.S.; Tai, L.K.; Chen, Y.C.; Ku, H.H.; Li, H.Y.; et al. Functional improvement of focal cerebral ischemia injury by subdural transplantation of induced pluripotent stem cells with fibrin glue. Stem Cells Dev. 2010, 19, 1757–1767. [Google Scholar] [CrossRef]
- Luo, L.; He, Y.; Wang, X.; Key, B.; Lee, B.H.; Li, H.; Ye, Q. Potential roles of dental pulp stem cells in neural regeneration and repair. Stem Cells Int. 2018, 2018, 1731289. [Google Scholar]
- Venkatesh, K.; Sen, D. Mesenchymal stem cells as a source of dopaminergic neurons: A potential cell based therapy for parkinson's disease. Curr. Stem Cell Res. 2017, 12, 326–347. [Google Scholar] [CrossRef] [PubMed]
- Honmou, O. Stem cell therapy for dementia. Brain Nerve 2015, 67, 93–98. [Google Scholar] [PubMed]
- Hsuan, Y.C.; Lin, C.H.; Chang, C.P.; Lin, M.T. Mesenchymal stem cell-based treatments for stroke, neural trauma, and heat stroke. Brain Behav. 2016, 6, e00526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.G.; Long, G.; Tyler, G.; Stefan, A.; Broadfoot, S.J.; Piccinini, A.M.; Middleton, J.; Kehoe, O. Mesenchymal stem cell-conditioned medium reduces disease severity and immune responses in inflammatory arthritis. Sci. Rep. 2017, 7, 18019. [Google Scholar] [CrossRef] [PubMed]
- Aboutaleb, N.; Faezi, M.; Nasseri Maleki, S.; Nazarinia, D.; Razavi Tousi, S.M.T.; Hashemirad, N. Conditioned medium obtained from mesenchymal stem cells attenuates focal cerebral ischemia reperfusion injury through activation of erk1/erk2-bdnf signaling pathway. J. Chem. Neuroanat. 2019, 97, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Van Velthoven, C.T.; Kavelaars, A.; van Bel, F.; Heijnen, C.J. Repeated mesenchymal stem cell treatment after neonatal hypoxia-ischemia has distinct effects on formation and maturation of new neurons and oligodendrocytes leading to restoration of damage, corticospinal motor tract activity, and sensorimotor function. J. Neurosci. 2010, 30, 9603–9611. [Google Scholar] [CrossRef] [PubMed]
- van Velthoven, C.T.; Kavelaars, A.; van Bel, F.; Heijnen, C.J. Mesenchymal stem cell treatment after neonatal hypoxic-ischemic brain injury improves behavioral outcome and induces neuronal and oligodendrocyte regeneration. Brain Behav. Immun. 2010, 24, 387–393. [Google Scholar] [CrossRef] [PubMed]
- van Velthoven, C.T.; Sheldon, R.A.; Kavelaars, A.; Derugin, N.; Vexler, Z.S.; Willemen, H.L.; Maas, M.; Heijnen, C.J.; Ferriero, D.M. Mesenchymal stem cell transplantation attenuates brain injury after neonatal stroke. Stroke 2013, 44, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Tang, S.C.; Lee, J.E.; Tsai, J.C.; Lai, D.M.; Lin, W.C.; Lin, C.P.; Tu, Y.K.; Hsieh, S.T. Impaired microcirculation after subarachnoid hemorrhage in an in vivo animal model. Sci Rep. 2018, 8, 13315. [Google Scholar] [CrossRef]
- Miller, B.A.; Turan, N.; Chau, M.; Pradilla, G. Inflammation, vasospasm, and brain injury after subarachnoid hemorrhage. Biomed. Res. Int. 2014, 2014, 384342. [Google Scholar] [CrossRef] [PubMed]
- Provencio, J.J. Inflammation in subarachnoid hemorrhage and delayed deterioration associated with vasospasm: A review. Acta Neurochir. Suppl. 2013, 115, 233–238. [Google Scholar] [PubMed]
- Ito, D.; Imai, Y.; Ohsawa, K.; Nakajima, K.; Fukuuchi, Y.; Kohsaka, S. Microglia-specific localisation of a novel calcium binding protein, iba1. Brain Res. Mol. Brain Res. 1998, 57, 1–9. [Google Scholar] [CrossRef]
- Noailles, A.; Fernandez-Sanchez, L.; Lax, P.; Cuenca, N. Microglia activation in a model of retinal degeneration and tudca neuroprotective effects. J. Neuroinflamm. 2014, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, K.; Imai, Y.; Kanazawa, H.; Sasaki, Y.; Kohsaka, S. Involvement of iba1 in membrane ruffling and phagocytosis of macrophages/microglia. J. Cell Sci. 2000, 113, 3073–3084. [Google Scholar] [PubMed]
- Kim, H.J.; Park, J.S. Usage of human mesenchymal stem cells in cell-based therapy: Advantages and disadvantages. Dev. Reprod 2017, 21, 1–10. [Google Scholar] [CrossRef]
- Khan, F.A.; Almohazey, D.; Alomari, M.; Almofty, S.A. Isolation, culture, and functional characterization of human embryonic stem cells: Current trends and challenges. Stem Cells Int. 2018, 2018, 1429351. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Liang, G.; Zhang, Y. Genetic and epigenetic variations in ipscs: Potential causes and implications for application. Cell Stem Cell 2013, 13, 149–159. [Google Scholar] [CrossRef]
- Cittadini, A.; Monti, M.G.; Castiello, M.C.; D'Arco, E.; Galasso, G.; Sorriento, D.; Saldamarco, L.; De Paulis, A.; Napoli, R.; Iaccarino, G.; et al. Insulin-like growth factor-1 protects from vascular stenosis and accelerates re-endothelialization in a rat model of carotid artery injury. J. Thromb. Haemost. 2009, 7, 1920–1928. [Google Scholar] [CrossRef]
- Tang, X.; Chen, F.; Lin, Q.; You, Y.; Ke, J.; Zhao, S. Bone marrow mesenchymal stem cells repair the hippocampal neurons and increase the expression of igf-1 after cardiac arrest in rats. Exp. Med. 2017, 14, 4312–4320. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Estevez, V.; Defterali, C.; Vicario-Abejon, C. Igf-i: A key growth factor that regulates neurogenesis and synaptogenesis from embryonic to adult stages of the brain. Front. Neurosci. 2016, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, S.; Arafa, D.; Tropea, D. Insulin-like growth factor 1: At the crossroads of brain development and aging. Front. Cell Neurosci. 2017, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Torres Aleman, I. Insulin-like growth factor-1 and central neurodegenerative diseases. Endocrinol. Metab. Clin. N. Am. 2012, 41, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Paulsson, J.F.; Blinder, P.; Burstyn-Cohen, T.; Du, D.; Estepa, G.; Adame, A.; Pham, H.M.; Holzenberger, M.; Kelly, J.W.; et al. Reduced igf-1 signaling delays age-associated proteotoxicity in mice. Cell 2009, 139, 1157–1169. [Google Scholar] [CrossRef]
- George, C.; Gontier, G.; Lacube, P.; Francois, J.C.; Holzenberger, M.; Aid, S. The alzheimer's disease transcriptome mimics the neuroprotective signature of igf-1 receptor-deficient neurons. Brain 2017, 140, 2012–2027. [Google Scholar] [CrossRef]
- Al-Tamimi, Y.Z.; Orsi, N.M.; Quinn, A.C.; Homer-Vanniasinkam, S.; Ross, S.A. A review of delayed ischemic neurologic deficit following aneurysmal subarachnoid hemorrhage: Historical overview, current treatment, and pathophysiology. World Neurosurg. 2010, 73, 654–667. [Google Scholar] [CrossRef]
- Nie, G.; Wen, X.; Liang, X.; Zhao, H.; Li, Y.; Lu, J. Additional evidence supports association of common genetic variants in mmp3 and timp2 with increased risk of chronic achilles tendinopathy susceptibility. J. Sci. Med. Sport 2019. [Google Scholar] [CrossRef]
- Hu, B.H.; Cai, Q.; Hu, Z.; Patel, M.; Bard, J.; Jamison, J.; Coling, D. Metalloproteinases and their associated genes contribute to the functional integrity and noise-induced damage in the cochlear sensory epithelium. J. Neurosci. 2012, 32, 14927–14941. [Google Scholar] [CrossRef]
- Maloney-Wilensky, E.; Le Roux, P. The physiology behind direct brain oxygen monitors and practical aspects of their use. Childs Nerv. Syst. 2010, 26, 419–430. [Google Scholar] [CrossRef]
- Gerhard Vogel, H. Psychotropic and neurotropic activity. In Drug Discovery and Evaluation; Springer: Berlin/Heidelberg, Germany, 2007; pp. 565–876. [Google Scholar]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.-F.; Chen, K.-W.; Chien, Y.; Lai, Y.-H.; Hsieh, S.-T.; Ma, H.-Y.; Wang, K.-C.; Shiau, C.-Y. Dental Pulp Stem Cell-Derived Factors Alleviate Subarachnoid Hemorrhage-Induced Neuroinflammation and Ischemic Neurological Deficits. Int. J. Mol. Sci. 2019, 20, 3747. https://doi.org/10.3390/ijms20153747
Chen T-F, Chen K-W, Chien Y, Lai Y-H, Hsieh S-T, Ma H-Y, Wang K-C, Shiau C-Y. Dental Pulp Stem Cell-Derived Factors Alleviate Subarachnoid Hemorrhage-Induced Neuroinflammation and Ischemic Neurological Deficits. International Journal of Molecular Sciences. 2019; 20(15):3747. https://doi.org/10.3390/ijms20153747
Chicago/Turabian StyleChen, Te-Fu, Kuo-We Chen, Yueh Chien, Ying-Hsiu Lai, Sung-Tsang Hsieh, Hsin-Yi Ma, Kou-Chung Wang, and Chia-Yang Shiau. 2019. "Dental Pulp Stem Cell-Derived Factors Alleviate Subarachnoid Hemorrhage-Induced Neuroinflammation and Ischemic Neurological Deficits" International Journal of Molecular Sciences 20, no. 15: 3747. https://doi.org/10.3390/ijms20153747