GARP as an Immune Regulatory Molecule in the Tumor Microenvironment of Glioblastoma Multiforme

Abstract
1. Introduction
2. Results
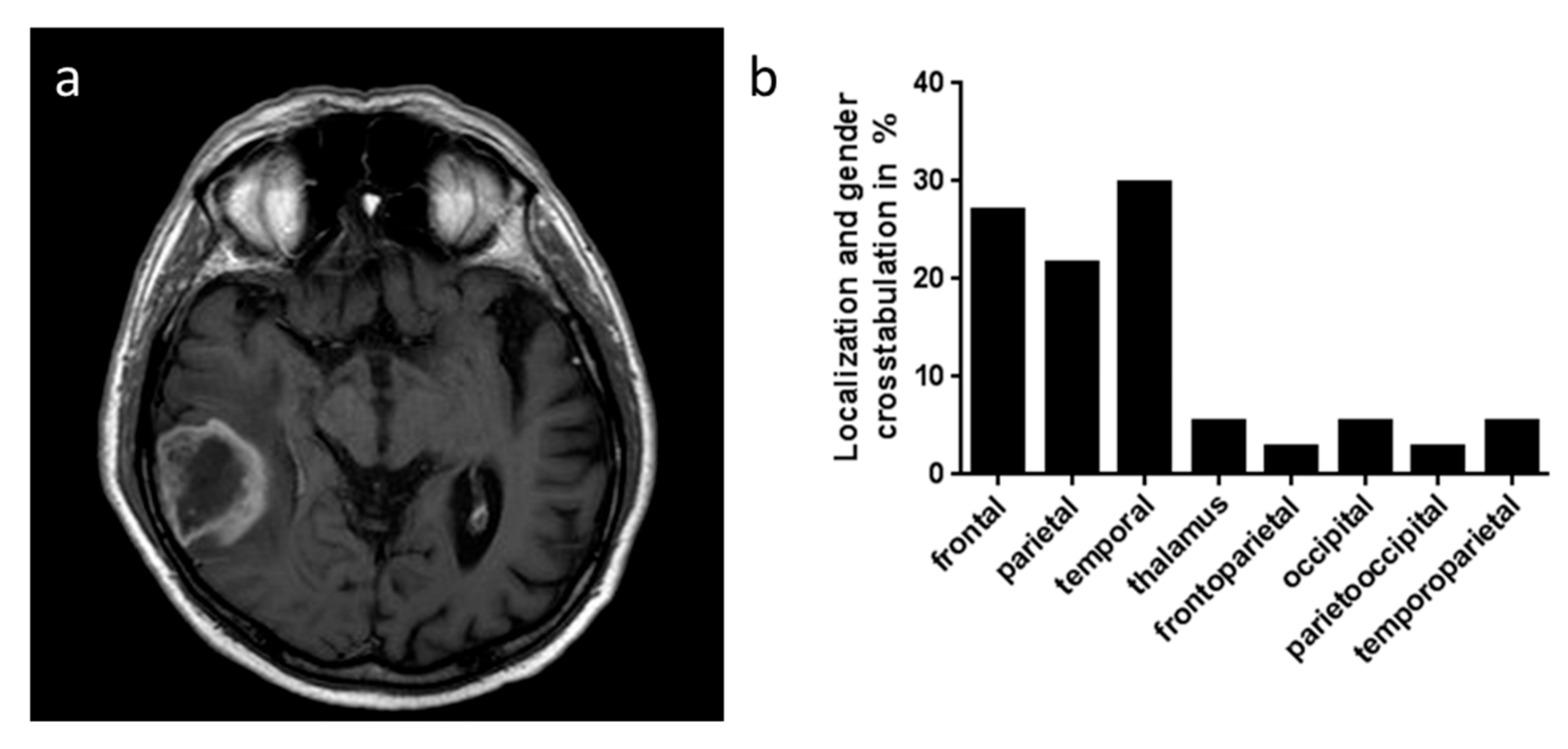
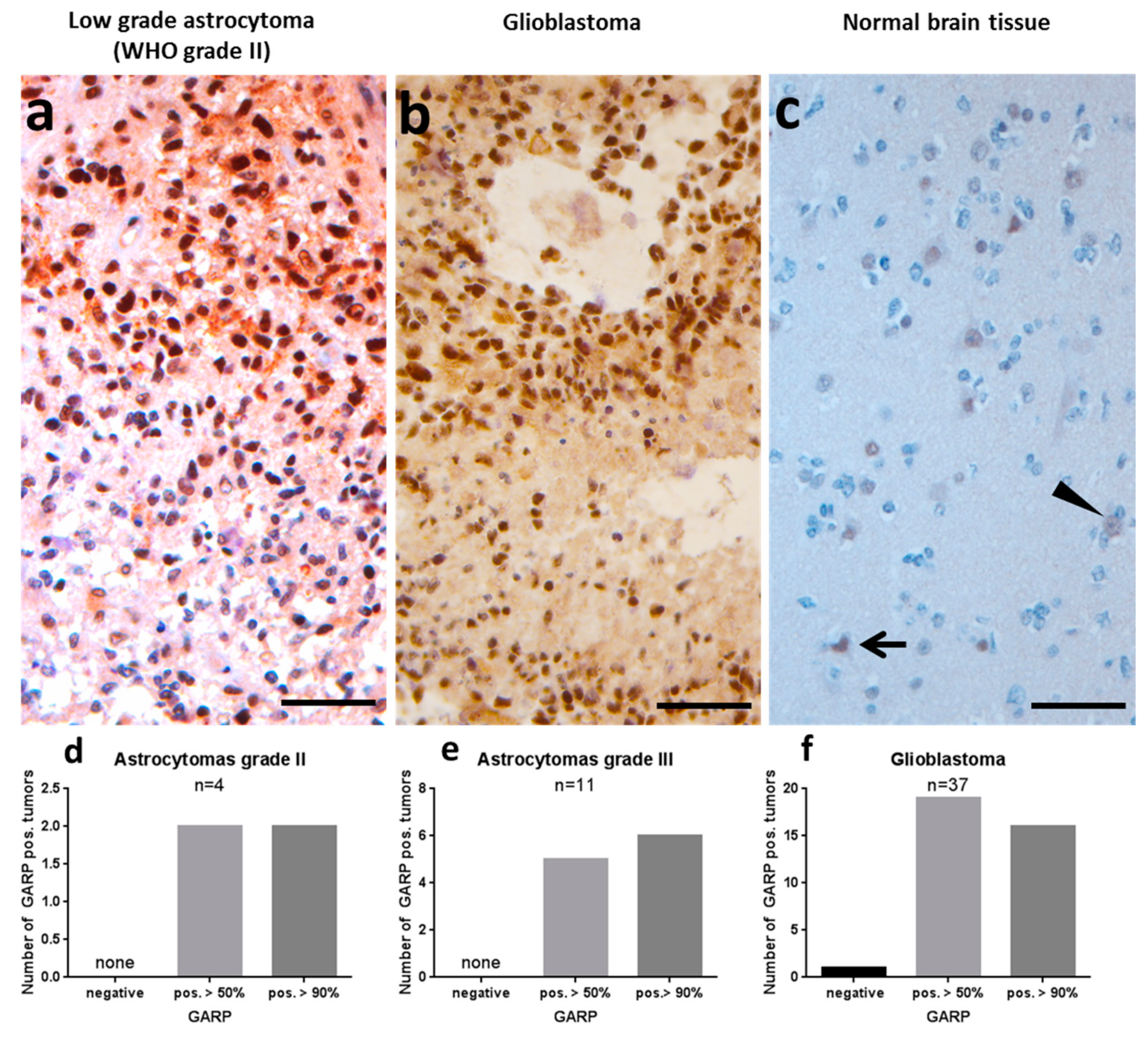
2.1. GARP Expression on Immunohistochemistry of Glioblastoma and Low-Grade Astrocytomas
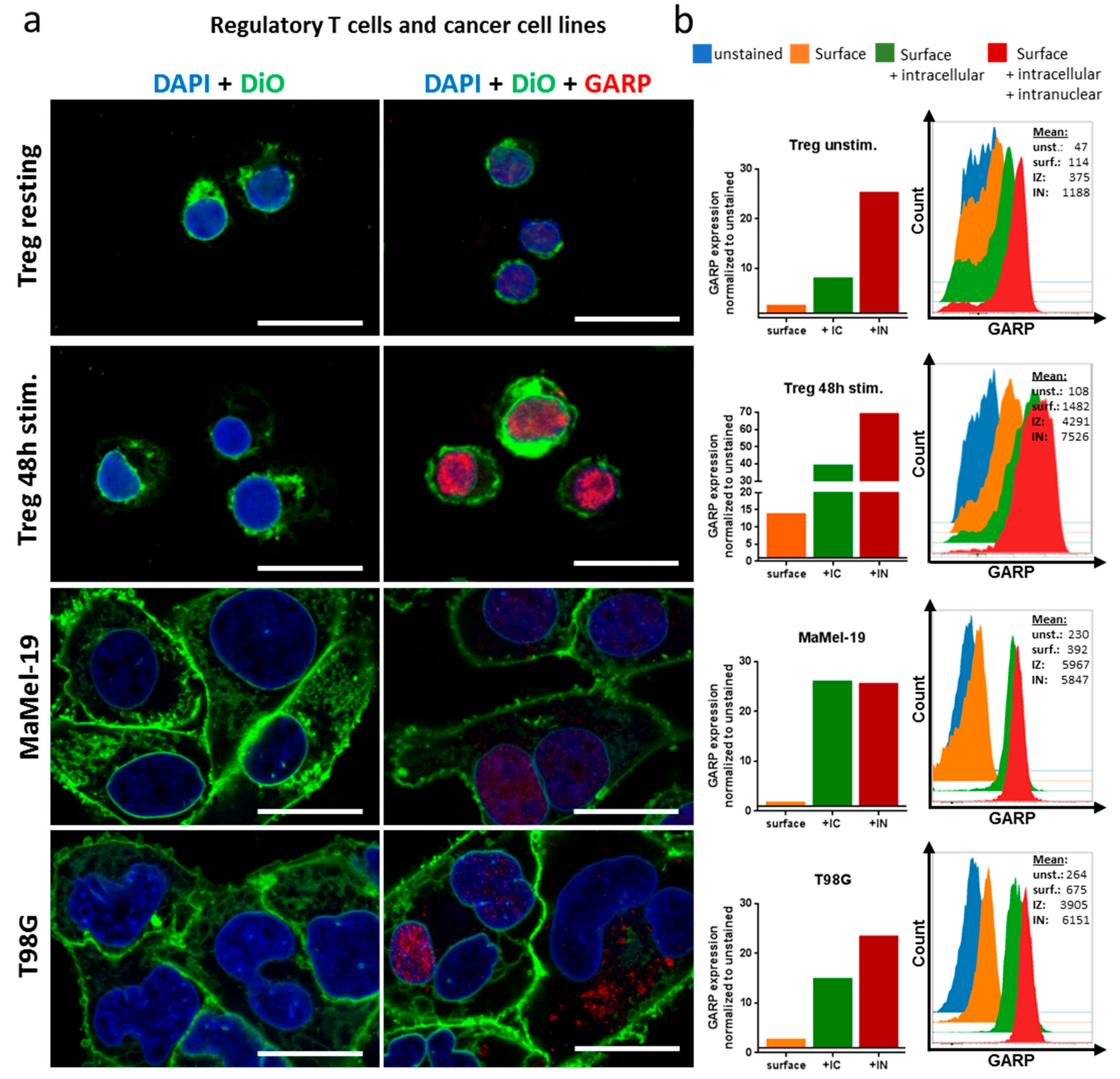
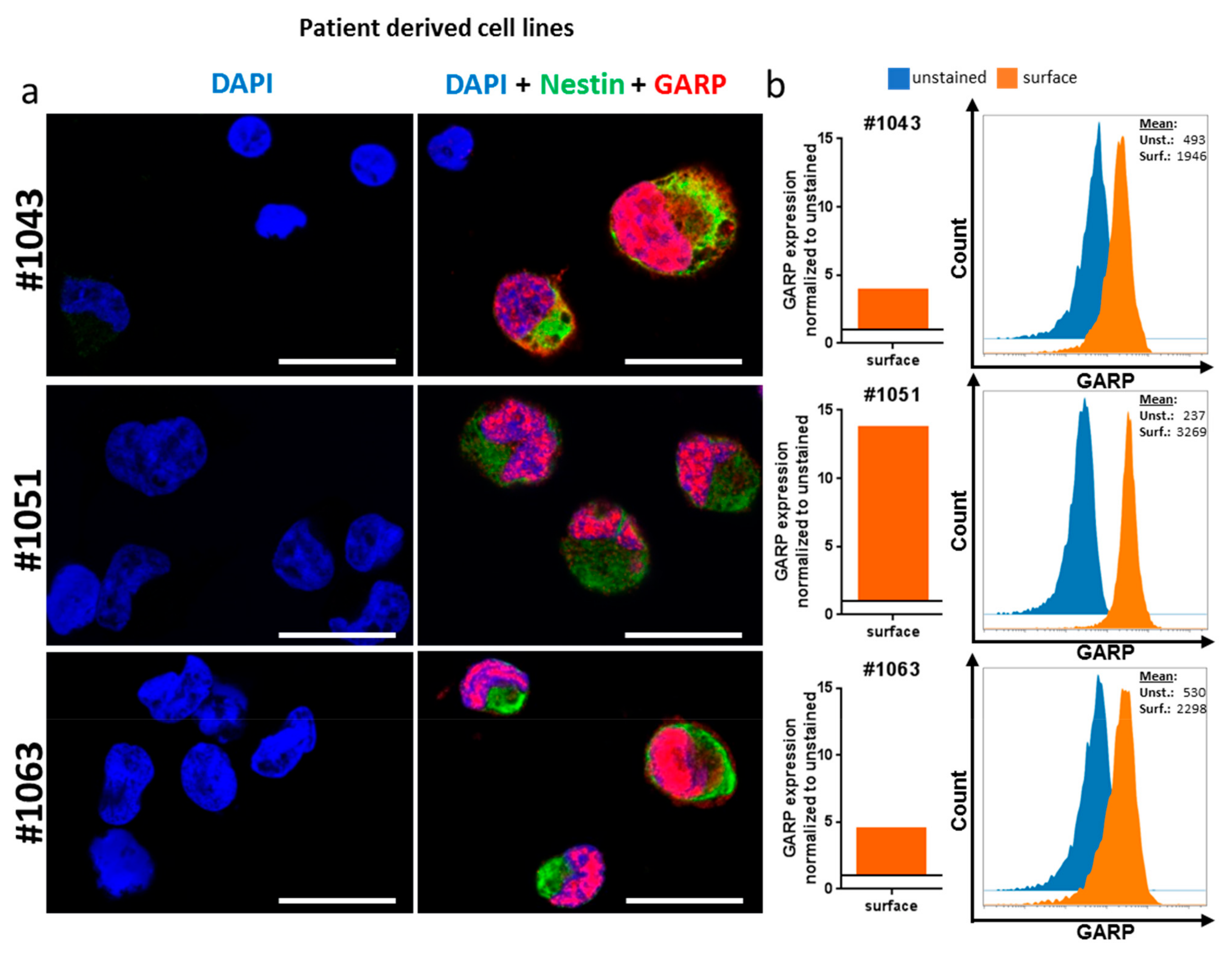
2.2. GARP Expressed on the Surface of GB and in the Cytoplasma and Nucleus
2.3. GB Cell Line T98G Suppresses T Effector Cell Function
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Isolation and Stimulation of Human CD4+ T Cells and Treg
4.3. Flow Cytometry
4.4. Confocal Microscopy
4.5. GARP-Immunohistochemistry
4.6. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Treg | Regulatory T cells |
| GARP | Glycoprotein A repetition domain |
| GB | Glioblastoma |
| IC | Intracellular |
| IN | Intranuclear |
| sGARP | Soluble GARP |
| tolDC | Tolerogenic dendritic cells |
| MDSC | Myeloid-derived suppressor cells |
| GSC | Glioma stem cells |
| TME | Tumor microenvironment |
| LRR | Leucine-rich domains |
| RTK | Tyrosine kinase receptors |
| PD-1 | Programmed death 1 |
| CTLA-4 | Cytotoxic T-lymphocyte associated antigen 4 |
| CFSE | Carboxyfluorescein succinimidyl ester |
References
- Quail, D.F.; Joyce, J.A. The Microenvironmental Landscape of Brain Tumors. Cancer Cell 2017, 31, 326–341. [Google Scholar] [CrossRef] [PubMed]
- Roesch, S.; Rapp, C.; Dettling, S.; Herold-Mende, C. When Immune Cells Turn Bad-Tumor-Associated Microglia/Macrophages in Glioma. Int. J. Mol. Sci. 2018, 19, 436. [Google Scholar] [CrossRef] [PubMed]
- See, A.P.; Parker, J.J.; Waziri, A. The role of regulatory T cells and microglia in glioblastoma-associated immunosuppression. J. Neurooncol. 2015, 123, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J.; van Waes, C.; Allen, C.T. Overcoming barriers to effective immunotherapy: MDSCs, TAMs, and Tregs as mediators of the immunosuppressive microenvironment in head and neck cancer. Oral Oncol. 2016, 58, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Hilf, N.; Kuttruff-Coqui, S.; Frenzel, K.; Bukur, V.; Stevanović, S.; Gouttefangeas, C.; Platten, M.; Tabatabai, G.; Dutoit, V.; van der Burg, S.H.; et al. Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature 2019, 565, 240–245. [Google Scholar] [CrossRef]
- Calinescu, A.-A.; Kamran, N.; Baker, G.; Mineharu, Y.; Lowenstein, P.R.; Castro, M.G. Overview of current immunotherapeutic strategies for glioma. Immunotherapy 2015, 7, 1073–1104. [Google Scholar] [CrossRef]
- Thomas, R.P.; Recht, L.; Nagpal, S. Advances in the management of glioblastoma: The role of temozolomide and MGMT testing. Clin. Pharmacol. 2013, 5, 1–9. [Google Scholar] [CrossRef]
- Sampson, J.H.; Maus, M.V.; June, C.H. Immunotherapy for Brain Tumors. J. Clin. Oncol. 2017, 35, 2450–2456. [Google Scholar] [CrossRef]
- Mangani, D.; Weller, M.; Roth, P. The network of immunosuppressive pathways in glioblastoma. Biochem. Pharmacol. 2017, 130, 1–9. [Google Scholar] [CrossRef]
- Lucca, L.E.; Hafler, D.A. Co-inhibitory blockade while preserving tolerance: Checkpoint inhibitors for glioblastoma. Immunol. Rev. 2017, 276, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Kesarwani, P.; Kant, S.; Prabhu, A.; Chinnaiyan, P. The interplay between metabolic remodeling and immune regulation in glioblastoma. Neuro-Oncology 2017, 19, 1308–1315. [Google Scholar] [CrossRef]
- Wesolowska, A.; Kwiatkowska, A.; Slomnicki, L.; Dembinski, M.; Master, A.; Sliwa, M.; Franciszkiewicz, K.; Chouaib, S.; Kaminska, B. Microglia-derived TGF-β as an important regulator of glioblastoma invasion—An inhibition of TGF-β-dependent effects by shRNA against human TGF-β type II receptor. Oncogene 2007, 27, 918. [Google Scholar] [CrossRef] [PubMed]
- Razavi, S.-M.; Lee, K.E.; Jin, B.E.; Aujla, P.S.; Gholamin, S.; Li, G. Immune Evasion Strategies of Glioblastoma. Front. Surg. 2016, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Perea, F.; Bernal, M.; Sánchez-Palencia, A.; Carretero, J.; Torres, C.; Bayarri, C.; Gómez-Morales, M.; Garrido, F.; Ruiz-Cabello, F. The absence of HLA class I expression in non-small cell lung cancer correlates with the tumor tissue structure and the pattern of T cell infiltration. Int. J. Cancer 2017, 140, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, T.F. Identifying and overcoming immune resistance mechanisms in the melanoma tumor microenvironment. Clin. Cancer Res. 2006, 12, 2326s–2330s. [Google Scholar] [CrossRef]
- Nduom, E.K.; Weller, M.; Heimberger, A.B. Immunosuppressive mechanisms in glioblastoma. Neuro-Oncology 2015, 17 (Suppl. 7), vii9–vii14. [Google Scholar] [CrossRef]
- Hahn, S.A.; Stahl, H.F.; Becker, C.; Correll, A.; Schneider, F.-J.; Tuettenberg, A.; Jonuleit, H. Soluble GARP has potent antiinflammatory and immunomodulatory impact on human CD4⁺ T cells. Blood 2013, 122, 1182–1191. [Google Scholar] [CrossRef]
- Tran, D.Q.; Andersson, J.; Wang, R.; Ramsey, H.; Unutmaz, D.; Shevach, E.M. GARP (LRRC32) is essential for the surface expression of latent TGF-beta on platelets and activated FOXP3+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 13445–13450. [Google Scholar] [CrossRef]
- Wang, R.; Wan, Q.; Kozhaya, L.; Fujii, H.; Unutmaz, D. Identification of a regulatory T cell specific cell surface molecule that mediates suppressive signals and induces Foxp3 expression. PLoS ONE 2008, 3, e2705. [Google Scholar] [CrossRef]
- Hahn, S.A.; Neuhoff, A.; Landsberg, J.; Schupp, J.; Eberts, D.; Leukel, P.; Bros, M.; Weilbaecher, M.; Schuppan, D.; Grabbe, S.; et al. A key role of GARP in the immune suppressive tumor microenvironment. Oncotarget 2016, 7, 42996–43009. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.-K.; Shen, M.-F.; Yao, H.-W.; Liu, Y.-S. Enhanced Expression of TGFBI Promotes the Proliferation and Migration of Glioma Cells. Cell. Physiol. Biochem. 2018, 49, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Mani, J.; Kumar, P.; Haridas, S.; Upadhyay, P.; Bhaskar, S. Activation of anti-tumor immune response and reduction of regulatory T cells with Mycobacterium indicus pranii (MIP) therapy in tumor bearing mice. PLoS ONE 2011, 6, e25424. [Google Scholar] [CrossRef] [PubMed]
- Tucci, M.; Stucci, S.; Passarelli, A.; Giudice, G.; Dammacco, F.; Silvestris, F. The immune escape in melanoma: Role of the impaired dendritic cell function. Expert Rev. Clin. Immunol. 2014, 10, 1395–1404. [Google Scholar] [CrossRef] [PubMed]
- Umansky, V.; Sevko, A. Melanoma-induced immunosuppression and its neutralization. Semin. Cancer Biol. 2012, 22, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Roncarolo, M.G.; Gregori, S.; Battaglia, M.; Bacchetta, R.; Fleischhauer, K.; Levings, M.K. Interleukin-10-secreting type 1 regulatory T cells in rodents and humans. Immunol. Rev. 2006, 212, 28–50. [Google Scholar] [CrossRef]
- Vermeersch, E.; Denorme, F.; Maes, W.; de Meyer, S.F.; Vanhoorelbeke, K.; Edwards, J.; Shevach, E.M.; Unutmaz, D.; Fujii, H.; Deckmyn, H.; et al. The role of platelet and endothelial GARP in thrombosis and hemostasis. PLoS ONE 2017, 12, e0173329. [Google Scholar] [CrossRef]
- Ollendorff, V.; Noguchi, T.; de Lapeyriere, O.; Birnbaum, D. The GARP gene encodes a new member of the family of leucine-rich repeat-containing proteins. Cell Growth Differ. 1994, 5, 213–219. [Google Scholar]
- Bencheikh, L.; Diop, M.B.K.; Rivière, J.; Imanci, A.; Pierron, G.; Souquere, S.; Naimo, A.; Morabito, M.; Dussiot, M.; de Leeuw, F.; et al. Dynamic gene regulation by nuclear colony-stimulating factor 1 receptor in human monocytes and macrophages. Nat. Commun. 2019, 10, 1935. [Google Scholar] [CrossRef]
- de Groot, J.F.; Piao, Y.; Tran, H.; Gilbert, M.; Wu, H.-K.; Liu, J.; Bekele, B.N.; Cloughesy, T.; Mehta, M.; Robins, H.I.; et al. Myeloid biomarkers associated with glioblastoma response to anti-VEGF therapy with aflibercept. Clin. Cancer Res. 2011, 17, 4872–4881. [Google Scholar] [CrossRef]
- Ilhan-Mutlu, A.; Wagner, L.; Wöhrer, A.; Jungwirth, S.; Marosi, C.; Fischer, P.; Preusser, M. Blood alterations preceding clinical manifestation of glioblastoma. Cancer Investig. 2012, 30, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Martens, A.; Wistuba-Hamprecht, K.; Geukes Foppen, M.; Yuan, J.; Postow, M.A.; Wong, P.; Romano, E.; Khammari, A.; Dreno, B.; Capone, M.; et al. Baseline Peripheral Blood Biomarkers Associated with Clinical Outcome of Advanced Melanoma Patients Treated with Ipilimumab. Clin. Cancer Res. 2016, 22, 2908–2918. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Cloughesy, T.; Perry, J.R.; Wick, W. Standards of care for treatment of recurrent glioblastoma—Are we there yet? Neuro-Oncology 2013, 15, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Seystahl, K.; Wick, W.; Weller, M. Therapeutic options in recurrent glioblastoma—An update. Crit. Rev. Oncol. Hematol. 2016, 99, 389–408. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.V.; Somani, A.-K.; Young, A.B.; Massari, J.V.; Ohtola, J.; Sugiyama, H.; Garaczi, E.; Babineau, D.; Cooper, K.D.; McCormick, T.S. Signal peptide cleavage is essential for surface expression of a regulatory T cell surface protein, leucine rich repeat containing 32 (LRRC32). BMC Biochem. 2011, 12, 27. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Alvarez-Breckenridge, C.A.; Wang, Q.-E.; Yu, J. TGF-β signaling and its targeting for glioma treatment. Am. J. Cancer Res. 2015, 5, 945–955. [Google Scholar] [PubMed]
- Lim, M.; Xia, Y.; Bettegowda, C.; Weller, M. Current state of immunotherapy for glioblastoma. Nat. Rev. Clin. Oncol. 2018, 15, 422–442. [Google Scholar] [CrossRef]
- Xue, S.; Hu, M.; Iyer, V.; Yu, J. Blocking the PD-1/PD-L1 pathway in glioma: A potential new treatment strategy. J. Hematol. Oncol. 2017, 10, 81. [Google Scholar] [CrossRef]
- Barrantes-Freer, A.; Kim, E.; Bielanska, J.; Giese, A.; Mortensen, L.S.; Schulz-Schaeffer, W.J.; Stadelmann, C.; Brück, W.; Pardo, L.A. Human glioma-initiating cells show a distinct immature phenotype resembling but not identical to NG2 glia. J. Neuropathol. Exp. Neurol. 2013, 72, 307–324. [Google Scholar] [CrossRef]
- Barrantes-Freer, A.; Renovanz, M.; Eich, M.; Braukmann, A.; Sprang, B.; Spirin, P.; Pardo, L.A.; Giese, A.; Kim, E.L. CD133 Expression Is Not Synonymous to Immunoreactivity for AC133 and Fluctuates throughout the Cell Cycle in Glioma Stem-Like Cells. PLoS ONE 2015, 10, e0130519. [Google Scholar] [CrossRef]
- Fischer, U.; Kim, E.; Keller, A.; Meese, E. Specific amplifications and copy number decreases during human neural stem cells differentiation towards astrocytes, neurons and oligodendrocytes. Oncotarget 2017, 8, 25872–25884. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kotecha, N.; Krutzik, P.O.; Irish, J.M. Web-based analysis and publication of flow cytometry experiments. Curr. Protoc. Cytom. 2010, 53. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glioma Grade III and IV | Male | Female | Total | Lost To Follow Up |
|---|---|---|---|---|
| Number of patients | 26 | 11 | 37 | |
| Age at the onset mean±SD (yr.) | 69.05 ± 11.08 | 71.38 ± 11.72 | 68.78 ± 13.36 | |
| Side hemispheric | ||||
| right | 9 | 3 | 12 | |
| left | 17 | 8 | 25 | |
| bilateral | 0 | 0 | 0 | |
| Surgery | ||||
| resection | 19 | 3 | 27 | |
| biopsy | 7 | 8 | 10 | |
| First histological diagnosis | ||||
| grade IV | 23 | 10 | 33 | |
| grade III | 3 | 1 | 4 | |
| Localization | ||||
| frontal | 5 | 5 | 10 | |
| parietal | 6 | 2 | 8 | |
| temporal | 9 | 2 | 11 | |
| thalamic | 2 | 0 | 2 | |
| fronto-parietal | 0 | 1 | 1 | |
| occipital | 2 | 0 | 2 | |
| perieto-occipital | 0 | 1 | 1 | |
| temporo-parietal | 2 | 0 | 2 | |
| Radiation therapy | 20 | 9 | 29 | 3 |
| Chemotherapy | 20 | 6 | 26 | 5 |
| Survival mean ± SD (mon.) | 11.68 ± 15.22 | 9.38 ± 4.98 | 11.07 ± 13.27 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimmer, N.; Kim, E.; Schupp, J.; Sprang, B.; Leukel, P.; Khafaji, F.; Ringel, F.; Sommer, C.; Tuettenberg, J.; Tuettenberg, A. GARP as an Immune Regulatory Molecule in the Tumor Microenvironment of Glioblastoma Multiforme. Int. J. Mol. Sci. 2019, 20, 3676. https://doi.org/10.3390/ijms20153676
Zimmer N, Kim E, Schupp J, Sprang B, Leukel P, Khafaji F, Ringel F, Sommer C, Tuettenberg J, Tuettenberg A. GARP as an Immune Regulatory Molecule in the Tumor Microenvironment of Glioblastoma Multiforme. International Journal of Molecular Sciences. 2019; 20(15):3676. https://doi.org/10.3390/ijms20153676
Chicago/Turabian StyleZimmer, Niklas, Ella Kim, Jonathan Schupp, Bettina Sprang, Petra Leukel, Fatemeh Khafaji, Florian Ringel, Clemens Sommer, Jochen Tuettenberg, and Andrea Tuettenberg. 2019. "GARP as an Immune Regulatory Molecule in the Tumor Microenvironment of Glioblastoma Multiforme" International Journal of Molecular Sciences 20, no. 15: 3676. https://doi.org/10.3390/ijms20153676
APA StyleZimmer, N., Kim, E., Schupp, J., Sprang, B., Leukel, P., Khafaji, F., Ringel, F., Sommer, C., Tuettenberg, J., & Tuettenberg, A. (2019). GARP as an Immune Regulatory Molecule in the Tumor Microenvironment of Glioblastoma Multiforme. International Journal of Molecular Sciences, 20(15), 3676. https://doi.org/10.3390/ijms20153676