Exploring the N-Glycosylation Profile of Glycoprotein B from Human Cytomegalovirus Expressed in CHO and Nicotiana tabacum BY-2 Cells

 and
and
Abstract

1. Introduction
2. Results
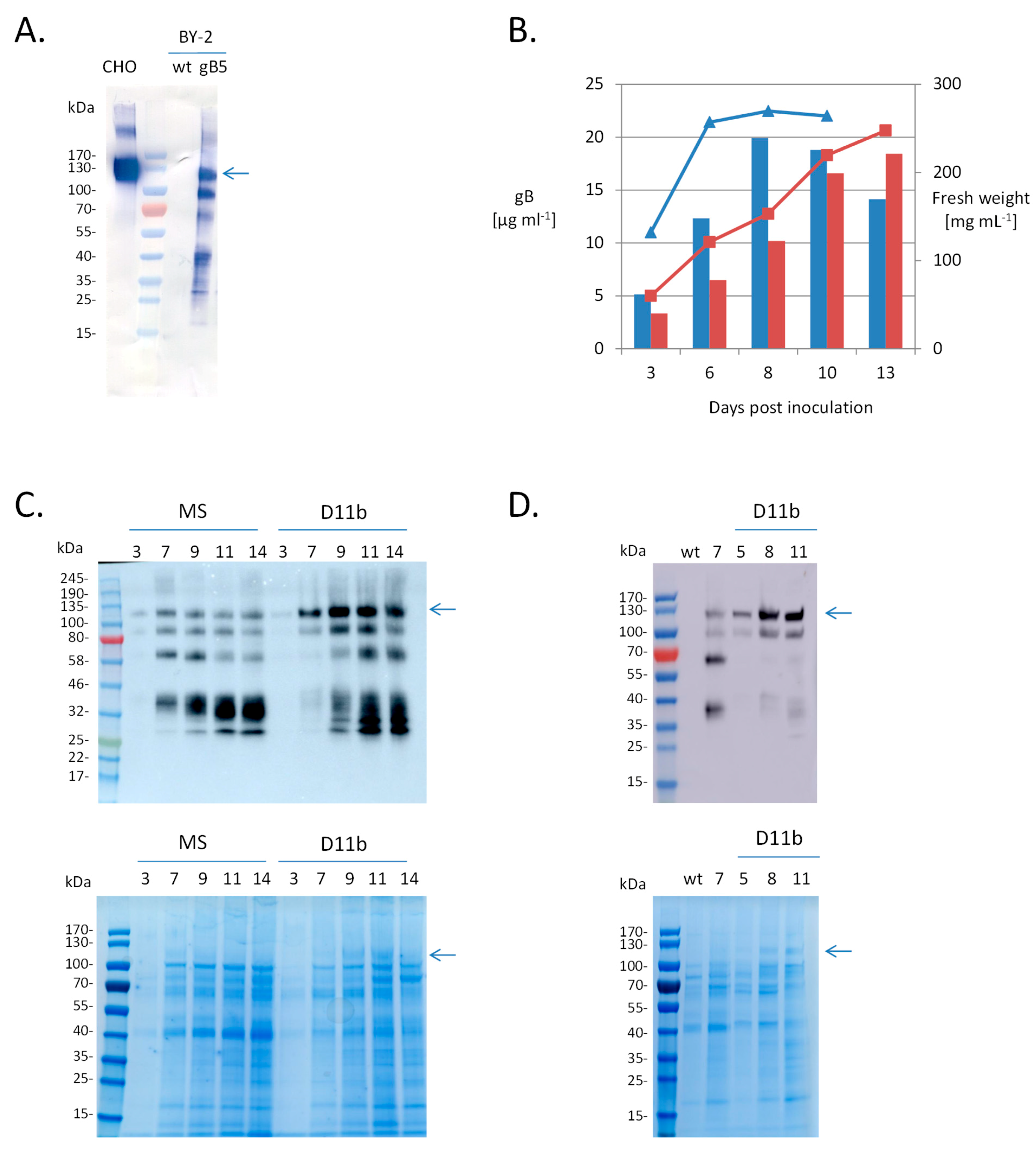
2.1. Expression of HCMV gB Ectodomain in Nicotiana tabacum BY-2 Cells
2.2. Purification of BY-2-Produced gB
2.3. N-Glycan Repertoire of CHO-Produced gB
2.4. Site-Specific N-Glycan Analysis
2.5. Glycosite Occupancy
3. Discussion
4. Materials and Methods
4.1. Expression of gB Ectodomain in BY-2 Cells
4.2. Semi-Purification of gB from BY-2 Culture Medium
4.3. Expression and Purification of gB Ectodomain in CHO Cells
4.4. Glycoproteomic Workflow
4.5. Glycoproteomic Data Treatment
4.6. N-Glycosylation Profile of gB Produced in CHO Cells
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bagdonaite, I.; Wandall, H.H. Global aspects of viral glycosylation. Glycobiology 2018, 28, 443–467. [Google Scholar] [CrossRef] [PubMed]
- Margolin, E.; Chapman, R.; Williamson, A.L.; Rybicki, E.P.; Meyers, A.E. Production of complex viral glycoproteins in plants as vaccine immunogens. Plant Biotechnol. J. 2018, 16, 1531–1545. [Google Scholar] [CrossRef]
- MEDICAGO. Medicago Announces Phase 3 Study of VLP Quadrivalent Influenza Vaccine. Available online: https://media.medicago.com/webfolder_download/951cb9f9114b2ed4e132b1c563ba870f/medicago-announces-phase-3-study-of-vlp-quadrivalent-influenza-vaccine/6936dd00ddfd77a354f293f8144269ffb22c029c/medicago-announces-phase-3-study-of-vlp-quadrivalent-influenza-vaccine.pdf (accessed on 30 July 2019).
- Santos, R.B.; Abranches, R.; Fischer, R.; Sack, M.; Holland, T. Putting the spotlight back on plant suspension cultures. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, N. On the way to commercializing plant cell culture platform for biopharmaceuticals: Present status and prospect. Pharm. Bioprocess. 2014, 2, 499–518. [Google Scholar] [CrossRef] [PubMed]
- Shaaltiel, Y.; Bartfeld, D.; Hashmueli, S.; Baum, G.; Brill-Almon, E.; Galili, G.; Dym, O.; Boldin-Adamsky, S.A.; Silman, I.; Sussman, J.L.; et al. Production of glucocerebrosidase with terminal mannose glycans for enzyme replacement therapy of Gaucher’s disease using a plant cell system. Plant Biotechnol. J. 2007, 5, 579–590. [Google Scholar] [CrossRef]
- Tekoah, Y.; Shulman, A.; Kizhner, T.; Ruderfer, I.; Fux, L.; Nataf, Y.; Bartfeld, D.; Ariel, T.; Gingis-Velitski, S.; Hanania, U.; et al. Large-scale production of pharmaceutical proteins in plant cell culture-the protalix experience. Plant Biotechnol. J. 2015, 13, 1199–1208. [Google Scholar] [CrossRef]
- Smith, M.; Mason, H.; Shuler, M. Hepatitis B surface antigen (HBsAg) expression in plant cell culture: Kinetics of antigen accumulation in batch culture and its intracellular form. Biotechnol. Bioeng. 2002, 80, 812–822. [Google Scholar] [CrossRef]
- Lienard, D.; Dinh, O.T.; Van Oort, E.; Van Overtvelt, L.; Bonneau, C.; Wambre, E.; Bardor, M.; Cosette, P.; Didier-Laurent, A.; De Borne, F.D.; et al. Suspension-cultured BY-2 tobacco cells produce and mature immunologically active house dust mite allergens. Plant Biotechnol. J. 2007, 5, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Arteaga, A.; Shuler, M.L. Influence of culture medium supplementation of tobacco NT1 cell suspension cultures on the N-glycosylation of human secreted alkaline phosphatase. Biotechnol. Bioeng. 2007, 97, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Raven, N.; Rasche, S.; Kuehn, C.; Anderlei, T.; Klöckner, W.; Schuster, F.; Henquet, M.; Bosch, D.; Büchs, J.; Fischer, R.; et al. Scaled-up manufacturing of recombinant antibodies produced by plant cells in a 200-L orbitally-shaken disposable bioreactor. Biotechnol. Bioeng. 2015, 112, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Holland, T.; Sack, M.; Rademacher, T.; Schmale, K.; Altmann, F.; Stadlmann, J.; Fischer, R.; Hellwig, S. Optimal nitrogen supply as a key to increased and sustained production of a monoclonal full-size antibody in BY-2 suspension culture. Biotechnol. Bioeng. 2010, 107, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Navarre, C.; Smargiasso, N.; Duvivier, L.; Nader, J.; Far, J.; de Pauw, E.; Boutry, M. N-Glycosylation of an IgG antibody secreted by Nicotiana tabacum BY-2 cells can be modulated through co-expression of human β-1,4-galactosyltransferase. Transgenic Res. 2017, 26, 375–384. [Google Scholar] [CrossRef]
- Hanania, U.; Ariel, T.; Tekoah, Y.; Fux, L.; Sheva, M.; Gubbay, Y.; Weiss, M.; Oz, D.; Azulay, Y.; Turbovski, A.; et al. Establishment of a tobacco BY2 cell line devoid of plant-specific xylose and fucose as a platform for the production of biotherapeutic proteins. Plant Biotechnol. J. 2017, 15, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Jung, H.S.; Lee, S.J.; Park, C.I.; Lim, S.M.; Park, H.; Kim, B.S.; Na, K.H.; Han, G.J.; Bae, J.W.; et al. Glycan structure and serum half-life of recombinant CTLA4Ig, an immunosuppressive agent, expressed in suspension-cultured rice cells with coexpression of human β1,4-galactosyltransferase and human CTLA4Ig. Glycoconj. J. 2015, 32, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-W.; Kim, N.-S.; Jang, S.-H.; Shin, Y.-J.; Yang, M.-S. Production and characterization of recombinant human acid α-glucosidase in transgenic rice cell suspension culture. J. Biotechnol. 2016, 226, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Le Mauff, F.; Mercier, G.; Chan, P.; Burel, C.; Vaudry, D.; Bardor, M.; Vezina, L.P.; Couture, M.; Lerouge, P.; Landry, N. Biochemical composition of haemagglutinin-based influenza virus-like particle vaccine produced by transient expression in tobacco plants. Plant Biotechnol. J. 2015, 13, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, Y.; Sack, M.; Montefiori, D.; Forthal, D.; Mao, L.; Abanto, S.H.; Urban, L.; Landucci, G.; Fischer, R.; Jiang, X. Rapid High-Level Production of Functional HIV Broadly Neutralizing Monoclonal Antibodies in Transient Plant Expression Systems. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Burke, H.G.; Heldwein, E.E. Crystal Structure of the Human Cytomegalovirus Glycoprotein B. PLoS Pathogens 2015, 11. [Google Scholar] [CrossRef]
- Sharma, S.; Wisner, T.W.; Johnson, D.C.; Heldwein, E.E. HCMV gB shares structural and functional properties with gB proteins from other herpesviruses. Virology 2013, 435, 239–249. [Google Scholar] [CrossRef]
- Schleiss, M.R. Recombinant cytomegalovirus glycoprotein B vaccine: Rethinking the immunological basis of protection. Proc. Natl. Acad. Sci. USA 2018, 115, 6110–6112. [Google Scholar] [CrossRef] [PubMed]
- Schleiss, M.R. Cytomegalovirus vaccines under clinical development. J. Virus Erad. 2016, 2, 198–207. [Google Scholar] [PubMed]
- Chandramouli, S.; Ciferri, C.; Nikitin, P.A.; Caló, S.; Gerrein, R.; Balabanis, K.; Monroe, J.; Hebner, C.; Lilja, A.E.; Settembre, E.C.; et al. Structure of HCMV glycoprotein B in the postfusion conformation bound to a neutralizing human antibody. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Cao, Z.; Wang, S.; Lee, R.B.; Wang, X.; Murata, H.; Adler, S.P.; McVoy, M.A.; Snapper, C.M. Novel trimeric human cytomegalovirus glycoprotein B elicits a high-titer neutralizing antibody response. Vaccine 2018, 36, 5580–5590. [Google Scholar] [CrossRef] [PubMed]
- Britt, W.J.; Vugler, L.G. Processing of the gp55-116 envelope glycoprotein complex (gB) of human cytomegalovirus. J. Virol. 1989, 63, 403–410. [Google Scholar] [PubMed]
- Bagdonaite, I.; Nordén, R.; Joshi, H.J.; King, S.L.; Vakhrushev, S.Y.; Olofsson, S.; Wandall, H.H. Global mapping of o-glycosylation of varicella zoster virus, human cytomegalovirus, and Epstein-Barr virus. J. Biol. Chem. 2016, 291, 12014–12028. [Google Scholar] [CrossRef] [PubMed]
- Tackaberry, E.S.; Dudani, A.K.; Prior, F.; Tocchi, M.; Sardana, R.; Altosaar, I.; Ganz, P.R. Development of biopharmaceuticals in plant expression systems: Cloning, expression and immunological reactivity of human cytomegalovirus glycoprotein B (UL55) in seeds of transgenic tobacco. Vaccine 1999, 17, 3020–3029. [Google Scholar] [CrossRef]
- Wright, K.E.; Prior, F.; Sardana, R.; Altosaar, I.; Dudani, A.K.; Ganz, P.R.; Tackaberry, E.S. Sorting of glycoprotein B from human cytomegalovirus to protein storage vesicles in seeds of transgenic tobacco. Transgenic Res. 2001, 10, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Tackaberry, E.S.; Prior, F.; Bell, M.; Tocchi, M.; Porter, S.; Mehic, J.; Ganz, P.R.; Sardana, R.; Altosaar, I.; Dudani, A. Increased yield of heterologous viral glycoprotein in the seeds of homozygous transgenic tobacco plants cultivated underground. Genome 2003, 46, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Tackaberry, E.S.; Prior, F.A.; Rowlandson, K.; Tocchi, M.; Mehic, J.; Porter, S.; Walsh, M.; Schleiss, M.R.; Ganz, P.R.; Sardana, R.K.; et al. Sustained expression of human cytomegalovirus glycoprotein B (UL55) in the seeds of homozygous rice plants. Mol. Biotechnol. 2008, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, N.; Grömping, U.; Lipperts, A.; Raven, N.; Fischer, R.; Schillberg, S. Optimization of BY-2 cell suspension culture medium for the production of a human antibody using a combination of fractional factorial designs and the response surface method. Plant Biotechnol. J. 2013, 11, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.; Britt, W.J.; Compton, T. Expression, purification, and characterization of a soluble form of human cytomegalovirus glycoprotein B. Virology 1997, 239, 198–205. [Google Scholar] [CrossRef] [PubMed]
- CFG. Symbol and Text Nomenclature for Representation of Glycan Structure. Available online: http://www.functionalglycomics.org/static/consortium/Nomenclature.shtml (accessed on 30 July 2019).
- Morsa, D.; Baiwir, D.; La Rocca, R.; Zimmerman, T.A.; Hanozin, E.; Grifnée, E.; Longuespée, R.; Meuwis, M.-A.; Smargiasso, N.; De Pauw, E.; et al. Multi-Enzymatic Limited Digestion—The Next-Generation Sequencing for Proteomics? J. Proteome Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Castilho, A.; Beihammer, G.; Pfeiffer, C.; Göritzer, K.; Montero-Morales, L.; Vavra, U.; Maresch, D.; Grünwald-Gruber, C.; Altmann, F.; Steinkellner, H.; et al. An oligosaccharyltransferase from Leishmania major increases the N-glycan occupancy on recombinant glycoproteins produced in Nicotiana benthamiana. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Montero-Morales, L.; Maresch, D.; Castilho, A.; Turupcu, A.; Ilieva, K.M.; Crescioli, S.; Karagiannis, S.N.; Lupinek, C.; Oostenbrink, C.; Altmann, F.; et al. Recombinant plant-derived human IgE glycoproteomics. J. Proteom. 2017, 161, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Bosch, D.; Schots, A. Plant glycans: Friend or foe in vaccine development? Expert Rev. Vaccines 2010, 9, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.J.; Landry, N.; Trépanier, S.; Mercier, G.; Dargis, M.; Couture, M.; D’Aoust, M.A.; Vézina, L.P. Human antibody response to N-glycans present on plant-made influenza virus-like particle (VLP) vaccines. Vaccine 2014, 32, 6098–6106. [Google Scholar] [CrossRef]
- Mercx, S.; Smargiasso, N.; Chaumont, F.; de Pauw, E.; Boutry, M.; Navarre, C. Inactivation of the β(1,2)-xylosyltransferase and the α(1,3)-fucosyltransferase genes in Nicotiana tabacum BY-2 cells by a multiplex CRISPR/Cas9 strategy results in glycoproteins without plant-specific glycans. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Mercx, S.; Tollet, J.; Magy, B.; Navarre, C.; Boutry, M. Gene inactivation by CRISPR-Cas9 in Nicotiana tabacum BY-2 suspension cells. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Yang, S.; Hu, Y.; Sokoll, L.; Zhang, H. Simultaneous quantification of N- and O-glycans using a solid-phase method. Nat. Protoc. 2017, 12, 1229–1244. [Google Scholar] [CrossRef]
- Göritzer, K.; Maresch, D.; Altmann, F.; Obinger, C.; Strasser, R. Exploring Site-Specific N-Glycosylation of HEK293 and Plant-Produced Human IgA Isotypes. J. Proteome Res. 2017, 16, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Kang, P.; Mechref, Y.; Novotny, M.V. High-throughput solid-phase permethylation of glycans prior to mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 721–734. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N-Glycosite | Peptides Identified after PNGase | N-Glycan Occupancy | CHO | BY-2 |
|---|---|---|---|---|
| N68-N73 | ANETIYNTTLK | N68-N73 non-glycosylated | 0.7 | 2.9 |
| ADETIYNTTLK | N68 glycosylated/N73 non-glycosylated | 56.5 | 43.4 | |
| ANETIYDTTLK | N68 non-glycosylated/N73 glycosylated | 3.2 | 3.5 | |
| ADETIYDTTLK | N68-N73 glycosylated | 39.7 | 50.2 | |
| N85 | YGDVVGVNTTK | N85 non-glycosylated | 2.3 | ND |
| YGDVVGVDTTK | N85 glycosylated | 97.7 | 100 | |
| N286 | NASYFGENADK | N286 non-glycosylated | 9.7 | ND |
| DASYFGENADK | N286 glycosylated | 90.3 | 100 | |
| N383 | QEVNMSDSALDCVR | N383 non-glycosylated | 4.3 | 3.2 |
| QEVDMSDSALDCVR | N383 glycosylated | 95.7 | 96.8 | |
| N405-N409 | LQQIFNTSYNQTYEK | N405-N409 non-glycosylated | 0.3 | ND |
| LQQIFDTSYNQTYEK | N405 glycosylated/N409 non-glycosylated | ND | ND | |
| LQQIFNTSYDQTYEK | N405 non-glycosylated/N409 glycosylated | 28.5 | 36.4 | |
| LQQIFDTSYDQTYEK | N405-N409 glycosylated | 71.1 | 63.6 | |
| N208-N302-N341-N417-N452-N554-N585 | NXS/T peptides | non-glycosylated | ND | ND |
| DXS/T peptides | glycosylated | 100 | 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smargiasso, N.; Nader, J.; Rioux, S.; Mazzucchelli, G.; Boutry, M.; De Pauw, E.; Chaumont, F.; Navarre, C. Exploring the N-Glycosylation Profile of Glycoprotein B from Human Cytomegalovirus Expressed in CHO and Nicotiana tabacum BY-2 Cells. Int. J. Mol. Sci. 2019, 20, 3741. https://doi.org/10.3390/ijms20153741
Smargiasso N, Nader J, Rioux S, Mazzucchelli G, Boutry M, De Pauw E, Chaumont F, Navarre C. Exploring the N-Glycosylation Profile of Glycoprotein B from Human Cytomegalovirus Expressed in CHO and Nicotiana tabacum BY-2 Cells. International Journal of Molecular Sciences. 2019; 20(15):3741. https://doi.org/10.3390/ijms20153741
Chicago/Turabian StyleSmargiasso, Nicolas, Joseph Nader, Stéphane Rioux, Gabriel Mazzucchelli, Marc Boutry, Edwin De Pauw, François Chaumont, and Catherine Navarre. 2019. "Exploring the N-Glycosylation Profile of Glycoprotein B from Human Cytomegalovirus Expressed in CHO and Nicotiana tabacum BY-2 Cells" International Journal of Molecular Sciences 20, no. 15: 3741. https://doi.org/10.3390/ijms20153741
APA StyleSmargiasso, N., Nader, J., Rioux, S., Mazzucchelli, G., Boutry, M., De Pauw, E., Chaumont, F., & Navarre, C. (2019). Exploring the N-Glycosylation Profile of Glycoprotein B from Human Cytomegalovirus Expressed in CHO and Nicotiana tabacum BY-2 Cells. International Journal of Molecular Sciences, 20(15), 3741. https://doi.org/10.3390/ijms20153741