Roles of MicroRNAs in Establishing and Modulating Stem Cell Potential


School of Life Science and Technology, ShanghaiTech University, 230 Haike Rd, Shanghai 201210, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(15), 3643; https://doi.org/10.3390/ijms20153643
Submission received: 30 June 2019
/
Revised: 18 July 2019
/
Accepted: 22 July 2019
/
Published: 25 July 2019
(This article belongs to the Special Issue MicroRNAs and Stem Cells)
Abstract
:Early embryonic development in mammals, from fertilization to implantation, can be viewed as a process in which stem cells alternate between self-renewal and differentiation. During this process, the fates of stem cells in embryos are gradually specified, from the totipotent state, through the segregation of embryonic and extraembryonic lineages, to the molecular and cellular defined progenitors. Most of those stem cells with different potencies in vivo can be propagated in vitro and recapitulate their differentiation abilities. Complex and coordinated regulations, such as epigenetic reprogramming, maternal RNA clearance, transcriptional and translational landscape changes, as well as the signal transduction, are required for the proper development of early embryos. Accumulated studies suggest that Dicer-dependent noncoding RNAs, including microRNAs (miRNAs) and endogenous small-interfering RNAs (endo-siRNAs), are involved in those regulations and therefore modulate biological properties of stem cells in vitro and in vivo. Elucidating roles of these noncoding RNAs will give us a more comprehensive picture of mammalian embryonic development and enable us to modulate stem cell potencies. In this review, we will discuss roles of miRNAs in regulating the maintenance and cell fate potential of stem cells in/from mouse and human early embryos.
1. Introduction
In mammals, early embryonic development can be divided into two stages, the pre-implantation stage and the post-implantation stage. During the preimplantation stage, zygotes go through multiple rounds of cell divisions, forming mature blastocysts which contain three compartments: the epiblast (EPI), the primitive endoderm (PrE), and the trophectoderm (TE), with distinctive developmental potential to give rise to the embryo, the yolk sac, and the placenta, respectively [1,2]. After implantation, the EPI, PrE, and TE are further specified to form embryonic and extraembryonic tissues. Also, shortly after implantation, a crucial type of stem cells, primordial germ cells (PGCs), is specified for the fate of gametes, initiating the next round of the life cycle. The whole process, albeit complex, can be viewed as a cascade of specification, with continuous self-renewal and differentiation of stem cells with various cell fate potential. After decades of efforts, different stem cells from both pre-implantation and post-implantation embryos can be cultured in vitro and still maintain their differentiation potential in vivo. Both animal and cell culture models greatly facilitate our understanding of key events during this earliest stage of life.
Noncoding RNAs can modulate gene expression through different mechanisms [3,4,5]. They are grossly classified by their sizes as short (19–31 nucleotides [nt]), midsize (~20–200 nt), and long (>200 nt) noncoding RNAs. Among short noncoding RNAs, microRNAs (miRNAs), endogenous small-interfering RNAs (endo-siRNAs), and PIWI-interacting RNAs (piRNAs) are the three classes most extensively studied [6]. miRNAs and endo-siRNAs share some common cellular machineries for biogenesis and use similar effector protein complexes for post-transcriptional silencing of specific genes [3]. In mammals, miRNAs are able to regulate almost all biological processes, whereas functions of endo-siRNAs are relatively unknown [4,7]. Substantial evidence demonstrates that miRNAs and endo-siRNAs participate in early embryogenesis in mice. Both in vitro and in vivo models suggest that miRNAs exert biological effects through regulating the self-renewal and differentiation of stem cells in/from early embryos. Since functions of miRNAs in pluripotent stem cells are well-covered elsewhere [8], in this review, we will focus on comparing in vivo and in vitro roles of miRNAs in regulating the potential of stem cells during early embryogenesis in mice and humans.
2. Biogenesis of miRNAs and siRNAs
MicroRNAs (miRNAs) are short non-coding RNAs, ranging from 22 to 24 nt in length, that repress gene expression at the post-transcriptional level in plants and animals [4]. In animals, miRNAs participate in a variety of biological processes, including the development and diseases [9]. miRNAs are typically transcribed by RNA polymerase II as primary miRNAs (pri-miRNAs), ranging from hundreds to thousands of nucleotides [10] (Figure 1A). Pri-miRNAs can either be monocistronic or polycistronic, encoding multiple miRNAs in the same transcript (Figure 1A). Mammalian pri-miRNAs are methylated by methyltransferase-like 3 (Mettl3) [11] and processed into 60–70 nt precursor miRNAs (pre-miRNAs) by microprocessor complexes, which consist of an RNase III enzyme, Drosha, and an RNA-binding protein, DiGeorge syndrome critical region gene 8 (Dgcr8) [12]. Dgcr8 binds to the pri-miRNA by recognizing an N6-methyladenylated GGAC motif [11], while Drosha cuts the pri-miRNA duplex at its hairpin structure [12]. Processed pre-miRNAs are then exported from the nucleus to the cytoplasm by the exportin 5 (Xpo5)/RanGTP complex [13,14] (Figure 1A). In the cytoplasm, pre-miRNAs are further processed into mature miRNA duplexes through removing terminal loops by another RNase III enzyme, Dicer [15] (Figure 1A). One of the mature miRNA strands (the guide strand) is then loaded onto Argonaute (Ago) protein and, together with other associated proteins, forms the RNA-induced silencing complex (RISC) [16]. In RISC, the miRNA recognizes its target mRNAs through base pairing between the short (6–8 nt) seed region of the miRNA and miRNA target sites on mRNAs [17,18] (Figure 1A). The RISC then either destabilizes target mRNAs or suppresses the translation of mRNAs, both of which lead to the post-transcriptional silencing [18,19]. Due to the partial complementarity and the short seed sequence, one miRNA is able to modulate the expression of hundreds of genes [20].
In addition to its role in making mature miRNAs, Dicer is also required for the maturation of short interfering RNAs (siRNAs). In eukaryotes, endogenous siRNAs (endo-siRNAs) can be generated from different sources of transcripts that form duplexes [21] (Figure 1B). In contrast to miRNAs, siRNAs are generated independently of microprocessors [22]. Another difference is the higher degree of complementarity between siRNAs and target mRNAs, which activates the slicing activity of Ago2 and leads to the cleavage of target mRNAs (Figure 1B). As a consequence, the silencing effect of siRNAs is usually considered to be stronger than that of miRNAs, although miRNAs can also perform potent inhibitory activities depending on the relative abundance between miRNAs and their targets [23,24]. Even though endo-siRNAs play important roles in heterochromatin formation and gene regulation in lower organisms [24], their functions in mammals are largely unknown and still remain to be investigated.
3. Development of Early Embryos in Mice and Humans
3.1. Formation of Gametes
In mice and humans, formation and development of germ cell lineages start with the specification of primordial germ cells (PGCs) in early embryos. In mice, PGCs can be identified as a cluster of ~40 cells at the base of the incipient allantois at ~E7.25 (embryonic day 7.25) [25,26]. PGCs then migrate along different compartments to reach genital ridges (precursors of gonads) at ~E10.5 [27,28,29,30], in where PGCs differentiate toward precursors for oocytes (oogenic pathway) or sperm (spermatogenic pathway). The initial step of the formation of oocytes, or oogenesis, is the formation of primary oocytes, which happens before or shortly after birth. After puberty, primary oocytes go through meiosis I to form secondary oocytes and first polar bodies. In vertebrates, secondary oocytes then progress through a part of meiosis II and arrest at metaphase II. After fertilization, meiosis II is completed, forming the mature 1N ovum (egg cell) and the secondary polar body. In both meiosis I and II, daughter cells are segregated in an asymmetric manner, leaving most of the cytoplasmic material in mature oocytes. In contrast, sperms are formed through two symmetric divisions of primary spermatocytes, followed by the generation of tailless spermatids. Spermatids then transform into sperms, losing most of its cytoplasm [31]. Thus, a fundamental difference between oocytes and sperms is the content of cytoplasmic materials. In mammals, a sperm carries only 10–20 fg RNA, while oocytes can carry 0.5–0.7 ng RNA [32]. Despite of the different RNA content, the relatively small amount of RNAs in sperms could still play important roles in fertilized embryos [33].
3.2. From Zygotes to Blastocysts
In mice, following fertilization, zygotes undergo three cleavages to form 8-cell embryos [34] (Figure 2). One major event, the maternal-to-zygotic transition (MZT), occurs during the first three cleavages. During the MZT, transcription from the zygotic genome is activated, known as the zygotic genome activation (ZGA), to support the subsequent embryonic development [35,36]. In mice, this process is further divided into the minor ZGA, which occurs in the middle S phase of the 1-cell embryo, and the major ZGA, which peaks between the 2-cell and 4-cell stage, characterized by a more extensive genomic reprogramming [37,38,39]. Another crucial event happens during the MZT is the clearance of maternal RNAs, which is triggered by the meiotic maturation of oocytes. By the 2-cell stage, 90% of maternal transcripts are degraded [40]. Failed clearance of maternal transcripts in mice led to the infertility, highlighting the importance of switching transcriptomes during the very early embryonic development [41,42].
Blastomeres of early 8-cell stage mouse embryos are still totipotent, capable of contributing to all embryonic and extra-embryonic cell lineages [43,44,45]. Subsequently, increased intercellular adhesion between blastomeres leads to the compaction, an apical-basal polarization appearing at the late 8-cell stage [45]. After compaction, several rounds of cleavage take place to form early blastocysts, which contain two distinct cell populations: the external layer of the embryo, or the trophectoderm (TE), gives rise to the placenta, while internal cells of the embryo form the inner cell mass (ICM) [34,45,46] (Figure 2B). At E4.5, the ICM subsequently segregates into the epiblast (EPI), which generates future cell lineages of the embryo proper, and the primitive endoderm (PrE), which gives rise to the yolk sac [34,44,45] (Figure 2C).
In humans, the minor ZGA occurs at the 2-cell stage [47,48], while the major wave of ZGA takes place between the 4-cell and 8-cell stage on embryonic day 3 (E3) [49]. Mid-preimplantation gene activation (MGA), peaking at the 8-cell stage, is also a period when maternally inherited RNAs and proteins are degraded [37,38,39]. Following the ZGA, the embryo undergoes compaction to form the morula that marks the first morphological indication of a break in radial symmetry, followed by the formation of the blastocyst between days 5 and 7 post-fertilization (Figure 2B). Despite the delayed timing, mature human blastocysts also consist of three lineages (EPI, PrE, and TE), whose developmental potential is considered to be in parallel to corresponding compartments in mice (Figure 2C).
3.3. Development after Implantation
Implantation, or the attachment of embryos to uteri, happens at E4.5 in mice, while human embryos undergo one additional round of cell division before implantation on day 7 [49]. In mice, the implanted embryo further elongates to form a radially symmetric egg cylinder containing the more specified epiblast (EPI) and the visceral endoderm (VE), derived from the PrE, as well as the extraembryonic ectoderm (ExE) and the ectoplacental cone, both of which are derived from the TE [50] (Figure 2D). During further development, the EPI gives rise to three germ layers (ectoderm, mesoderm, and endoderm) of the embryo, the extraembryonic mesoderm of the yolk sac, the amnion, as well as the allantois [51]. The VE will differentiate into the extraembryonic endoderm (visceral endoderm and parietal endoderm), which gives rise to the yolk sac (including visceral yolk sac and parietal yolk sac) [52,53] (Figure 2D). Finally, progenies of the TE, including the ExE and the ectoplacental cone, will generate trophoblast lineages of the placenta [54] (Figure 2D).
4. Stem Cells Derived from Early Embryos
Both mouse and human preimplantation embryos are composed of the epiblast (EPI), the primitive endoderm (PrE), and the trophectoderm (TE), which are able to give rise to the three germ layers of the fetus, the yolk sac, and the placenta, respectively [55]. After years of efforts, three in vitro cell culture models have been established to recapitulate potentials of those three distinctive lineages: embryonic stem cells (ESCs), extraembryonic endoderm (XEN) stem cells, and trophoblast stem cells (TSCs) [56,57,58,59,60] (Figure 2). There are two more types of stem cells that can be isolated shortly after implantation: the epiblast stem cell (EpiSC) which recapitulates the potential of the more primed epiblast (Figure 2E and see later), and the primordial germ cell (PGC), the very first precursor of the germ cell lineage [26,61,62] (Figure 2F). it is worthy of note that there are substantial differences in expression profiles of those three lineages of preimplantation embryos between mice and humans [63]. Thus, functions and regulatory mechanisms of those stem cells may not be completely conserved in those two species. Another thing that needs to be mentioned is that derived stem cells could be in a differentiated or de-differentiated state, owing to the signaling cues from the culture medium or feeder layers. In the next section, we will briefly discuss the in vitro models of stem cells in early embryos, particularly the differences between mice and humans, as well as different cell fate potential resulted from culture conditions.
4.1. Embryonic Stem Cells
Mouse ESCs (mESCs) were isolated from the ICM or EPI at E3.5 and E4.5 [56,64] (Figure 2). Initially, mESCs were cultured on mitomycin C-treated STO fibroblasts (feeder layers) with growth medium containing calf serum. Later, it was found that supplementing the leukemia inhibitory factor (LIF) inhibits embryonic stem cell differentiation [65]. mESCs recapitulate the pluripotent potential of the EPI and can differentiate into almost all cell types of three germ layers in vitro. Importantly, mESCs can form chimeras while being aggregated with or injected into the morula/blastocyst, demonstrating their pluripotency in vivo [66]. If the microinjection experiment is performed by using the 4N (tetraploid) recipient embryos, mESCs are even able to generate whole-ESC mice, since 4N cells can only form extraembryonic, but not embryonic, tissues, undeniably proving the full pluripotency of mESCs [67].
Human ESCs (hESCs) were first isolated from the ICM of the blastocyst produced by in vitro fertilization [58]. The primitive hESCs are maintained on irradiated mouse embryonic fibroblasts as feeder layers with the growth medium supplemented with fetal bovine serum [58]. Later, researchers demonstrated that a medium supplemented with basic FGF (bFGF) and BMP signaling inhibitor can sustain undifferentiated proliferation of hESCs in the absence of feeder layers or the conditioned medium [68]. Although mESCs and hESCs share similar embryonic origins, there are fundamental differences between mESCs and hESCs: (1) the morphology of hESC colonies is flattened, while mESC colonies are in a dome shape; (2) different pluripotency markers, such as SSEA-3 and SSEA-4, are expressed in hESCs, while SSEA-1 is expressed in mESCs; (3) the self-renewal of hESCs is dependent on FGF/TGFβ signaling pathways, while mESCs use LIF/BMP4 signaling pathways for their self-renewal [58,64,65,68,69,70]. These differences raise a question as to whether mouse and human ESCs are in equivalent states of cell potency. Actually, the morphology and the culture conditions of hESCs more resemble those of mouse epiblast stem cells (EpiSCs) derived from the post-implantation embryo (see later) [71,72], suggesting that hESCs cultured in the conditions mentioned above probably correspond to the EPI of the post-implantation embryo, which is more ready for lineage specification. This cell fate potential is coined the “primed” pluripotency state, in order to distinguish it from the “naïve” pluripotency state of the EPI in the pre-implantation embryo (Box 1).
Box 1. Culture conditions and pluripotent states of mESCs.
Efforts to optimize the culture condition for mESCs can be regarded as the process of figuring out signaling pathways for the self-renewal and differentiation of mESCs. For decades, mESCs were mostly cultured with calf serum on the feeder layer, supplemented by leukemia inhibitory factor (LIF). Further studies suggest that the serum can be replaced by bone morphogenetic proteins (BMPs) and the LIF/BMP combination is sufficient to sustain the self-renewal of ESCs [73]. LIF activates the Janus-associated kinase (JAK)/Stat3 pathway, as well as mitogen-activated protein kinase (Erk) cascade [74]. Strikingly, mESCs don’t require the mitogenic Erk signaling pathway for proliferation and, inversely, the Erk pathway actually promotes differentiation, as inhibiting the Erk pathway can sustain mESCs in the absence of LIF for a period of time [75,76]. In addition to the Erk cascade, activation of the Wnt signaling pathway by inhibiting of glycogen synthase kinase 3 (GSK3) also supports mESC self-renewal [77]. Through the combination of the Mek/Erk inhibitor, PD0325901, and the GSK3 inhibitor, CHIR99021, the Smith group figured out a condition (2i) for sustaining mESCs in a defined medium without the need for serum or feeder layers [78]. mESCs cultured in this medium are in a state termed “ground”, or “naïve”, since no external signaling is required for the maintenance of pluripotency, analogous to the EPI in the blastocyst. Although it is not essential, in practice, LIF is combined with 2i (2i/LIF) to promote the colony propagation of ESCs [76]. With the advent of 2i or 2i/LIF culture condition, the conventional serum/LIF/feeder culture is viewed as a condition leading to more heterogeneous mESCs, containing both the ground state population and the cells exited from the ground state [79].
If hESCs are actually in the primed pluripotency state, like mouse EpiSCs, is there a naïve state of hESCs that is the counterpart of mESCs? If there is, what could be the culture condition for it? Since 2010, several breakthrough studies have confirmed the existence of the naïve pluripotency state of hESCs using different approaches [80,81,82,83,84,85,86,87] (Figure 2). Some observations have been made that have indicated the naïve pluripotency of hESCs, such as the dome-like morphology, the activation of specific enhancer for Oct4, X chromosome reactivation (XaXa), higher mitochondrial respiration, expression of specific transposable elements, similar expression profiles compared to morula/early embryos, DNA hypomethylation, and the ability to form interspecies chimeras [80,81]. However, there is still no consensus as to whether all criteria should be applied to assess the naïve pluripotency, especially the chimera assay. It also needs to be noted that the genomic instability could be a problem for current conditions [88,89,90]. In summary, despite the great breakthrough, more studies are needed in order to understand naïve pluripotency in humans.
4.2. Extraembryonic Endoderm (XEN) Stem Cells
The extraembryonic endoderm (XEN) (stem) cell is another stem cell line derived from the mouse preimplantation blastocyst or the post-implantation embryo, representing the developmental potential of the PrE [57] (Figure 2). After implantation, the PrE segregates into two subpopulations, the parietal endoderm and the visceral endoderm (Figure 2), which will give rise to part of the parietal yolk sac during the early developmental stage and to the visceral yolk sac during the late developmental stage, respectively [91]. In the chimera assay, XEN cells contribute to both the parietal endoderm and the visceral endoderm, despite a strong tendency toward the former [92]. Different from the skewed potential in vivo, XEN cells can be differentiated into visceral endoderm-like cells in vitro [93,94,95].
Human XEN cells are not established, by any means. Again, this could be due to some fundamental differences in PrEs between mice and humans [96,97]. For example, mouse XEN cells can be converted from mESCs by ectopic expression of master transcription factors of the PrE, such as Gata6 or Sox17 [98]. Meanwhile, overexpression of Gata6 and Sox17 is sufficient to convert hESCs into a XEN-like morphology; such cells, however, cannot be maintained [99], suggesting that a different culture condition could be needed for deriving and culturing human XEN cells.
4.3. Trophoblast Stem Cells
Mouse trophoblast stem cells (mTSCs), established 17 years after the derivation of mESC, can be derived from the TE of the E3.5 blastocyst or the extraembryonic ectoderm (ExE) of the post-implantation embryo [60] (Figure 2). In vivo, mTSCs recapitulate the developmental potential of the TE, forming trophoblast lineages in the placenta of the chimeric embryo [60]. Yet, in contrast to well-developed procedures for differentiating mESCs into various cell types in vitro, ways of differentiating mTSCs into specific trophoblast lineages are still incomplete. Two of the most important mouse trophoblast lineages, trophoblast giant cells (TGCs) and syncytiotrophoblasts (SynTs), are able to differentiate from mTSCs. Nonetheless, protocols for generating homogenous TGCs/SynTs, as well as other trophoblast subtypes, such as spongiotrophoblasts, are still lacking. Considering the underestimated importance of placenta defects that can lead to embryonic lethal phenotypes [100], more investigations on mTSCs should be conducted to understand the development of extraembryonic tissues.
Derivation of human trophoblast stem cells (hTSCs) has been even more lagged, possibly due to the different gene expression profiles of TEs between mice and humans [101,102]. hTSCs can be derived from day 7 blastocysts or from first trimester (6–8 weeks) placenta [59] (Figure 2). Different from mTSCs, which rely on Fgf4 and conditioned medium (can be replaced by Tgfβ or activin) for the self-renewal, a much more complicated chemical cocktail is required for the derivation and maintenance of hTSCs [59]. hTSCs can be differentiated in vitro into two major types of human trophoblasts, the extravillous trophoblasts (EVTs) and the syncytiotrophoblasts (STBs) [59]. Even though hTSCs can differentiate into EVTs and STBs, whether it is equivalent to the human TE is still unclear, since cytotrophoblasts (CTBs) of human placenta can also give rise to those two trophoblast lineages. Apparently, more characterization is needed for this new stem cells, such as comparing the expression profile of the hTSC with the human TE, identifying the core regulatory circuitry for its self-renewal, as well as examining its potential to form interspecies chimeras.
4.4. Epiblast Stem Cells
Mouse epiblast stem cells (EpiSCs) are derived from the epiblast of the post-implantation embryo [71,72] (Figure 2). EpiSCs cultured in vitro represent the “primed” pluripotent state of epiblast in utero, forming derivatives of three germ layers but rarely incorporating into chimeras following blastocyst injection [71]. As aforementioned, primed hESCs cultured in the canonical condition share a lot of characteristics of mouse EpiSCs, such as morphology, expression of signature genes, signaling pathways for self-renewal, inactivation of one X chromosome (XaXi), and glycolytic metabolism, indicating they are in the same pluripotency state [69].
4.5. Primordial Germ Cells
In mice, primordial germ cells (PGCs) arise from post-implantation epiblasts (Figure 2). Primary mouse PGCs isolated from post-implantation embryos can be cultured after reprogrammed by bFGF, Kit ligand, and LIF as the format of embryonic germ (EG) cells, which are not identical to PGCs [103,104,105]. For humans, it is impracticable to isolate PGCs since they are specified at week 2–3 post-fertilization [106]. This difficulty is now circumvented by a breakthrough method that induces PGC-like cells (PGCLCs) from pluripotent stem cells in vitro [107,108,109,110]. In mice, PGCLCs can further differentiate to functional gametes, suggesting they’re functionally comparable to authentic PGCs [107,111]. A similar approach can also induce human PGCLCs, which acquires the germ cell fate [112]. Importantly, PGCLCs can only be differentiated from naïve pluripotent stem cells through an epiblast-like state (epiblast-like cells, or EpiLCs), a route recapitulating the specification of PGCs in utero [107,113] (Figure 2).
5. Roles of microRNAs in Early Embryos
5.1. Expression of miRNAs in Early Blastomeres
After fertilization, zygotes receive cytoplasmic materials from both sperms and oocytes. As mentioned above, most RNAs, including miRNAs, come from oocytes. The expression of miRNAs in mature oocytes and zygotes is similar, suggesting that zygotic miRNAs are mainly maternally inherited [114,115]. Whether specific miRNAs from sperms play important roles in zygotes for the subsequent development remains an open question. For example, two miRNA clusters, miR-34b/34c and miR-449a/b/c, are highly enriched in sperms but absent in oocytes [116]. Inhibition of miR-34c by injecting antagonistic linked nucleic acids (LNAs) into zygotes led to attenuated first cleavage of zygotes, suggesting that paternal miR-34c could participate in embryo development [117,118]. However, genetic ablation of miR-34b/c and miR-449a/b/c in mice draws a more complicate picture: knocking out both miR-34b/c and miR-449a/b/c (dKO) led to a severe defect of spermatogenesis. Yet, the dKO round spermatids, while being injected to oocytes, were able to fertilize oocytes and to support the normal embryo development, suggesting that deficiency of miR-34/449 only affects the development of sperms (the formation of tails) but does not influence the development of fertilized zygotes [116,118]. The discrepancy between those two experiments could be due to the off-target effect of LNAs. Even though miR-34b/c and miR-449a/b/c are dispensable for the development of mouse embryos, the miR-34c level in spermatozoa is corrected with the outcome of intracytoplasmic sperm injection (ICSI), suggesting miR-34c could be beneficial for the development of human embryos [119]. Moreover, besides of the canonical inhibitory mechanism through mRNA destabilization, paternally inherited miRNAs have been shown to play important roles in the epigenetic inheritance of zygotes [120,121].
After fertilization, the expressions of many miRNAs (mostly maternally inherited) are down-regulated more than two-fold during the oocyte-to-1-cell transition and the minor ZGA [39]. The most drastic change of total miRNA levels happens during the MZT, when total amount of miRNA is down-regulated by 60% [114]. The degradation of miRNAs is significantly slowed down since the MGA [39], suggesting that the de novo synthesis of miRNAs takes place between the 2-cell and 4-cell stage. Using a novel high throughput microarray assay, Yang and colleagues discovered 67 differentially expressed miRNAs classified into four stage-dependent groups: 7 miRNAs in oocytes, 7 miRNAs in 2-cell blastomeres, 25 miRNAs in 8-cell morulae, and 28 miRNAs in blastocysts [122]. The most abundant maternal miRNAs in zygotes are the let-7 and miR-17~92 miRNAs, whose expression are elevated during oogenesis and then inherited by zygotes [114]. The expression of let-7 and miR-17~92 is increased again after the 2-cell embryo stage in mice, correlating with the de novo biogenesis of miRNAs [114]. However, the most extensively up-regulated miRNAs in 4-cell blastomeres are the miR-290 miRNA cluster, miR-290~295 [123], whose expression is increased 15-fold and 24-fold at the 4-cell and 8-cell stage, respectively, compared to the 2-cell stage [114] (Figure 3). In humans, the majority of miRNAs detected in human oocytes are inherited by zygotes and significantly down-regulated in blastocysts, such as miR-31, miR-16, let-7a, miR-145, miR-210, and miR-212 [124,125]. One of the most up-regulated miRNAs in human blastocysts is miR-371~373, the human homologue of miR-290~295 cluster in mice [126] (Figure 3).
5.2. Functions of miRNAs in Pre-Implantation Embryos
Although global and specific changes of miRNA expression profiles seem to suggest their functional roles in embryo development, it is surprising that miRNAs may be dispensable for the embryonic development, at least before implantation [127,128,129]. General approaches to study functions of miRNAs is to deplete miRNA biogenesis proteins, such as Dicer, Drosha, or Dgcr8 (Figure 1). However, in pre-implantation embryos, depleting miRNAs in gametes and zygotes exhibited different phenotypes. For example, since dicer−/− mice are embryonic lethal [128,129,130], zygotic dicer−/− mice can only be made from intercrossing dicer+/- mice. In this crossing, dicer-deficient oocytes, which are generated from dicer+/− oogonia, will still contain Dicer in the cytoplasm, and this maternal Dicer will be inherited by dicer−/− zygotes, even though there will be no de novo synthesis of Dicer anymore. To completely deplete inherited Dicer in zygotes, a maternal knockout approach (for instance, using the Zona pellucida glycoprotein 3 promoter (Zp3)-Cre line expressing Cre in developing oocytes) is often employed [114,131,132]. For Dicer, the zygotic knockout resulted in the embryonic arrest at E7.5, with the normal blastocyst formation at E3.5, suggesting that the de novo synthesis of miRNAs is dispensable for blastocyst formation [128,129,130]. On the other hand, maternal depletion of Dicer in oocytes resulted in the meiotic arrest of oocytes with defects in spindle and chromosomal segregation [114,131,133] (Box 2). The maturation defect of oocytes in the dicer-deficient mouse model makes it difficult to study maternally inherited miRNAs in zygotes [114,131,133]. Eventually, the functional role of maternally inherited miRNAs was addressed by the maternal dgcr8 knockout: dgcr8-deficient oocytes mature normally and, when fertilized by dgcr8-deficient sperms (maternal-zygotic dgcr8−/− or MZ dgcr8−/− zygotes), are able to develop into normal blastocysts with the proper segregation of the TE and ICM identified by marker staining [133]. Although the observation above seems to suggest that Dgcr8-dependent miRNAs are dispensable for segregation of the EPI, PrE, and TE lineages, stem cells derived from those lineages exhibit defects in either self-renewal or differentiation, indicating possible discrepancies between in vitro and in vivo models [128,134,135,136,137] (see later). Since the segregation of three lineages in blastocysts is only confirmed by lineage markers, more detailed analyses, such as comparing expression profiles of three lineages between wildtype and MZ dgcr8−/− blastocysts, will help to decide if there is any defect at this stage. Also, it needs to note that some miRNAs are generated through Dgcr8- or even Dicer-independent pathway [22,138,139] (Figure 1A), and their possible roles in early embryonic development cannot be excluded. Nonetheless, despite of these caveats, current evidence tends to argue that miRNAs, either inherited or de novo synthesized, are not essential for the development up to the blastocyst stage.
Box 2. Function of Dicer in mouse oocytes.
Transposable elements (TEs) make ~40% of mammalian genomes. During gametogenesis, extensive epigenetic reprogramming, such as DNA demethylation, happens, leading to the reactivation of TEs. To maintain genomic integrity, mechanisms defending the genome of the germline are evolved by the host. One of them is a mechanism dependent on a class of small RNAs, PIWI (P-element-induced wimpy testis)-interacting RNAs (piRNAs) [140]. Generated through a Dicer-independent mechanism, piRNAs encoding TE sequences associate with PIWI proteins to silence TEs through degrading TE transcripts and/or silencing genomic loci of TEs [141,142]. In male mouse germline, three PIWI proteins, Mili, Miwi, and Miwi2, are expressed, and all of them play essential roles in spermatogenesis [143,144,145]. Knocking out PIWI proteins led to aberrant activation of TEs, causing spermatogenic arrest [146]. Interestingly, the piRNA pathway is dispensable in female mouse germline, possibly due to the lack of the fourth PIWI family member, PIWIL3, which exists in the oocytes of other mammals [147]. An alternative to compensate for the loss of the piRNA pathway and to silence TEs is the endo-siRNA pathway (Figure 2). It has been demonstrated that deficiency of Dicer in mouse oocytes leads to a decreased level of endo-siRNAs and upregulation of TEs [148,149]. Interestingly, an alternative isoform of Dicer, DicerO, is specifically expressed in oocytes, instead of the one expressed in the somatic cell, DicerS [150]. DicerO is more efficient at processing endo-siRNAs than miRNAs, and specifically knocking out dicerO is sufficient to phenocopy the MZ dicer knockout, leading to meiotically arrested oocytes with spindle defects [150]. Since dicero−/− oocytes only exhibit minor defects in the miRNA expression, and the miRNA activity is suppressed in mouse oocytes [150], the loss of endo-siRNAs could be responsible for the knockout phenotype. However, this doesn’t exclude the possibility that miRNAs may play roles in silencing TEs at later stages of embryonic development.
5.3. Functions of miRNAs in Post-Implantation Embryos
In contrast to pre-implantation embryos, roles of miRNAs in post-implantation embryos are more evident: zygotic depletion of dgcr8 or dicer in mice led to growth arrest between E6.5 and E7.5 [129,136]. In implanted zygotic dicer−/− embryos, the initial specification of the epiblast is normal compared with wildtype ones, despite the expression of miR-290 miRNAs (miR-291-3p, miR-295, and miR-291-5p) was not compromised in dicer−/− embryos for unknown reasons [128]. Defects of dicer−/− embryos only become prominent at the gastrulation stage, in the elongation of the primitive streak and in the specification of definitive endoderm [128]. Those defects are in parallel with differentiation defects in dicer- or dgcr8-deficient ESCs (see below). It remains to be decided whether defects in embryonic development happen during the transition from the naïve to the primed state of EPI, or during the lineage specification from a normally primed epiblast. Since Dicer controls the biogenesis of both miRNAs and endo-siRNAs, dgcr8 knockout will be a more appropriate model to perform more detailed analyses. Moreover, since the miR-290 cluster is still expressed in dicer−/− embryos, double knockout of dicer and miR-290 may elucidate the role of miRNAs in post-implantation embryos.
In addition to the essential roles of miRNAs in epiblast development, miRNAs are also required for the development of extraembryonic lineages. In dicer−/− post-implantation embryos, the number of trophoblast stem cells in the ExE, the derivative of the TE lineage, was reduced at E6.5 [128]. Also, the patterning of the VE, the derivative of the PrE lineage, was also severely compromised [128]. Importantly, conditional knockout of dicer in the epiblast using the Sox2Cre line extended the survival of post-implantation embryos up to E9.5 with normal extraembryonic tissues, suggesting that defects of extraembryonic lineages in post-implantation embryos are not due to defects in epiblast development. Contrarily, defects in extraembryonic lineages could be among the causes for the failure of primitive streak elongation [128]. At this stage, apoptotic cells are greatly increased in dicerepi−/− embryos, suggesting the main role of miRNAs (or endo-siRNAs) in embryonic tissues could be protecting the cells from dying upon specification [128]. In addition, these results also highlighted the importance of miRNAs in the specification of extraembryonic lineages in utero. With the advent of the era of single-cell transcriptomics, tracing defects of various cell subtypes in dicer−/− or dgcr8−/− embryos, including both embryonic and extraembryonic lineages, will help to address roles of miRNAs in the development of post-implantation embryos.
Knocking out dicer or dgcr8 leads to, in most cases, the most severe phenotype since the majority of miRNAs were ablated. In dicer- or dgcr8-deficient mouse model, it is usually a hard task to figure out which miRNA(s) is responsible for a specific phenotype. Actually, most miRNA-knockout mouse strains are either viable or without obvious phenotypes [7] (Table 1). The miR-290 cluster is the best guess, since it is highly upregulated during the early embryonic development and is also highly enriched in mESCs [123,151] (Figure 3). Surprisingly, knockout of miR-290 only resulted in partially embryonic lethality after E8.5 [126] (Table 1). Around 50~60% of miR-290−/− embryos exhibit two types of developmental defects, the localization of embryos outside of the yolk sac, as well as delayed development (e.g., fewer somites) of embryos [126]. Considering the delayed phenotypes compared with dicer−/− or dgcr8−/− embryos, it cannot be excluded that defects of miR-290−/− embryos could be due to the abnormal development of extraembryonic tissues. Actually, a recent study demonstrated that miR-290 is highly expressed throughout embryogenesis from E2.5 to E6.5, disappears in the embryo proper at E7.5, but remains highly expressed in the yolk sac and placenta [152]. Knocking out miR-290 resulted in the loss of trophoblast progenitors, the reduced-size placenta, and the defect in the maternal-fetal transport [152] (Table 1). Taken together, those results suggest that miR-290 deficiency recapitulates phenotypes of dicer/dgcr8 knockout more in extraembryonic tissues, rather than in the epiblast. Since the development of embryos and extraembryonic tissues are coordinated, tissue-specific knockouts in extraembryonic lineages will be helpful to clarify the complexity.
5.4. miRNA Activities during Early Development
Why are miRNAs, no matter whether inherited or de novo synthesized, present but do not seem to be functional during the early embryonic development until the blastocyst stage? One possibility is that the expression of miRNA biogenesis proteins, such as Drosha, Dgcr8, Xpo5, and Dicer, are drastically downregulated during zygotic cleavages and their expression levels remain low until the blastocyst stage [164]. The lack of miRNA biogenesis explains why zygotic knockout of dicer or dgcr8 does not exhibit phenotypes up to the blastocyst stage since the biogenesis of miRNAs is already low even in wildtype embryos. However, low expression levels of miRNA biogenesis proteins may not be the rate-limiting factor for miRNA maturation, since some miRNAs, such as the miR-29b and miR-290 clusters, have been found to be expressed at the 2-cell, 4-cell, and 8-cell stage [114,165]. A complete survey of 238 miRNAs during early development of mouse embryos also revealed several miRNA expression patterns and some miRNAs are up-regulated at the early embryonic stage [118]. In summary, it is still unclear about the amount and dynamic of miRNAs at each embryonic stage. A landscape of miRNA expression profiles at each stage of early embryonic development will be crucial to addressing this question.
The following question is, even if mature miRNAs are made in early embryos, are they functional? In other words, does the miRNA effector machinery work in early embryos? Interestingly, there is clear evidence demonstrating that the miRNA activity is strongly suppressed in mouse oocytes, since wildtype and dgcr8−/− oocytes exhibited basically identical mRNA expression profiles, and miRNA activity is low in oocytes, as demonstrated by the miRNA reporter assay [132,133]. Suppression of miRNA activity is attributed to the expression of an alternative Ago2 isoform, which lacks all known functional domains [166]. The inhibitory effect might be relieved after the transcription of normal ago2 at the 4-cell and 8-cell stage. Thus, examining miRNA activities at different embryonic stages using reporters will help to clarify this issue [132].
6. Functions of miRNAs in Stem Cells
Two important features of pluripotent stem cells are self-renewal and differentiation. Regular in vitro differentiation methods for testing the potency of pluripotent stem cells includes the withdrawal of LIF and providing cues for the specification toward specific germ layers and their progenies. However, with more understanding of different states of pluripotency, the roles of miRNAs in pluripotent stem cells should be addressed with more cautiousness. For example, differentiation defects of ESCs could be due to (1) the naïve-to-prime transition of ESCs, (2) defects in entering/exiting an intermediate state such as the formative state [79], or (3) the lineage specification process (differentiation) of primed ESCs. Indeed, recent evidence suggests that some differentiation defects of ESCs could be due to the naïve-to-prime transition. Here, we discuss recent findings on functions of miRNAs in ESCs of different pluripotent states, in TSCs, in XEN cells, and in PGCs.
6.1. Functions of miRNAs in the Expanded Pluripotency State
Conventional pluripotent stem cells, by definition, are only capable of generating the embryonic portion, forming derivatives of three germ layers. However, blastomeres at the earlier embryonic stage, such as the 2-cell stage in mice, exhibit a totipotent potential, giving rise to both embryonic and extraembryonic tissues [167]. So far, no in vitro cell culture models recapitulate the “authentic” totipotency of early blastomeres in vivo, which, by themselves, are able to give rise to the whole embryo. Yet, several recent studies indicate that pluripotent stem cells can be converted to and stably maintained in a state with expanded potential and, while being injected into early mouse embryos, are able to form both embryonic and extraembryonic tissues in chimeras [168,169,170,171]. Choi and colleagues found that knockout of miR-34 in embryonic stem cells resulted in a “bi-directional” cell fate potential, giving rise to both embryonic and extraembryonic lineages in chimeras [168]. Subsequent studies suggested that miR-34 blocks the transition from the pluripotency state toward the bi-directional state [168]. Deficiency of miR-34a led to the activation of the MuERV-L endogenous retroviruses, a feature shared by totipotent 2-cell blastomeres [168]. Interestingly, the level of miR-34 also increases during the formation of blastocysts, suggesting that miR-34 may also restrict the totipotency and play a role in the segregation of the EPI/PrE/TE lineages in vivo [168]. Since the development and the formation of chimeras could be two different processes, more studies are needed to elucidate the role of miR-34 in the development of early embryos.
6.2. Functions of miRNAs in the Naïve Pluripotency State
Mouse ESCs represent the naïve cell fate potential of the ICM/EPI. However, there are differences between in vivo and in vitro model [134,136]. Both dicer−/− and dgcr8−/− ESCs maintain the normal ESC morphology, express pluripotent marker genes, but exhibit defects in proliferation due to the G1 phase accumulation [134,136]. In contrast, no obvious defects were observed in dicer−/− post-implantation embryos until E5.5, suggesting there are no discernible defects in the proliferation and differentiation of epiblasts until then [128]. Also, while differentiating into embryoid bodies (EBs), dicer−/− ESCs failed to express either endodermal (hnf4) or the mesodermal (brychyury, bmp4, and gata1) markers [134]. In contrast, careful examination of embryos with whole-body or epiblast-specific depletion of dicer revealed that there are no defects in the induction of brychyury (the initial formation of the primitive streak) or the formation of visceral endoderm [128]. In aggregate, there are evident differences in both cell proliferation and differentiation between pluripotent stem cells in vitro and in vivo. There are several explanations for the differences between dicer- and dgcr8-deficient ESCs and embryos. First, the maternal Dicer could compensate the loss of zygotic Dicer, enabling the dicer−/− epiblast to proliferate and differentiate to certain extend. Second, both dicer−/− and dgcr8−/− ESCs are difficult to obtain since the escape from the initial proliferation arrest may be required for the derivation [134,136]. Hence, some properties of dicer−/− or dgcr8−/− ESCs could be altered during derivation and are no longer identical to the dicer−/− or dgcr8−/− EPI. Third, in vitro differentiation conditions are likely not able to recapitulate the elegantly controlled signal sequences during the development. Even though the roles of miRNAs in proliferation seem to be different in vitro and in vivo (despite the complexity due to maternal Dicer), the lethality of dicer−/− and dgcr8−/− embryos at the gastrulation stage indicates that miRNAs do participate in the post-implantation development, which can be studied using dicer−/− or dgcr8−/− ESCs as differentiation models.
The miRNA profiling has been performed to study the function of miRNAs in mESCs [171]. Two miRNA clusters are dominantly expressed in naïve mESCs: miR-290~295 and miR-17~92 [172] (Figure 3A,C,D). Two paralogs of miR-17~92, miR-106a~363 and miR-106b~25, are also expressed in mESCs at the lower level [171] (Figure 3C,D). miR-290~295, a polycistronic miRNA containing six miRNAs, harbors the seed sequence AAGUGC which is also shared by another miRNA cluster, miR-302~367, which is also expressed in mESCs [173] (Figure 3B). Consistent with the same potential state, the human analog of miR-290~295, miR-371~373, is also highly expressed in naïve hESCs [174] (Figure 3A). Surprisingly, despite their high abundance, single knockout of miR-290~295 or double knockout of miR-290~295 and miR-302~367 in mESCs does not influence the expression of pluripotency genes, suggesting that knocking out those miRNAs does not phenocopy dicer or dgcr8 knockout that compromises the self-renewal of mESCs [126,175] (Table 2). It remains to be studied whether human miR-371~373 and/or miR-302~367 are also dispensable for the pluripotency or proliferation of naïve hESCs. Interestingly, in mice, triple-knockout of three other miRNAs (miR-17~92, miR-106a~363, and miR-106b~25) with the abundance secondary to miR-290~295 exhibits no effect on the embryonic development until E15 [153] (Table 1). Despite the laborious work, knockout of multiple miRNAs, including the miR-290 family and the miR-17~92 family, will help to answer whether there is functional redundancy of these miRNAs.
Another miRNA cluster, although it is not highly expressed, is actually important in regulating the stemness of ESCs. let-7 family miRNAs are highly redundant, containing 10 subfamilies distributed at 13 loci in mice and humans. Expression of mature let-7 is low in mESCs due to the antagonistic effect of Lin28. Lin28 proteins, including Lin28a and Lin28b, selectively bind to the loop of pre- and pri-let-7 and block their processing by Dicer and Drosha, respectively [186,187,188,189]. On the other hand, let-7 also targets lin28a and lin28b, forming negative feedback loops [190]. In general, let-7 is highly expressed in somatic tissues but absent in many types of stem cells, suggesting its role in promoting differentiation [180,191,192]. Overexpression of let-7 suppressed the self-renewal of dgcr8−/− ESCs, but not wildtype ESCs alone (with normal miR-290 expression) or dgcr8−/− ESCs that co-introduced with miR-290 miRNAs, suggesting that let-7 and miR-290 miRNAs play opposing roles in maintaining the self-renewal of mESCs [180] (Table 2). Subsequent analyses indicate that let-7 targets hundreds of genes within the pluripotency network, including sall4, nmyc, lin28, and many other targets that are indirectly positively regulated by miR-290 miRNAs [180]. Thus, decreased miR-290 and increased let-7 could be essential for the exit of the naïve pluripotency state.
6.3. Functions of miRNAs in the Primed Pluripotency State
There are two aspects to consider with regard to roles of miRNAs in pluripotent stem cells in the primed state: the specification of the primed state, and the differentiation from the primed state. For the former, it has been demonstrated that mouse EpiSCs can be converted from naïve ESCs and this in vitro model recapitulates the naïve-to-primed transition in utero [193] (Figure 1). A modified condition is also able to convert naïve ESCs to epiblast-like cells (EpiLCs), which are in an intermediate state between the naïve (ESC) and the primed (EpiSC) cell potential [107,194,195] (Figure 1). dgcr8−/− ESCs were significantly less efficient to be converted to EpiLCs, suggesting miRNAs are important for priming naïve ESCs [175]. Surprisingly, double knockout (dKO) of two miRNAs expressed abundantly in ESCs and EpiSCs, miR-290~295 and miR-302~367, respectively, does not influence the efficiency of EpiLC colony formation, but only influences silencing of naïve pluripotency markers and induction of some early post-implantation markers [175] (Table 2). Those different phenotypes between dgcr8−/− and dKO ESCs during the naïve-to-primed transition suggest other miRNAs also participate in this transition. Recently, a study by Du and colleagues demonstrated a protein involved in the biogenesis of polycistronic miRNAs (such as miR-17~92), Isy1, is necessary for the naïve-to-primed transition [196]. A set of miRNAs, including miR-17~92 and miR-290, were up-regulated during the naïve-to-primed transition under the positive regulation of Isy1 [196]. Importantly, overexpression of miR-20 (one member of the miR-17~92 family) is sufficient to rescue the inability of dgcr8−/− ESCs to be converted to the primed state [196]. Taken together, these results suggest that the naïve-to-primed transition could be controlled by the cooperation of multiple families of miRNAs. it is noteworthy to mention that EpiLCs may not represents all aspects of EpiSCs. For example, a strong cell death is observed in EpiLCs 48 h after Fgf2/activin induction while EpiSCs can be maintained well with same factors [107]. Thus, EpiSCs converted from ESCs, as well as dicer−/− or dgcr8−/− EpiSCs either derived from the post-implantation embryo or generated through genome editing, will elucidate the function of miRNAs in the establish, maintenance, and differentiation of/from the primed state.
As mentioned previously, let-7 plays an important role in the differentiation of mESCs from the naïve state [180]. Again, this effect could take place during the naïve-to-primed transition or during the differentiation from the primed state. A recent study addressed this question by demonstrating that knockdown of lin28 in mESCs delayed the naïve-to-primed transition while overexpression of lin28 exhibited the opposite effect [197]. Following studies suggest that Lin28 regulates the naïve-to-primed transition by targeting the let-7-dnmt3a/b-dppa3 axis [197]. Interestingly, only Lin28a, but not Lin28b, is up-regulated during the naïve-to-primed transition, suggesting those two isoforms play different roles in the exit of the naïve state [197].
Compared to mice, the expression and the function of miRNAs in human embryos are still unclear. Recent studies using cell culture models showed both similarity and disparity in mice and humans. In mESCs, although both miR-290~295 and miR-302~367 clusters are enriched, the former is dominantly expressed in the naïve state [175,198]. In contrast, miR-302~367 are dominantly expressed in EpiLCs and EpiSCs, making itself a characteristic maker for the primed pluripotent state [199]. Consistently, miR-371~373 (the analog of mouse miR-290~295) and miR-302~367 are dominantly expressed in the naïve and primed hESCs, respectively [174,197] (Figure 3). Ectopic expression of miR-290~295 miRNAs in mice or miR-302~367 miRNAs in humans enhances the acquisition of induced pluripotency during somatic reprogramming toward induced pluripotent stem cells [198,200]. Thus, in addition to the expression profile and the shared seed sequence (hence, similar or identical targets), functions of those miRNAs could also be conserved.
Despite comparable expression profiles of miRNAs in the naïve and the primed state, a loss-of-function study suggests that miRNAs could function differently in mouse and human pluripotent stem cells. Teijeiro and colleagues found that directly knockout DICER1 in human hESCs is not possible [173]. Instead, DICER1 is absolutely essential for the self-renewal of primed hESCs and can only be depleted transiently [173]. This result in human ESCs is different from mouse ones since dicer−/− ESCs can still propagate despite of G1 accumulation [135]. The authors then found that DICER1-deficient hESCs exhibit no obvious mitotic defects but are prone to apoptosis [173]. Also, knockdown of DICER or DROSHA in hESCs only led to the slow growth, with the normal expression of pluripotent genes [201]. In aggregate, those results suggest that a basal level of miRNA or endo-siRNA expression is necessary for the survival of hESCs, but not mESCs. Overexpression of either miR-302~367, miR-371~173, or miR-17~92 clusters resumed the proliferation of DICER1-deficient hESCs, possibly through blocking cell death by targeting the death-receptor FAS [173]. Although this transient-depletion system is ideal to address some unanswered questions, such as the role of miRNAs in the naïve state, in the naïve-to-primed transition, and in the differentiation from the primed state, a recent study showed that the biogenesis of some miRNAs is independent on DICER, at least in certain human cells [202]. Thus, different models, including DROSHA knockout and specific knockouts of miRNAs are still necessary to address functions of miRNAs in hESCs during the naïve-to-primed-to-differentiation process.
6.4. Functions of miRNAs in the Cell Potential of the Trophectoderm Lineage
As mentioned above, dicer−/− mice exhibit strong defects in trophoblast lineages during the development of post-implantation embryos [128]. Consistently, depletion of dicer in mouse TSCs led to growth arrest and differentiation toward trophoblast giant cells, suggesting the essential role of miRNAs in the self-renewal of mTSCs [128]. Following studies demonstrate that the miR-290~295 cluster, in addition to its role in mESCs, is also important for maintaining the proliferation of mTSCs by targeting cell cycle inhibitors such as p21, p57, rbl2, and lats2 [128] (Table 2). Up-regulation of p57 in dicer−/− mTSCs could also promote the endoreduplication and the formation of trophoblast giant cells [128]. It is still unclear whether other miRNAs also play roles in mTSCs.
Since human TSCs were only successfully derived recently, the roles of miRNAs in the self-renewal or differentiation of TSCs remain to be investigated. Notably, one primate-specific miRNA cluster, the C19MC (chromosome 19 microRNA cluster), is expressed in the TE lineage [203]. The C19MC is the largest miRNA cluster in humans, spanning ~100 kb in chromosome 19q13.41 with ~46 miRNAs paternally expressed, especially in the placenta [204]. In the placenta, C19MC miRNAs control the migration and invasion of human trophoblasts in vitro and may elicit an antiviral response in distal tissues during pregnancy while transported by exosomes [205,206]. Some C19MC cluster miRNAs, such as miR-127 and miR-372, are also highly expressed in human TSCs, making it intriguing to know the roles they play in those earliest stem cells of the TE lineage [59]. Besides of the C19MC, an eutherian-specific miRNA cluster, C14MC (within the well-known dlk1-dio3 locus), is also highly expressed in the human placenta in a maternally imprinted manner [207,208]. Interestingly, C14MC miRNAs do not seem to be expressed in TSCs based on the miRNA-seq result [59]. Knocking out miRNA biogenesis proteins or the C19MC locus in hTSCs will address functions of miRNAs in this newly derived stem cell line.
6.5. Functions of miRNAs in the Cell Potency of the Primitive Endoderm Lineage
In vitro, mouse XEN cells recapitulate the differentiation potential of the PrE. Depletion of dicer in mouse XEN cells blocked the proliferation and led to up-regulation of downstream lineage markers, including hex, apoe, and amot for the visceral endoderm, as well as gata4, ttr, alk2, and bpmp2 for the parietal endoderm [128]. Ablation of dicer led to the decreased Erk1/2 signaling, suggesting that miRNAs regulates the self-renewal and differentiation through modulating the MAPK pathway in XEN cells [128]. Yet, it is still unclear which miRNA(s) is responsible for this phenotype. In the embryo, the PrE is specified from the ICM and this differentiation process in utero could be recapitulated in vitro by the treatment of retinoic acid/activin or ectopic expression of gata6 or sox17 [98,209,210,211,212]. In the future, these two systems can be employed to model the ICM-to-PE transition.
6.6. Functions of miRNAs in the Cell Potency of the Primordial Germ Cell Lineage
In mice, dicer knockouts are embryonic lethal around the timing of PGC specification [129]. Conditional depletion of dicer in epiblasts extended the survival of embryos until E9.5, making it an ideal model for studying the function of miRNAs in the PGC specification [128]. Considering that it is not feasible to study human PGCs in vivo, in vitro models are better choices to study this process. West and colleagues established an in vitro differentiation system to harvest Stella-positive cells in embryoid bodies (EBs) to study the specification of mouse PGCs [184]. In this system, knocking down lin28 abolished the formation of PGC-like cells while ectopic expression of lin28 exhibited an opposite effect, indicating let-7 could involve in PGC specification [184]. Indeed, further studies demonstrated that let-7 blocks the transition from ESCs to PGCs by targeting blimp1, a master regulator for the PGC specification, and this suppressive effect can be relieved by up-regulation of lin28 [184]. Importantly, these results in vitro can be recapitulated in mouse embryos [184], indicating that this in vitro model is a useful tool for investigating PGC specification. In addition to this EB-differentiation protocol, a procedure for differentiating PGCLCs from naïve ESCs has been established in both mice and humans [112,113] (Figure 2). Importantly, mouse PGCLCs are able to form functional gametes that can produce offspring [107,111]. A recent study employed this cell culture model and demonstrated the role of miR-372 and let-7 in human PGC speciation, showing that miR-372 promotes while let-7 inhibits the PGC specification [185]. Obviously, more studies employing this system will increase our understanding for roles of miRNAs in critical events of the early PGC specification.
7. Discussion
There are two issues that should be considered when exploring functions of miRNAs in the early embryonic development. First, it is necessary to distinguish miRNAs and the miRNA biogenesis machinery that is maternally inherited from that which is de novo synthesized after fertilization. Since dicer knockout leads to the meiotic arrest of oocytes, a phenotype that is more attributed to loss of endo-siRNAs [150], the dgcr8 knockout mouse model serves as an alternative model to study miRNA functions since dgcr8−/− oocytes are functionally normal [133]. Yet, the other complexity lies in the biogenesis process of miRNAs. Even though the generation of most miRNAs are dependent on the microprocessor (Drosha/Dgcr8) and/or Dicer, some non-canonical miRNAs indeed exist. For example, mirtrons are generated in the dgcr8-dependent, dicer-dependent manner [22] (Figure 1). Moreover, a recent study in humans suggests that 5p miRNAs can be generated without Dicer [204]. Actually, there are different impacts on the miRNA biogenesis upon the knockout of DROSHA, DICER, or XPO5 in human cells [202]. Thus, careful analysis of expression profiles of miRNAs in different knockout models will be necessary to address miRNA functions in the early embryonic development.
The second difficulty in investigating roles of miRNAs in early embryogenesis is the functional redundancy of miRNAs. In mammals, a miRNA family containing similar or identical seed regions could consist of several individual miRNAs located at different chromosome loci and can compensate for the loss of other family members. This functional redundancy of miRNAs is best exemplified by miR-34/449 family which consists of six homologous miRNAs located at three genomic loci [158]. Mice with two-and-half loci (two alleles for each locus) depleted only exhibit a weak phenotype, whereas complete knockout of three loci led to postnatal mortality due to defects in ciliogenesis [158]. With the advent of CRISPR/Cas9-based genome editing technologies, knocking out all homologous miRNAs could reveal unexpected roles of miRNAs in the embryonic development or the stem cell potential.
In vitro cultured stem cells are valuable models for studying their cell potential in vivo. Cell culture models are particularly important for studies in humans, especially for the stem cells appeared at the later stage, like PGCs. In vitro cultured human embryos can be extended to 12–13 days [213,214], shortly after the first detection of PGCs in human embryos at day 11 [215]. Due to the ethical 14-day limit for culturing human embryos, only the very early speciation of PGCs can be studied with this system. The in vitro differentiation system of ESCs toward germ cells through PGCLC intermediates therefore provides a unique advantage to overcome the availability of human embryos and the scarce of PGCs. Same advantages can be applied to other in vitro cultured stem cells from human early embryos. However, three potential caveats should be kept in mind while using them to investigate roles of miRNAs. First, the genomic instability of ESCs cultured in the naïve state due to MEK inhibition [89], which might be resolved by a modified culturing condition, although more extensive characterization is still needed [88]. Second, the culture conditions for human TSCs have only recently been discovered, and the potency and property of hTSCs remain to be elucidated [59]. Third, the culture conditions for human XEN stem cells is not identified yet. Considering the fundamental difference in expression profiles between mouse and human blastocysts [63], it is reasonable to expect some disparities between mouse and human XEN cells in both regulatory circuitries and cell potencies.
With more understanding of miRNAs in stem cells, it is possible to alter their cell fate potential by manipulation at the level of specific miRNAs. For example, miR-290~295 and miR-302~367, both of which are enriched in ESCs, are able to promote the somatic reprogramming, i.e., the generation of iPSCs [8,200]. Also, in mice, deficiency of miR-34 led to the expanded cell fate potential toward both embryonic and extraembryonic lineages [168]. Hence, it is intriguing to see if inhibition or overexpression of certain miRNAs can drive stem cells cross barriers of the EPI, PE, and TE lineages in mice and in humans, an area not fully explored yet. Moreover, since sperm miR-34c correlates with the successful rate of ICSM, manipulating the activity of miR-34c in human zygotes could also be beneficial for in vitro fertilization [119]. Thus, with more extensive investigations, miRNAs could contribute to clinical applications in the future.
In summary, although there has been substantial evidence for the involvement of miRNAs in the development and differentiation, we just start to explore functions of miRNAs in stem cells of early embryos, especially in humans. With more advances in single-cell omics, culture conditions, and genome engineering technologies, tracing and studying roles of miRNAs in the formation and development of those “ancestor” cells will provide us new insights into secrets of the early stage of life.
Author Contributions
Conceptualization, C.-P.L., Z.Z. and L.Z.; Writing—Original Draft Preparation Z.Z., L.Z. and C.-P.L.; Writing—Review and Editing, C.-P.L., Z.Z. and L.Z.
Funding
This research was funded by National Natural Science Foundation of China #31871487.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rossant, J.; Tam, P.P.L. Exploring early human embryo development. Science 2018, 360, 1075–1076. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, K.; Hamada, H. Cell fate decisions and axis determination in the early mouse embryo. Development 2012, 139, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallaire, A.; Simard, M.J. The implication of microRNAs and endo-siRNAs in animal germline and early development. Dev. Biol. 2016, 416, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Kung, J.T.; Colognori, D.; Lee, J.T. Long noncoding RNAs: Past, present, and future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Pauli, A.; Rinn, J.L.; Schier, A.F. Non-coding RNAs as regulators of embryogenesis. Nat. Rev. Genet. 2011, 12, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, J.A.; Ventura, A. The biological functions of miRNAs: Lessons from in vivo studies. Trends Cell Biol. 2015, 25, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Li, M.A.; He, L. MicroRNAs as novel regulators of stem cell pluripotency and somatic cell reprogramming. Bioessays 2012, 34, 670–680. [Google Scholar] [CrossRef]
- Alvarez-Garcia, I.; Miska, E.A. MicroRNA functions in animal development and human disease. Development 2005, 132, 4653–4662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Alarcon, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nature 2015, 519, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, Y.; Yeom, K.H.; Kim, Y.K.; Jin, H.; Kim, V.N. The Drosha-DGCR8 complex in primary microRNA processing. Genes Dev. 2004, 18, 3016–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnsack, M.T.; Czaplinski, K.; Gorlich, D. Exportin 5 is a RanGTP-dependent dsRNA-binding protein that mediates nuclear export of pre-miRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, E.; Dahlberg, J.E. Substrate selectivity of exportin 5 and Dicer in the biogenesis of microRNAs. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.S.; Zamore, P.D. Why do miRNAs live in the miRNP? Genes Dev. 2002, 16, 1025–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.P.; Shih, I.H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of mammalian microRNA targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Lai, E.C. Micro RNAs are complementary to 3 UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef]
- van den Berg, A.; Mols, J.; Han, J.H. RISC-target interaction: Cleavage and translational suppression. Acta BBA Gene Regul. Mech. 2008, 1779, 668–677. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, T.W. Endo-siRNAs: Yet another layer of complexity in RNA silencing. Nat. Struct. Mol. Biol. 2008, 15, 546–548. [Google Scholar] [CrossRef]
- Babiarz, J.E.; Ruby, J.G.; Wang, Y.; Bartel, D.P.; Blelloch, R. Mouse ES cells express endogenous shRNAs, siRNAs, and other Microprocessor-independent, Dicer-dependent small RNAs. Genes Dev. 2008, 22, 2773–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvey, A.; Larsson, E.; Sander, C.; Leslie, C.S.; Marks, D.S. Target mRNA abundance dilutes microRNA and siRNA activity. Mol. Syst. Biol. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.M.; Baek, D.; Shin, C.; Bell, G.W.; Grimson, A.; Bartel, D.P. Weak seed-pairing stability and high target-site abundance decrease the proficiency of lsy-6 and other microRNAs. Nat. Struct. Mol. Biol. 2011, 18, 1139. [Google Scholar] [CrossRef] [PubMed]
- Chiquoine, A.D. The identification, origin, and migration of the primordial germ cells in the mouse embryo. Anat. Rec. 1954, 118, 135–146. [Google Scholar] [CrossRef]
- Ginsburg, M.; Snow, M.H.; McLaren, A. Primordial germ cells in the mouse embryo during gastrulation. Development 1990, 110, 521–528. [Google Scholar] [PubMed]
- Tam, P.P.; Snow, M.H. Proliferation and migration of primordial germ cells during compensatory growth in mouse embryos. J. Embryol. Exp. Morphol. 1981, 64, 133–147. [Google Scholar]
- Molyneaux, K.A.; Stallock, J.; Schaible, K.; Wylie, C. Time-lapse analysis of living mouse germ cell migration. Dev. Biol. 2001, 240, 488–498. [Google Scholar] [CrossRef]
- Seki, Y.; Yamaji, M.; Yabuta, Y.; Sano, M.; Shigeta, M.; Matsui, Y.; Saga, Y.; Tachibana, M.; Shinkai, Y.; Saitou, M. Cellular dynamics associated with the genome-wide epigenetic reprogramming in migrating primordial germ cells in mice. Development 2007, 134, 2627–2638. [Google Scholar] [CrossRef] [Green Version]
- Richardson, B.E.; Lehmann, R. Mechanisms guiding primordial germ cell migration: Strategies from different organisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 37–49. [Google Scholar] [CrossRef]
- O’Donnell, L. Mechanisms of spermiogenesis and spermiation and how they are disturbed. Spermatogenesis 2014, 4, e979623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszanska, B.; Borgul, A. Quantitation of nanogram amounts of nucleic acids in the presence of proteins by the ethidium bromide staining technique. Acta Biochim. Pol. 1990, 37, 59–63. [Google Scholar] [PubMed]
- Godia, M.; Swanson, G.; Krawetz, S.A. A history of why fathers’ RNA matters. Biol. Reprod. 2018, 99, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Oron, E.; Ivanova, N. Cell fate regulation in early mammalian development. Phys. Biol. 2012, 9, 045002. [Google Scholar] [CrossRef] [PubMed]
- Walser, C.B.; Lipshitz, H.D. Transcript clearance during the maternal-to-zygotic transition. Curr. Opin. Genet. Dev. 2011, 21, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Tadros, W.; Lipshitz, H.D. The maternal-to-zygotic transition: A play in two acts. Development 2009, 136, 3033–3042. [Google Scholar] [CrossRef] [PubMed]
- Bouniol, C.; Nguyen, E.; Debey, P. Endogenous transcription occurs at the 1-cell stage in the mouse embryo. Exp. Cell Res. 1995, 218, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Y.; Lin, J.M.; Liu, M.; Li, R.H.; Tian, B.; Zhang, X.; Xu, B.Y.; Liu, M.F.; Zhang, X.; Li, Y.P.; et al. Highly sensitive sequencing reveals dynamic modifications and activities of small RNAs in mouse oocytes and early embryos. Sci. Adv. 2016, 2, e1501482. [Google Scholar] [CrossRef]
- Schellander, K.; Hoelker, M.; Tesfaye, D. Selective degradation of transcripts in mammalian oocytes and embryos. Theriogenology 2007, 68, S107–S115. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, X.; Shi, J.; Yu, X.; Zhang, X.; Zhu, K.; Yi, Z.; Duan, E.; Li, L. BTG4 is a key regulator for maternal mRNA clearance during mouse early embryogenesis. J. Mol. Cell Biol. 2016, 8, 366–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Ji, S.Y.; Sha, Q.Q.; Dang, Y.; Zhou, J.J.; Zhang, Y.L.; Liu, Y.; Wang, Z.W.; Hu, B.; Sun, Q.Y.; et al. BTG4 is a meiotic cell cycle-coupled maternal-zygotic-transition licensing factor in oocytes. Nat. Struct. Mol. Biol. 2016, 23, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H.; McConnell, J.M. Lineage allocation and cell polarity during mouse embryogenesis. Semin. Cell Dev. Biol. 2004, 15, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, Y.; Ralston, A.; Stephenson, R.O.; Rossant, J. Cell and molecular regulation of the mouse blastocyst. Dev. Dyn. 2006, 235, 2301–2314. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.R.; Mermel, C.H.; Lu, J.; Lu, C.W.; Golub, T.R.; Daley, G.Q. microRNA Expression during Trophectoderm Specification. PLoS ONE 2009, 4, e6143. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z. MicroRNAs Modulate Hematopoietic Lineage Differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.M.; Ray, P.F.; Ao, A.; Winston, R.M.; Handyside, A.H. Paternal transcripts for glucose-6-phosphate dehydrogenase and adenosine deaminase are first detectable in the human preimplantation embryo at the three- to four-cell stage. Mol. Reprod. Dev. 1997, 48, 442–448. [Google Scholar] [CrossRef]
- Vassena, R.; Boue, S.; Gonzalez-Roca, E.; Aran, B.; Auer, H.; Veiga, A.; Izpisua Belmonte, J.C. Waves of early transcriptional activation and pluripotency program initiation during human preimplantation development. Development 2011, 138, 3699–3709. [Google Scholar] [CrossRef] [Green Version]
- Niakan, K.K.; Han, J.; Pedersen, R.A.; Simon, C.; Pera, R.A.R. Human pre-implantation embryo development. Development 2012, 139, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Hudson, Q.J.; Kulinski, T.M.; Huetter, S.P.; Barlow, D.P. Genomic imprinting mechanisms in embryonic and extraembryonic mouse tissues. Heredity 2010, 105, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Tam, P.P.; Rossant, J. Mouse embryonic chimeras: Tools for studying mammalian development. Development 2003, 130, 6155–6163. [Google Scholar] [CrossRef] [PubMed]
- Gardner, R.L.; Rossant, J. Investigation of the fate of 4-5 day post-coitum mouse inner cell mass cells by blastocyst injection. J. Embryol. Exp. Morphol. 1979, 52, 141–152. [Google Scholar] [PubMed]
- Kwon, G.S.; Viotti, M.; Hadjantonakis, A.K. The endoderm of the mouse embryo arises by dynamic widespread intercalation of embryonic and extraembryonic lineages. Dev. Cell 2008, 15, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Rossant, J. Cell lineage analysis in mammalian embryogenesis. Curr. Top. Dev. Biol. 1987, 23, 115–146. [Google Scholar]
- Rossant, J.; Tam, P.P. Blastocyst lineage formation, early embryonic asymmetries and axis patterning in the mouse. Development 2009, 136, 701–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Niakan, K.K.; Schrode, N.; Cho, L.T.; Hadjantonakis, A.K. Derivation of extraembryonic endoderm stem (XEN) cells from mouse embryos and embryonic stem cells. Nat. Protoc. 2013, 8, 1028–1041. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane, K.; Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell 2018, 22, 50–63. [Google Scholar] [CrossRef]
- Tanaka, S.; Kunath, T.; Hadjantonakis, A.K.; Nagy, A.; Rossant, J. Promotion of trophoblast stem cell proliferation by FGF4. Science 1998, 282, 2072–2075. [Google Scholar] [CrossRef]
- Shamblott, M.J.; Axelman, J.; Wang, S.; Bugg, E.M.; Littlefield, J.W.; Donovan, P.J.; Blumenthal, P.D.; Huggins, G.R.; Gearhart, J.D. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc. Natl. Acad. Sci. USA 1998, 95, 13726–13731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnpenny, L.; Brickwood, S.; Spalluto, C.M.; Piper, K.; Cameron, I.T.; Wilson, D.I.; Hanley, N.A. Derivation of human embryonic germ cells: An alternative source of pluripotent stem cells. Stem Cells 2003, 21, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Blakeley, P.; Fogarty, N.M.; del Valle, I.; Wamaitha, S.E.; Hu, T.X.; Elder, K.; Snell, P.; Christie, L.; Robson, P.; Niakan, K.K. Defining the three cell lineages of the human blastocyst by single-cell RNA-seq. Development 2015, 142, 3151–3165. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.G.; Heath, J.K.; Donaldson, D.D.; Wong, G.G.; Moreau, J.; Stahl, M.; Rogers, D. Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature 1988, 336, 688–690. [Google Scholar] [CrossRef]
- Eakin, G.S.; Hadjantonakis, A.K. Production of chimeras by aggregation of embryonic stem cells with diploid or tetraploid mouse embryos. Nat. Protoc. 2006, 1, 1145–1153. [Google Scholar] [CrossRef]
- Wen, D.C.; Saiz, N.; Rosenwaks, Z.; Hadjantonakis, A.K.; Rafii, S. Completely ES Cell-Derived Mice Produced by Tetraploid Complementation Using Inner Cell Mass (ICM) Deficient Blastocysts. PLoS ONE 2014, 9, e94730. [Google Scholar] [CrossRef]
- Xu, R.H.; Peck, R.M.; Li, D.S.; Feng, X.; Ludwig, T.; Thomson, J.A. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nat. Methods 2005, 2, 185–190. [Google Scholar] [CrossRef]
- Wu, J.; Belmonte, J.C.I. Dynamic Pluripotent Stem Cell States and Their Applications. Cell Stem Cell 2015, 17, 509–525. [Google Scholar] [CrossRef] [Green Version]
- Vallier, L.; Alexander, M.; Pedersen, R.A. Activin/Nodal and FGF pathways cooperate to maintain pluripotency of human embryonic stem cells. J. Cell Sci. 2005, 118, 4495–4509. [Google Scholar] [CrossRef] [Green Version]
- Brons, I.G.; Smithers, L.E.; Trotter, M.W.; Rugg-Gunn, P.; Sun, B.; Chuva de Sousa Lopes, S.M.; Howlett, S.K.; Clarkson, A.; Ahrlund-Richter, L.; Pedersen, R.A.; et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature 2007, 448, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Tesar, P.J.; Chenoweth, J.G.; Brook, F.A.; Davies, T.J.; Evans, E.P.; Mack, D.L.; Gardner, R.L.; McKay, R.D. New cell lines from mouse epiblast share defining features with human embryonic stem cells. Nature 2007, 448, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.L.; Nichols, J.; Chambers, I.; Smith, A. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell 2003, 115, 281–292. [Google Scholar] [CrossRef]
- Niwa, H.; Burdon, T.; Chambers, I.; Smith, A. Self-renewal of pluripotent embryonic stem cells is mediated via activation of STAT3. Genes Dev. 1998, 12, 2048–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdon, T.; Stracey, C.; Chambers, I.; Nichols, J.; Smith, A. Suppression of SHP-2 and ERK signalling promotes self-renewal of mouse embryonic stem cells. Dev. Biol. 1999, 210, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Wray, J.; Kalkan, T.; Smith, A.G. The ground state of pluripotency. Biochem. Soc. Trans. 2010, 38, 1027–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, N.; Meijer, L.; Skaltsounis, L.; Greengard, P.; Brivanlou, A.H. Maintenance of pluripotency in human and mouse embryonic stem cells through activation of Wnt signaling by a pharmacological GSK-3-specific inhibitor. Nat. Med. 2004, 10, 55–63. [Google Scholar] [CrossRef]
- Ying, Q.L.; Wray, J.; Nichols, J.; Batlle-Morera, L.; Doble, B.; Woodgett, J.; Cohen, P.; Smith, A. The ground state of embryonic stem cell self-renewal. Nature 2008, 453, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Smith, A. Formative pluripotency: The executive phase in a developmental continuum. Development 2017, 144, 365–373. [Google Scholar] [CrossRef]
- Takashima, Y.; Guo, G.; Loos, R.; Nichols, J.; Ficz, G.; Krueger, F.; Oxley, D.; Santos, F.; Clarke, J.; Mansfield, W.; et al. Resetting transcription factor control circuitry toward ground-state pluripotency in human. Cell 2014, 158, 1254–1269. [Google Scholar] [CrossRef]
- Theunissen, T.W.; Powell, B.E.; Wang, H.; Mitalipova, M.; Faddah, D.A.; Reddy, J.; Fan, Z.P.; Maetzel, D.; Ganz, K.; Shi, L.; et al. Systematic identification of culture conditions for induction and maintenance of naive human pluripotency. Cell Stem Cell 2014, 15, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Cheng, A.W.; Saha, K.; Kim, J.; Lengner, C.J.; Soldner, F.; Cassady, J.P.; Muffat, J.; Carey, B.W.; Jaenisch, R. Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs. Proc. Natl. Acad. Sci. USA 2010, 107, 9222–9227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gafni, O.; Weinberger, L.; Mansour, A.A.; Manor, Y.S.; Chomsky, E.; Ben-Yosef, D.; Kalma, Y.; Viukov, S.; Maza, I.; Zviran, A.; et al. Derivation of novel human ground state naive pluripotent stem cells. Nature 2013, 504, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Goke, J.; Ng, J.H.; Lu, X.; Gonzales, K.A.; Tan, C.P.; Tng, W.Q.; Hong, Z.Z.; Lim, Y.S.; Ng, H.H. Induction of a human pluripotent state with distinct regulatory circuitry that resembles preimplantation epiblast. Cell Stem Cell 2013, 13, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, G.; Singh, M.; Ghanbarian, A.T.; Rasko, T.; Szvetnik, A.; Cai, H.; Besser, D.; Prigione, A.; Fuchs, N.V.; et al. Primate-specific endogenous retrovirus-driven transcription defines naive-like stem cells. Nature 2014, 516, 405–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggal, G.; Warrier, S.; Ghimire, S.; Broekaert, D.; Van der Jeught, M.; Lierman, S.; Deroo, T.; Peelman, L.; Van Soom, A.; Cornelissen, R.; et al. Alternative Routes to Induce Naive Pluripotency in Human Embryonic Stem Cells. Stem Cells 2015, 33, 2686–2698. [Google Scholar] [CrossRef]
- Ware, C.B.; Nelson, A.M.; Mecham, B.; Hesson, J.; Zhou, W.; Jonlin, E.C.; Jimenez-Caliani, A.J.; Deng, X.; Cavanaugh, C.; Cook, S.; et al. Derivation of naive human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 4484–4489. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, B.; Ueda, M.; Sabri, S.; Brumbaugh, J.; Huebner, A.J.; Sahakyan, A.; Clement, K.; Clowers, K.J.; Erickson, A.R.; Shioda, K.; et al. Reduced MEK inhibition preserves genomic stability in naive human embryonic stem cells. Nat. Methods 2018, 15, 732–740. [Google Scholar] [CrossRef]
- Choi, J.; Huebner, A.J.; Clement, K.; Walsh, R.M.; Savol, A.; Lin, K.; Gu, H.; Di Stefano, B.; Brumbaugh, J.; Kim, S.Y.; et al. Prolonged Mek1/2 suppression impairs the developmental potential of embryonic stem cells. Nature 2017, 548, 219–223. [Google Scholar] [CrossRef]
- Yagi, M.; Kishigami, S.; Tanaka, A.; Semi, K.; Mizutani, E.; Wakayama, S.; Wakayama, T.; Yamamoto, T.; Yamada, Y. Derivation of ground-state female ES cells maintaining gamete-derived DNA methylation. Nature 2017, 548, 224–227. [Google Scholar] [CrossRef]
- Kunath, T.; Arnaud, D.; Uy, G.D.; Okamoto, I.; Chureau, C.; Yamanaka, Y.; Heard, E.; Gardner, R.L.; Avner, P.; Rossant, J. Imprinted X-inactivation in extra-embryonic endoderm cell lines from mouse blastocysts. Development 2005, 132, 1649–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Khan, M.; Zapiec, B.; Mombaerts, P. Efficient derivation of extraembryonic endoderm stem cell lines from mouse postimplantation embryos. Sci. Rep. 2016, 6, 39457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruithof-de Julio, M.; Alvarez, M.J.; Galli, A.; Chu, J.; Price, S.M.; Califano, A.; Shen, M.M. Regulation of extra-embryonic endoderm stem cell differentiation by Nodal and Cripto signaling. Development 2011, 138, 3885–3895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paca, A.; Seguin, C.A.; Clements, M.; Ryczko, M.; Rossant, J.; Rodriguez, T.A.; Kunath, T. BMP signaling induces visceral endoderm differentiation of XEN cells and parietal endoderm. Dev. Biol. 2012, 361, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artus, J.; Douvaras, P.; Piliszek, A.; Isern, J.; Baron, M.H.; Hadjantonakis, A.K. BMP4 signaling directs primitive endoderm-derived XEN cells to an extraembryonic visceral endoderm identity. Dev. Biol. 2012, 361, 245–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niakan, K.K.; Eggan, K. Analysis of human embryos from zygote to blastocyst reveals distinct gene expression patterns relative to the mouse. Dev. Biol. 2013, 375, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermitte, S.; Chazaud, C. Primitive endoderm differentiation: From specification to epithelium formation. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef]
- Cho, L.T.; Wamaitha, S.E.; Tsai, I.J.; Artus, J.; Sherwood, R.I.; Pedersen, R.A.; Hadjantonakis, A.K.; Niakan, K.K. Conversion from mouse embryonic to extra-embryonic endoderm stem cells reveals distinct differentiation capacities of pluripotent stem cell states. Development 2012, 139, 2866–2877. [Google Scholar] [CrossRef] [Green Version]
- Wamaitha, S.E.; del Valle, I.; Cho, L.T.; Wei, Y.; Fogarty, N.M.; Blakeley, P.; Sherwood, R.I.; Ji, H.; Niakan, K.K. Gata6 potently initiates reprograming of pluripotent and differentiated cells to extraembryonic endoderm stem cells. Genes Dev. 2015, 29, 1239–1255. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garcia, V.; Fineberg, E.; Wilson, R.; Murray, A.; Mazzeo, C.I.; Tudor, C.; Sienerth, A.; White, J.K.; Tuck, E.; Ryder, E.J.; et al. Placentation defects are highly prevalent in embryonic lethal mouse mutants. Nature 2018, 555, 463–468. [Google Scholar] [CrossRef]
- Kunath, T.; Yamanaka, Y.; Detmar, J.; MacPhee, D.; Caniggia, I.; Rossant, J.; Jurisicova, A. Developmental differences in the expression of FGF receptors between human and mouse embryos. Placenta 2014, 35, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Soncin, F.; Natale, D.; Parast, M.M. Signaling pathways in mouse and human trophoblast differentiation: A comparative review. Cell Mol. Life Sci. 2015, 72, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Matsui, Y.; Zsebo, K.; Hogan, B.L. Derivation of pluripotential embryonic stem cells from murine primordial germ cells in culture. Cell 1992, 70, 841–847. [Google Scholar] [CrossRef]
- Pesce, M.; Canipari, R.; Ferri, G.L.; Siracusa, G.; De Felici, M. Pituitary adenylate cyclase-activating polypeptide (PACAP) stimulates adenylate cyclase and promotes proliferation of mouse primordial germ cells. Development 1996, 122, 215–221. [Google Scholar] [PubMed]
- Resnick, J.L.; Bixler, L.S.; Cheng, L.; Donovan, P.J. Long-term proliferation of mouse primordial germ cells in culture. Nature 1992, 359, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Gell, J.J.; Tao, Y.; Sosa, E.; Clark, A.T. Modeling human infertility with pluripotent stem cells. Stem Cell Res. 2017, 21, 187–192. [Google Scholar] [CrossRef]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 2011, 146, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Mitsunaga, S.; Odajima, J.; Yawata, S.; Shioda, K.; Owa, C.; Isselbacher, K.J.; Hanna, J.H.; Shioda, T. Relevance of iPSC-derived human PGC-like cells at the surface of embryoid bodies to prechemotaxis migrating PGCs. Proc. Natl. Acad. Sci. USA 2017, 114, E9913–E9922. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Saburi, S.; Yamanouchi, K.; Tojo, H.; Tachi, C. In vitro studies on PGC or PGC-like cells in cultured yolk sac cells and embryonic stem cells of the mouse. Arch. Histol. Cytol. 2000, 63, 229–241. [Google Scholar] [CrossRef]
- Chuang, C.Y.; Lin, K.I.; Hsiao, M.; Stone, L.; Chen, H.F.; Huang, Y.H.; Lin, S.P.; Ho, H.N.; Kuo, H.C. Meiotic competent human germ cell-like cells derived from human embryonic stem cells induced by BMP4/WNT3A signaling and OCT4/EpCAM (epithelial cell adhesion molecule) selection. J. Biol. Chem. 2012, 287, 14389–14401. [Google Scholar] [CrossRef]
- Hayashi, K.; Ogushi, S.; Kurimoto, K.; Shimamoto, S.; Ohta, H.; Saitou, M. Offspring from Oocytes Derived from in Vitro Primordial Germ Cell-like Cells in Mice. Science 2012, 338, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Yokobayashi, S.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Kurimoto, K.; Ohta, H.; Moritoki, Y.; Iwatani, C.; Tsuchiya, H.; et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell 2015, 17, 178–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, N.; Weinberger, L.; Tang, W.W.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Kaneda, M.; O’Carroll, D.; Hajkova, P.; Barton, S.C.; Sun, Y.A.; Lee, C.; Tarakhovsky, A.; Lao, K.; Surani, M.A. Maternal microRNAs are essential for mouse zygotic development. Genes Dev. 2007, 21, 644–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, N.; Kropp, J.; Khatib, H. MicroRNA Signaling in Embryo Development. Biology 2017, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bao, J.; Kim, M.; Yuan, S.; Tang, C.; Zheng, H.; Mastick, G.S.; Xu, C.; Yan, W. Two miRNA clusters, miR-34b/c and miR-449, are essential for normal brain development, motile ciliogenesis, and spermatogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, E2851–E2857. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.P.; Guo, W.W.; Zhang, Z.H.; Yu, W.X. A coupled convolutional neural network for small and densely clustered ship detection in SAR images. Sci. China Inform. Sci. 2019, 62. [Google Scholar] [CrossRef]
- Liu, W.M.; Pang, R.T.K.; Chiu, P.C.N.; Wong, B.P.C.; Lao, K.Q.; Lee, K.F.; Yeung, W.S.B. Sperm-borne microRNA-34c is required for the first cleavage division in mouse. Proc. Natl. Acad. Sci. USA 2012, 109, 490–494. [Google Scholar] [CrossRef]
- Cui, L.; Fang, L.; Shi, B.; Qiu, S.; Ye, Y. Spermatozoa micro ribonucleic acid–34c level is correlated with intracytoplasmic sperm injection outcomes. Fertil. Steril. 2015, 104, 312–317. [Google Scholar] [CrossRef]
- Rassoulzadegan, M.; Grandjean, V.; Gounon, P.; Vincent, S.; Gillot, I.; Cuzin, F. RNA-mediated non-mendelian inheritance of an epigenetic change in the mouse. Nature 2006, 441, 469–474. [Google Scholar] [CrossRef]
- Rodgers, A.B.; Morgan, C.P.; Leu, N.A.; Bale, T.L. Transgenerational epigenetic programming via sperm microRNA recapitulates effects of paternal stress. Proc. Natl. Acad. Sci. USA 2015, 112, 13699–13704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.H.; Bai, W.T.; Zhang, L.; Yin, G.W.; Wang, X.H.; Wang, J.; Zhao, H.X.; Han, Y.B.; Yao, Y.Q. Determination of microRNAs in mouse preimplantation embryos by microarray. Dev. Dyn. 2008, 237, 2315–2327. [Google Scholar] [CrossRef] [PubMed]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic Stem Cell-Specific MicroRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar] [CrossRef]
- Foshay, K.M.; Gallicano, G.I. miR-17 family miRNAs are expressed during early mammalian development and regulate stem cell differentiation. Dev. Biol. 2009, 326, 431–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbluth, E.M.; Shelton, D.N.; Sparks, A.E.T.; Devor, E.; Christenson, L.; Van Voorhis, B.J. MicroRNA expression in the human blastocyst. Fertil. Steril. 2013, 99, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, L.A.; Dennis, L.M.; Gill, M.E.; Houbaviy, H.; Markoulaki, S.; Fu, D.; White, A.C.; Kirak, O.; Sharp, P.A.; Page, D.C.; et al. Mir-290-295 deficiency in mice results in partially penetrant embryonic lethality and germ cell defects. Proc. Natl. Acad. Sci. USA 2011, 108, 14163–14168. [Google Scholar] [CrossRef]
- Liu, W.M.; Niu, Z.R.; Li, Q.; Pang, R.T.K.; Chiu, P.C.N.; Yeung, W.S.B. MicroRNA and Embryo Implantation. Am. J. Reprod. Immunol. 2016, 75, 263–271. [Google Scholar] [CrossRef]
- Spruce, T.; Pernaute, B.; Di-Gregorio, A.; Cobb, B.S.; Merkenschlager, M.; Manzanares, M.; Rodriguez, T.A. An Early Developmental Role for miRNAs in the Maintenance of Extraembryonic Stem Cells in the Mouse Embryo. Dev. Cell 2010, 19, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef]
- Pernaute, B.; Spruce, T.; Rodriguez, T.A.; Manzanares, M. miRNA-mediated regulation of cell signaling and homeostasis in the early mouse embryo. Cell Cycle 2011, 10, 584–591. [Google Scholar] [CrossRef]
- Murchison, E.P.; Stein, P.; Xuan, Z.; Pan, H.; Zhang, M.Q.; Schultz, R.M.; Hannon, G.J. Critical roles for Dicer in the female germline. Genes Dev. 2007, 21, 682–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Flemr, M.; Stein, P.; Berninger, P.; Malik, R.; Zavolan, M.; Svoboda, P.; Schultz, R.M. MicroRNA Activity Is Suppressed in Mouse Oocytes. Curr. Biol. 2010, 20, 265–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, N.; Baehner, L.; Moltzahn, F.; Melton, C.; Shenoy, A.; Chen, J.; Blelloch, R. MicroRNA Function Is Globally Suppressed in Mouse Oocytes and Early Embryos. Curr. Biol. 2010, 20, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murchison, E.P.; Partridge, J.F.; Tam, O.H.; Cheloufi, S.; Hannon, G.J. Characterization of Dicer-deficient murine embryonic stem cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12135–12140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Medvid, R.; Melton, C.; Jaenisch, R.; Blelloch, R. DGCR8 is essential for microRNA biogenesis and silencing of embryonic stem cell self-renewal. Nat. Genet. 2007, 39, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Baskerville, S.; Shenoy, A.; Babiarz, J.E.; Baehner, L.; Blelloch, R. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat. Genet. 2008, 40, 1478–1483. [Google Scholar] [CrossRef]
- Cheloufi, S.; Dos Santos, C.O.; Chong, M.M.; Hannon, G.J. A dicer-independent miRNA biogenesis pathway that requires Ago catalysis. Nature 2010, 465, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes, D.; Xue, H.; Taylor, D.W.; Patnode, H.; Mishima, Y.; Cheloufi, S.; Ma, E.; Mane, S.; Hannon, G.J.; Lawson, N.D.; et al. A novel miRNA processing pathway independent of Dicer requires Argonaute2 catalytic activity. Science 2010, 328, 1694–1698. [Google Scholar] [CrossRef]
- Klattenhoff, C.; Theurkauf, W. Biogenesis and germline functions of piRNAs. Development 2008, 135, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Houwing, S.; Kamminga, L.M.; Berezikov, E.; Cronembold, D.; Girard, A.; van den Elst, H.; Filippov, D.V.; Blaser, H.; Raz, E.; Moens, C.B.; et al. A role for Piwi and piRNAs in germ cell maintenance and transposon silencing in Zebrafish. Cell 2007, 129, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Vagin, V.V.; Sigova, A.; Li, C.; Seitz, H.; Gvozdev, V.; Zamore, P.D. A distinct small RNA pathway silences selfish genetic elements in the germline. Science 2006, 313, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Carmell, M.A.; Girard, A.; van de Kant, H.J.; Bourc’his, D.; Bestor, T.H.; de Rooij, D.G.; Hannon, G.J. MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline. Dev. Cell 2007, 12, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Lin, H. Miwi, a murine homolog of piwi, encodes a cytoplasmic protein essential for spermatogenesis. Dev. Cell 2002, 2, 819–830. [Google Scholar] [CrossRef]
- Kuramochi-Miyagawa, S.; Kimura, T.; Yomogida, K.; Kuroiwa, A.; Tadokoro, Y.; Fujita, Y.; Sato, M.; Matsuda, Y.; Nakano, T. Two mouse piwi-related genes: Miwi and mili. Mech. Dev. 2001, 108, 121–133. [Google Scholar] [CrossRef]
- Kuramochi-Miyagawa, S.; Watanabe, T.; Gotoh, K.; Totoki, Y.; Toyoda, A.; Ikawa, M.; Asada, N.; Kojima, K.; Yamaguchi, Y.; Ijiri, T.W.; et al. DNA methylation of retrotransposon genes is regulated by Piwi family members MILI and MIWI2 in murine fetal testes. Genes Dev. 2008, 22, 908–917. [Google Scholar] [CrossRef] [Green Version]
- Roovers, E.F.; Rosenkranz, D.; Mahdipour, M.; Han, C.T.; He, N.; Chuva de Sousa Lopes, S.M.; van der Westerlaken, L.A.; Zischler, H.; Butter, F.; Roelen, B.A.; et al. Piwi proteins and piRNAs in mammalian oocytes and early embryos. Cell Rep. 2015, 10, 2069–2082. [Google Scholar] [CrossRef]
- Watanabe, T.; Totoki, Y.; Toyoda, A.; Kaneda, M.; Kuramochi-Miyagawa, S.; Obata, Y.; Chiba, H.; Kohara, Y.; Kono, T.; Nakano, T.; et al. Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes. Nature 2008, 453, 539–543. [Google Scholar] [CrossRef]
- Tam, O.H.; Aravin, A.A.; Stein, P.; Girard, A.; Murchison, E.P.; Cheloufi, S.; Hodges, E.; Anger, M.; Sachidanandam, R.; Schultz, R.M.; et al. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 2008, 453, 534–538. [Google Scholar] [CrossRef] [Green Version]
- Flemr, M.; Malik, R.; Franke, V.; Nejepinska, J.; Sedlacek, R.; Vlahovicek, K.; Svoboda, P. A retrotransposon-driven dicer isoform directs endogenous small interfering RNA production in mouse oocytes. Cell 2013, 155, 807–816. [Google Scholar] [CrossRef]
- Razak, S.R.; Ueno, K.; Takayama, N.; Nariai, N.; Nagasaki, M.; Saito, R.; Koso, H.; Lai, C.Y.; Murakami, M.; Tsuji, K.; et al. Profiling of microRNA in human and mouse ES and iPS cells reveals overlapping but distinct microRNA expression patterns. PLoS ONE 2013, 8, e73532. [Google Scholar] [CrossRef] [PubMed]
- Paikari, A.; Belair, C.D.; Saw, D.; Blelloch, R. The eutheria-specific miR-290 cluster modulates placental growth and maternal-fetal transport. Development 2017, 144, 3731–3743. [Google Scholar] [CrossRef] [PubMed]
- Ventura, A.; Young, A.G.; Winslow, M.M.; Lintault, L.; Meissner, A.; Erkeland, S.J.; Newman, J.; Bronson, R.T.; Crowley, D.; Stone, J.R.; et al. Targeted deletion reveals essential and overlapping functions of the miR-17 through 92 family of miRNA clusters. Cell 2008, 132, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.C.; Vidigal, J.A.; Mu, P.; Yao, E.; Singh, I.; Gonzalez, A.J.; Concepcion, C.P.; Bonetti, C.; Ogrodowski, P.; Carver, B.; et al. An allelic series of miR-17 approximately 92-mutant mice uncovers functional specialization and cooperation among members of a microRNA polycistron. Nat. Genet. 2015, 47, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Z.; O’Loughlin, E.; Wang, L.; Fan, X.; Lai, E.C.; Yi, R. MicroRNA-205 controls neonatal expansion of skin stem cells by modulating the PI(3)K pathway. Nat. Cell Biol. 2013, 15, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef]
- Song, R.; Walentek, P.; Sponer, N.; Klimke, A.; Lee, J.S.; Dixon, G.; Harland, R.; Wan, Y.; Lishko, P.; Lize, M.; et al. miR-34/449 miRNAs are required for motile ciliogenesis by repressing cp110. Nature 2014, 510, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Parchem, R.J.; Moore, N.; Fish, J.L.; Parchem, J.G.; Braga, T.T.; Shenoy, A.; Oldham, M.C.; Rubenstein, J.L.; Schneider, R.A.; Blelloch, R. miR-302 Is Required for Timing of Neural Differentiation, Neural Tube Closure, and Embryonic Viability. Cell Rep. 2015, 12, 760–773. [Google Scholar] [CrossRef] [Green Version]
- Hasuwa, H.; Ueda, J.; Ikawa, M.; Okabe, M. miR-200b and miR-429 function in mouse ovulation and are essential for female fertility. Science 2013, 341, 71–73. [Google Scholar] [CrossRef]
- Tan, C.L.; Plotkin, J.L.; Veno, M.T.; von Schimmelmann, M.; Feinberg, P.; Mann, S.; Handler, A.; Kjems, J.; Surmeier, D.J.; O’Carroll, D.; et al. MicroRNA-128 governs neuronal excitability and motor behavior in mice. Science 2013, 342, 1254–1258. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Guerau-de-Arellano, M.; Costinean, S.; Williams, J.L.; Bottoni, A.; Mavrikis Cox, G.; Satoskar, A.R.; Croce, C.M.; Racke, M.K.; Lovett-Racke, A.E.; et al. miR-29ab1 deficiency identifies a negative feedback loop controlling Th1 bias that is dysregulated in multiple sclerosis. J. Immunol. 2012, 189, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Bezprozvannaya, S.; Williams, A.H.; Qi, X.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. microRNA-133a regulates cardiomyocyte proliferation and suppresses smooth muscle gene expression in the heart. Genes Dev. 2008, 22, 3242–3254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Lopez, J.; del Mazo, J. Expression dynamics of microRNA biogenesis during preimplantation mouse development. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2012, 1819, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Mineno, J.; Okamoto, S.; Ando, T.; Sato, M.; Chono, H.; Izu, H.; Takayama, M.; Asada, K.; Mirochnitchenko, O.; Inouye, M.; et al. The expression profile of microRNAs in mouse embryos. Nucleic Acids Res. 2006, 34, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Freimer, J.W.; Krishnakumar, R.; Cook, M.S.; Blelloch, R. Expression of Alternative Ago2 Isoform Associated with Loss of microRNA-Driven Translational Repression in Mouse Oocytes. Curr. Biol. 2018, 28, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Marikawa, Y.; Alarcon, V.B. Establishment of trophectoderm and inner cell mass lineages in the mouse embryo. Mol. Reprod. Dev. 2009, 76, 1019–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Lin, C.P.; Risso, D.; Chen, S.; Kim, T.A.; Tan, M.H.; Li, J.B.; Wu, Y.; Chen, C.; Xuan, Z.; et al. Deficiency of microRNA miR-34a expands cell fate potential in pluripotent stem cells. Science 2017, 355. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ryan, D.J.; Wang, W.; Tsang, J.C.; Lan, G.; Masaki, H.; Gao, X.; Antunes, L.; Yu, Y.; Zhu, Z.; et al. Establishment of mouse expanded potential stem cells. Nature 2017, 550, 393–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liu, B.; Xu, J.; Wang, J.; Wu, J.; Shi, C.; Xu, Y.; Dong, J.; Wang, C.; Lai, W.; et al. Derivation of Pluripotent Stem Cells with In Vivo Embryonic and Extraembryonic Potency. Cell 2017, 169, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Marson, A.; Levine, S.S.; Cole, M.F.; Frampton, G.M.; Brambrink, T.; Johnstone, S.; Guenther, M.G.; Johnston, W.K.; Wernig, M.; Newman, J.; et al. Connecting microRNA genes to the core transcriptional regulatory circuitry of embryonic stem cells. Cell 2008, 134, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Xiao, C.H.; Xiang, G.M.; Ran, X.Z. Regulation of Embryonic Stem Cell Self-Renewal and Differentiation by MicroRNAs. Cell. Reprogram. 2017, 19, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Teijeiro, V.; Yang, D.; Majumdar, S.; Gonzalez, F.; Rickert, R.W.; Xu, C.; Koche, R.; Verma, N.; Lai, E.C.; Huangfu, D. DICER1 Is Essential for Self-Renewal of Human Embryonic Stem Cells. Stem Cell Rep. 2018, 11, 616–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faridani, O.R.; Abdullayev, I.; Hagemann-Jensen, M.; Schell, J.P.; Lanner, F.; Sandberg, R. Single-cell sequencing of the small-RNA transcriptome. Nat. Biotechnol. 2016, 34, 1264–1266. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.L.; Zhang, Q.; Yan, Y.; Li, T.T.; Duan, F.F.; Hao, J.; Wang, X.W.; Shi, M.; Wu, D.R.; Guo, W.T.; et al. Pluripotency-associated miR-290/302 family of microRNAs promote the dismantling of naive pluripotency. Cell Res. 2016, 26, 350–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.X.; Ravi, A.; Calabrese, J.M.; Medeiros, L.A.; Kirak, O.; Dennis, L.M.; Jaenisch, R.; Burge, C.B.; Sharp, P.A. A latent pro-survival function for the mir-290-295 cluster in mouse embryonic stem cells. PLoS Genet. 2011, 7, e1002054. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Hao, J.; Guo, W.T.; Liao, L.Q.; Huang, S.Y.; Guo, X.; Bao, X.; Esteban, M.A.; Wang, Y. A DGCR8-Independent Stable MicroRNA Expression Strategy Reveals Important Functions of miR-290 and miR-183-182 Families in Mouse Embryonic Stem Cells. Stem Cell Rep. 2017, 9, 1618–1629. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.T.; Wang, X.W.; Yan, Y.L.; Li, Y.P.; Yin, X.; Zhang, Q.; Melton, C.; Shenoy, A.; Reyes, N.A.; Oakes, S.A.; et al. Suppression of epithelial-mesenchymal transition and apoptotic pathways by miR-294/302 family synergistically blocks let-7-induced silencing of self-renewal in embryonic stem cells. Cell Death Differ. 2015, 22, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Melton, C.; Li, Y.P.; Shenoy, A.; Zhang, X.X.; Subramanyam, D.; Blelloch, R. miR-294/miR-302 promotes proliferation, suppresses G1-S restriction point, and inhibits ESC differentiation through separable mechanisms. Cell Rep. 2013, 4, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Melton, C.; Judson, R.L.; Blelloch, R. Opposing microRNA families regulate self-renewal in mouse embryonic stem cells. Nature 2010, 463, 621–626. [Google Scholar] [CrossRef]
- Kim, B.M.; Choi, M.Y. Non-canonical microRNAs miR-320 and miR-702 promote proliferation in Dgcr8-deficient embryonic stem cells. Biochem. Biophys. Res. Commun. 2012, 426, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Lin, Y.; Zhao, Z.A.; Lu, X.; Yu, Y.; Zhang, X.; Wang, Q.; Li, L. MicroRNA-127 Promotes Mesendoderm Differentiation of Mouse Embryonic Stem Cells by Targeting Left-Right Determination Factor 2. J. Biol. Chem. 2016, 291, 12126–12135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernaute, B.; Spruce, T.; Smith, K.M.; Sanchez-Nieto, J.M.; Manzanares, M.; Cobb, B.; Rodriguez, T.A. MicroRNAs control the apoptotic threshold in primed pluripotent stem cells through regulation of BIM. Genes Dev. 2014, 28, 1873–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.A.; Viswanathan, S.R.; Yabuuchi, A.; Cunniff, K.; Takeuchi, A.; Park, I.H.; Sero, J.E.; Zhu, H.; Perez-Atayde, A.; Frazier, A.L.; et al. A role for Lin28 in primordial germ-cell development and germ-cell malignancy. Nature 2009, 460, 909–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, N.D.; Kissner, M.; Subramanyam, D.; Parchem, R.J.; Laird, D.J.; Blelloch, R.H. A miR-372/let-7 Axis Regulates Human Germ Versus Somatic Cell Fates. Stem Cells 2016, 34, 1985–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, I.; Joo, C.; Cho, J.; Ha, M.; Han, J.; Kim, V.N. Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA. Mol. Cell 2008, 32, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.A.; Thomson, J.M.; Hammond, S.M. Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing. RNA 2008, 14, 1539–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; LaPierre, R.J.; Pothoulakis, C.; Hagan, J.P.; Iliopoulos, D.; Gregory, R.I. Lin28A and Lin28B inhibit let-7 microRNA biogenesis by distinct mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef]
- Viswanathan, S.R.; Daley, G.Q.; Gregory, R.I. Selective blockade of microRNA processing by Lin28. Science 2008, 320, 97–100. [Google Scholar] [CrossRef]
- Yin, J.; Zhao, J.; Hu, W.; Yang, G.; Yu, H.; Wang, R.; Wang, L.; Zhang, G.; Fu, W.; Dai, L.; et al. Disturbance of the let-7/LIN28 double-negative feedback loop is associated with radio- and chemo-resistance in non-small cell lung cancer. PLoS ONE 2017, 12, e0172787. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.M.; Parker, J.; Perou, C.M.; Hammond, S.M. A custom microarray platform for analysis of microRNA gene expression. Nat. Methods 2004, 1, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, G.; Yang, J.; Nichols, J.; Hall, J.S.; Eyres, I.; Mansfield, W.; Smith, A. Klf4 reverts developmentally programmed restriction of ground state pluripotency. Development 2009, 136, 1063–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buecker, C.; Srinivasan, R.; Wu, Z.; Calo, E.; Acampora, D.; Faial, T.; Simeone, A.; Tan, M.; Swigut, T.; Wysocka, J. Reorganization of enhancer patterns in transition from naive to primed pluripotency. Cell Stem Cell 2014, 14, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Schulz, E.G.; Meisig, J.; Nakamura, T.; Okamoto, I.; Sieber, A.; Picard, C.; Borensztein, M.; Saitou, M.; Bluthgen, N.; Heard, E. The two active X chromosomes in female ESCs block exit from the pluripotent state by modulating the ESC signaling network. Cell Stem Cell 2014, 14, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Pirouz, M.; Choi, J.; Huebner, A.J.; Clement, K.; Meissner, A.; Hochedlinger, K.; Gregory, R.I. An Intermediate Pluripotent State Controlled by MicroRNAs Is Required for the Naive-to-Primed Stem Cell Transition. Cell Stem Cell 2018, 22, 851–864. [Google Scholar] [CrossRef]
- Lipchina, I.; Studer, L.; Betel, D. The expanding role of miR-302-367 in pluripotency and reprogramming. Cell Cycle 2012, 11, 1517–1523. [Google Scholar] [CrossRef]
- Judson, R.L.; Babiarz, J.E.; Venere, M.; Blelloch, R. Embryonic stem cell-specific microRNAs promote induced pluripotency. Nat. Biotechnol. 2009, 27, 459–461. [Google Scholar] [CrossRef]
- 199 Jouneau, A.; Ciaudo, C.; Sismeiro, O.; Brochard, V.; Jouneau, L.; Vandormael-Pournin, S.; Coppee, J.Y.; Zhou, Q.; Heard, E.; Antoniewski, C.; et al. Naive and primed murine pluripotent stem cells have distinct miRNA expression profiles. RNA 2012, 18, 253–264. [Google Scholar] [CrossRef]
- 200 Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 443–448. [Google Scholar] [CrossRef]
- Qi, J.; Yu, J.Y.; Shcherbata, H.R.; Mathieu, J.; Wang, A.J.; Seal, S.; Zhou, W.; Stadler, B.M.; Bourgin, D.; Wang, L.; et al. microRNAs regulate human embryonic stem cell division. Cell Cycle 2009, 8, 3729–3741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Export in 5, and DICER in microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef]
- Malnou, E.C.; Umlauf, D.; Mouysset, M.; Cavaille, J. Imprinted MicroRNA Gene Clusters in the Evolution, Development, and Functions of Mammalian Placenta. Front. Genet. 2018, 9, 706. [Google Scholar] [CrossRef] [PubMed]
- Noguer-Dance, M.; Abu-Amero, S.; Al-Khtib, M.; Lefevre, A.; Coullin, P.; Moore, G.E.; Cavaille, J. The primate-specific microRNA gene cluster (C19MC) is imprinted in the placenta. Hum. Mol. Genet. 2010, 19, 3566–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme-Axford, E.; Donker, R.B.; Mouillet, J.F.; Chu, T.; Bayer, A.; Ouyang, Y.; Wang, T.; Stolz, D.B.; Sarkar, S.N.; Morelli, A.E.; et al. Human placental trophoblasts confer viral resistance to recipient cells. Proc. Natl. Acad. Sci. USA 2013, 110, 12048–12053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Huang, F.; Wu, G.; Han, T.; Xu, F.; Weng, D.; Wu, C.; Zhang, X.; Yao, Y.; Zhu, X. MiR-519d-3p suppresses invasion and migration of trophoblast cells via targeting MMP-2. PLoS ONE 2015, 10, e0120321. [Google Scholar] [CrossRef]
- da Rocha, S.T.; Edwards, C.A.; Ito, M.; Ogata, T.; Ferguson-Smith, A.C. Genomic imprinting at the mammalian Dlk1-Dio3 domain. Trends Genet. 2008, 24, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.A.; Mungall, A.J.; Matthews, L.; Ryder, E.; Gray, D.J.; Pask, A.J.; Shaw, G.; Graves, J.A.; Rogers, J.; consortium, S.; et al. The evolution of the DLK1-DIO3 imprinted domain in mammals. PLoS Biol. 2008, 6, e135. [Google Scholar] [CrossRef] [PubMed]
- Niakan, K.K.; Ji, H.; Maehr, R.; Vokes, S.A.; Rodolfa, K.T.; Sherwood, R.I.; Yamaki, M.; Dimos, J.T.; Chen, A.E.; Melton, D.A.; et al. Sox17 promotes differentiation in mouse embryonic stem cells by directly regulating extraembryonic gene expression and indirectly antagonizing self-renewal. Genes Dev. 2010, 24, 312–326. [Google Scholar] [CrossRef] [Green Version]
- Rugg-Gunn, P.J.; Cox, B.J.; Lanner, F.; Sharma, P.; Ignatchenko, V.; McDonald, A.C.; Garner, J.; Gramolini, A.O.; Rossant, J.; Kislinger, T. Cell-surface proteomics identifies lineage-specific markers of embryo-derived stem cells. Dev. Cell 2012, 22, 887–901. [Google Scholar] [CrossRef]
- Qu, X.B.; Pan, J.; Zhang, C.; Huang, S.Y. Sox17 facilitates the differentiation of mouse embryonic stem cells into primitive and definitive endoderm in vitro. Dev. Growth Differ. 2008, 50, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Kanai-Azuma, M.; Hara, K.; Miyazaki, S.; Kanai, Y.; Monden, M.; Miyazaki, J. Sox17 plays a substantial role in late-stage differentiation of the extraembryonic endoderm in vitro. J. Cell Sci. 2007, 120, 3859–3869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deglincerti, A.; Croft, G.F.; Pietila, L.N.; Zernicka-Goetz, M.; Siggia, E.D.; Brivanlou, A.H. Self-organization of the in vitro attached human embryo. Nature 2016, 533, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.N.; Jedrusik, A.; Vuoristo, S.; Recher, G.; Hupalowska, A.; Bolton, V.; Fogarty, N.N.M.; Campbell, A.; Devito, L.; Ilic, D.; et al. Self-organization of the human embryo in the absence of maternal tissues. Nat. Cell Biol. 2016, 18, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Iwatani, C.; Tsuchiya, H.; Seita, Y.; Nakamura, S.; Shiraki, N.; Takakuwa, T.; et al. The Germ Cell Fate of Cynomolgus Monkeys Is Specified in the Nascent Amnion. Dev. Cell 2016, 39, 169–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Dicer-dependent biogenesis of miRNAs (A) and endo-siRNAs (B). (A) Biogenesis of miRNAs can be microprocessor- and DICER-dependent (monocistronic and polycistronic miRNAs), or microprocessor-independent and DICER-dependent (mirtrons), with very few exceptions. For the former, monocistronic and polycistronic miRNAs are transcribed by RNA polymerase II as primary miRNAs (pri-miRNAs), which are processed by the microprocessor complex (DROSHA and DGCR8) and exported to the cytoplasm as precursor miRNAs (pre-miRNAs) for the further processing by DICER. Mirtrons, in contrast, are located in the intron and are generated by splicing and trimming that do not need the microprocessor complex to form pre-miRNAs. In both cases, pre-miRNAs need DICER for forming short mature miRNA duplexes. One strand of the duplex is then loaded onto the RNA-interference complex (RISC), where the miRNA recognizes its target mRNA through imperfect base pairing, especially the complementation between the short “seed” sequence of the miRNA and its mRNA targets, performing post-transcriptional silencing on target mRNAs through degradation or translational repression. (B) The biogenesis of endo-siRNAs starts with the formation of duplexes from one or two transcripts with complementary sequences. Duplexes are exported to the cytoplasm and processed by DICER as well. Different from miRNAs, endo-siRNAs duplex with their target mRNAs with a higher degree of complementation, inducing the splicer activity of Ago2 for the cleavage of mRNAs, leading to its degradation. Please note that miRNA RISC can be formed with Ago1–4, while the RISC for endo-siRNAs contains Ago2.
Figure 1.
Dicer-dependent biogenesis of miRNAs (A) and endo-siRNAs (B). (A) Biogenesis of miRNAs can be microprocessor- and DICER-dependent (monocistronic and polycistronic miRNAs), or microprocessor-independent and DICER-dependent (mirtrons), with very few exceptions. For the former, monocistronic and polycistronic miRNAs are transcribed by RNA polymerase II as primary miRNAs (pri-miRNAs), which are processed by the microprocessor complex (DROSHA and DGCR8) and exported to the cytoplasm as precursor miRNAs (pre-miRNAs) for the further processing by DICER. Mirtrons, in contrast, are located in the intron and are generated by splicing and trimming that do not need the microprocessor complex to form pre-miRNAs. In both cases, pre-miRNAs need DICER for forming short mature miRNA duplexes. One strand of the duplex is then loaded onto the RNA-interference complex (RISC), where the miRNA recognizes its target mRNA through imperfect base pairing, especially the complementation between the short “seed” sequence of the miRNA and its mRNA targets, performing post-transcriptional silencing on target mRNAs through degradation or translational repression. (B) The biogenesis of endo-siRNAs starts with the formation of duplexes from one or two transcripts with complementary sequences. Duplexes are exported to the cytoplasm and processed by DICER as well. Different from miRNAs, endo-siRNAs duplex with their target mRNAs with a higher degree of complementation, inducing the splicer activity of Ago2 for the cleavage of mRNAs, leading to its degradation. Please note that miRNA RISC can be formed with Ago1–4, while the RISC for endo-siRNAs contains Ago2.

Figure 2.
Progression of cell potencies in early embryos and cultured cells. The pre-implantation embryonic development starts with fertilized oocytes, followed by the zygotic gene activation at the 4-cell stage and 2-cell stage blastomeres in humans and mice, respectively (A). Zygotes and 2-/4-cell stage blastomeres are totipotent, forming the whole fetus, including embryonic and extraembryonic lineages that will give rise to the yolk sac and placenta in the future. Through the morula stage, the embryonic and extraembryonic lineages are segregated in the early blastocyst, forming the inner cell mass (ICM) and the trophectoderm (TE) (B). Pluripotent stem cells are emerged from the ICM and further separated from the primitive endoderm (PrE) to form the epiblast (EPI) in the late blastocyst (C). These three main lineages (TE, PrE, and EPI) can give rise to three types of stem cells (trophoblast stem cells [TSC], extraembryonic endoderm [XEN] stem cells, and embryonic stem cells [ESC]) with corresponding cell potencies. In humans, the culture condition for XEN stem cells remains to be discovered. Both EPIs and ESCs are considered to be in the “naïve” or “ground” state and need to be “primed” for further differentiation. In utero, this priming happens after implantation, leading to the reduced pluripotency of the EPI in post-implantation embryos (D). In vitro, primed pluripotent stem cells can be directly derived from epiblasts of post-implantation embryos or converted from the naïve ESCs as EpiSC-like cells (EpiLCs) (E). Finally, the very first germ cell lineage, primordial germ cells (PGCs), are specified after implantation. Since PGCs are either impractical to be isolated (in humans) or difficult to maintain as the primary culture, they are usually substituted by converting the pluripotent stem cells to PGC-like cells, PGCLCs (F). The black dashed line indicates the corresponding cell potency.
Figure 2.
Progression of cell potencies in early embryos and cultured cells. The pre-implantation embryonic development starts with fertilized oocytes, followed by the zygotic gene activation at the 4-cell stage and 2-cell stage blastomeres in humans and mice, respectively (A). Zygotes and 2-/4-cell stage blastomeres are totipotent, forming the whole fetus, including embryonic and extraembryonic lineages that will give rise to the yolk sac and placenta in the future. Through the morula stage, the embryonic and extraembryonic lineages are segregated in the early blastocyst, forming the inner cell mass (ICM) and the trophectoderm (TE) (B). Pluripotent stem cells are emerged from the ICM and further separated from the primitive endoderm (PrE) to form the epiblast (EPI) in the late blastocyst (C). These three main lineages (TE, PrE, and EPI) can give rise to three types of stem cells (trophoblast stem cells [TSC], extraembryonic endoderm [XEN] stem cells, and embryonic stem cells [ESC]) with corresponding cell potencies. In humans, the culture condition for XEN stem cells remains to be discovered. Both EPIs and ESCs are considered to be in the “naïve” or “ground” state and need to be “primed” for further differentiation. In utero, this priming happens after implantation, leading to the reduced pluripotency of the EPI in post-implantation embryos (D). In vitro, primed pluripotent stem cells can be directly derived from epiblasts of post-implantation embryos or converted from the naïve ESCs as EpiSC-like cells (EpiLCs) (E). Finally, the very first germ cell lineage, primordial germ cells (PGCs), are specified after implantation. Since PGCs are either impractical to be isolated (in humans) or difficult to maintain as the primary culture, they are usually substituted by converting the pluripotent stem cells to PGC-like cells, PGCLCs (F). The black dashed line indicates the corresponding cell potency.

Figure 3.
Major miRNA clusters expressed in embryonic stem cells. (A) Mouse miR-290~295 cluster and human miR-371~373 are homologous miRNAs. Except for miR-293 in mice and miR-371 in humans, all miRNAs contain the “AAGUGC” motif in seed sequences (marked in red). (B) The sequence alignment of mouse and human miR-302~367 clusters, which are highly conserved and also contain the “AAGUGC” motif. (C,D) The structure and sequence alignment of miR-17~92, miR-106a~363, and miR-106b~25 clusters. Please note that the miR-17 family contains the full “AAGUGC”, while the miR-18, -19, and -92 families only contain a part of the “AAGUGC” motif.
Figure 3.
Major miRNA clusters expressed in embryonic stem cells. (A) Mouse miR-290~295 cluster and human miR-371~373 are homologous miRNAs. Except for miR-293 in mice and miR-371 in humans, all miRNAs contain the “AAGUGC” motif in seed sequences (marked in red). (B) The sequence alignment of mouse and human miR-302~367 clusters, which are highly conserved and also contain the “AAGUGC” motif. (C,D) The structure and sequence alignment of miR-17~92, miR-106a~363, and miR-106b~25 clusters. Please note that the miR-17 family contains the full “AAGUGC”, while the miR-18, -19, and -92 families only contain a part of the “AAGUGC” motif.


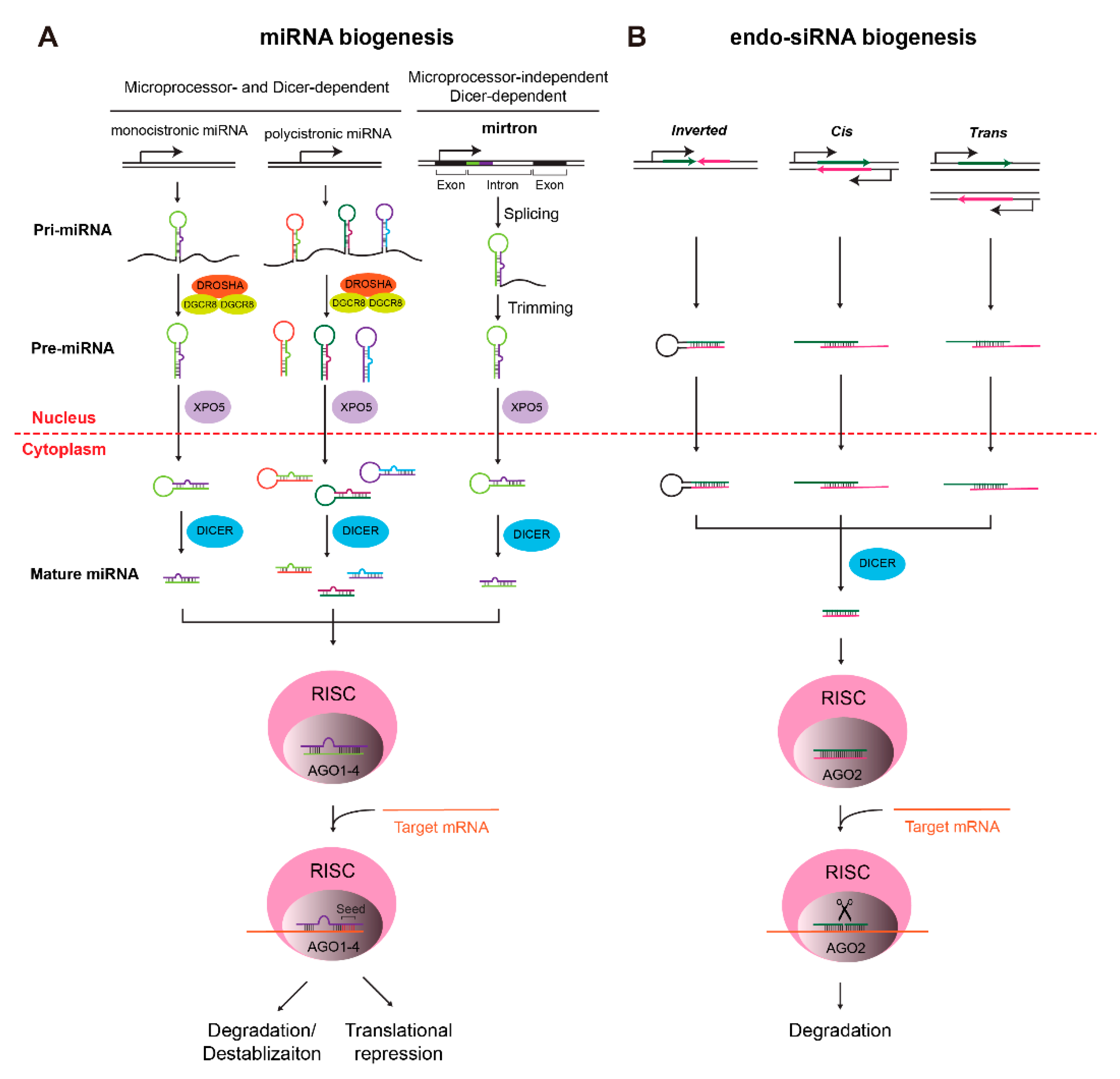{kind=link}
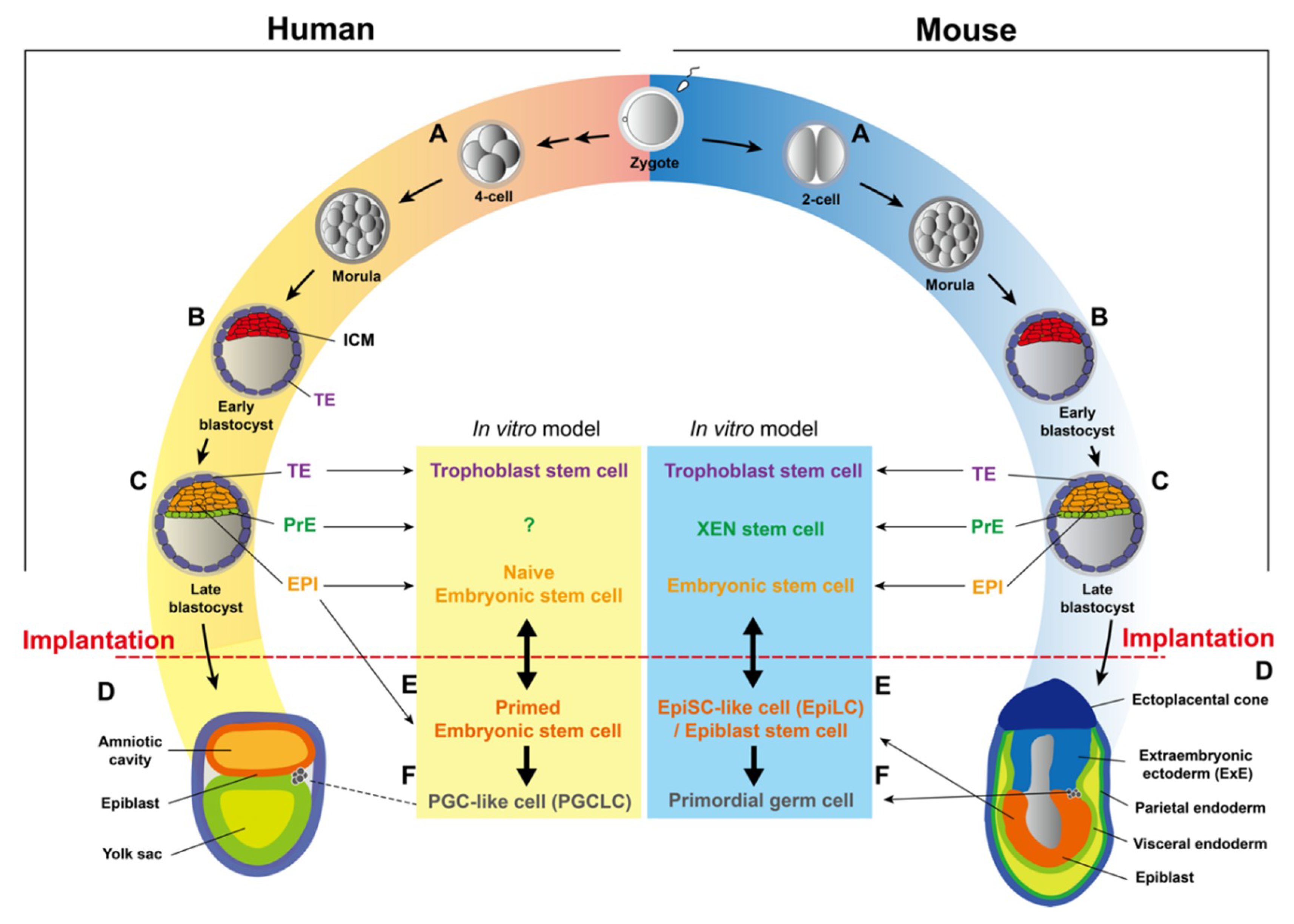{kind=link}
{kind=link}
Table 1.
Studies of specific miRNAs in knockout mouse models.
| Knockout Mouse Model | Perinatal Phenotype | Phenotype at Embryonic Stages | Target | Ref. |
|---|---|---|---|---|
| miR-17~92−/− | Early postnatal lethality and very specific defects in the development of heart, lungs, and B cells | Smaller size of embryos at E13.5 and E18.5; ventricular septal defects in heart at E18.5; severely hypoplastic lungs from E18.5–P0; greatly reduced percentage and absolute number of pre-B cells at E18.5 | bim | [153] |
| miR-17~92Δ17, miR-17~92Δ18, miR-17~92Δ19, miR-17~92Δ92, miR-17~92Δ17,18 and miR-17~92Δ17,18,92 | Depletion of individual or multiple miRNAs in this cluster led to variable phenotypes. Perinatal lethality and lung hypoplasia only in miR-17~92Δ17,18,92/Δ17,18,92 mice; Co-deletion of miR-17 and miR-18 has significant weight-reduction; miR-17~92Δ17/Δ17 mice show reduction of pre-B-cells. | miR-17~92Δ17,18/Δ17,18 mice display fusion of the proximal carpal bones. | tbx3, tbx20, smad6, heg1, klf2 and trip11 | [154] |
| miR-106b~25−/−; miR-17~92−/− double knockout | NA | Embryos die before E15 with much more severe phenotypes compared to embryos lacking miR-17~92 alone; edema and vascular congestion at E13.5 and E14.5 | bim | [153] |
| miR-17~9−/−; miR-106~2−/−; miR-106a~363−/− triple-knockout | NA | Embryos die before E15 with much more severe phenotype compared to embryos lacking miR-17~92 alone | bim | [153] |
| miR-290~295−/− | Germ cell deficiency in surviving adults | Partially penetrant embryonic lethality; miR-290~295−/− blastocysts at E3.5 exhibit no obvious phenotype; miR-290~295−/− embryo were lost between E11.5 and E18.5; at E10.5, about 16% of miR-290~295−/− embryos were partially or completely localized outside the yolk sac | NA | [126] |
| miR-290~295−/− | NA | Defects in placental growth; prematurely exit the cell cycle of trophoblast progenitor cells; reduced endoreduplicaton of TGCs; disorganized placenta with the reduced area of vasculature; Reduced diffusional exchange capacity | Multiple (combinationational effect) | [152] |
| miR-126−/− | Vascular abnormalities | About 40% of miR-126−/− mice died embryonically or perinatally with angiogenesis defects | spred-1 | [155] |
| miR-205−/− | Neonatal lethality with compromised epidermal and hair follicle growth | NA | phlda3 and inppl1 | [156] |
| miR-1~2−/− | Lethality at weaning with dysregulation of cardiogenesis | NA | NA | [157] |
| miR-34a−/−; miR-34b/34c−/−; miR-449−/− triple knockout | 60% animals lethal and surviving adults are infertile; respiratory dysfunction | NA | cp110 | [158] |
| miR-302 −/− and miR-302 −/−; miR-290 −/− double knockout | NA | Embryos are normal at E7.5; grossly abnormal in neural development at E9.5; severely abnormal brain development at E13.5; double knockouts arrest prior to neurulation | fgf15 | [159] |
| miR-200b −/−; miR-429−/− double knockout | Female infertility | NA | zeb1 | [160] |
| miR-12−/−- | Increased motor activity and fatal epilepsy | NA | slc6a1, slc1a1, scn2b, scn4b, cacna2d3, cagn2, car7 | [161] |
| miR-29ab1−/− | Shorter life span; reduced lymphoid organ cellularity | NA | t-bet and ifn-gamma | [162] |
| miR-133a-1−/−; miR-133a-2−/− double knockout | 50% perinatal lethality at P0 and P1 with ventricular septal defects; cardiomyopathy and heart failure | Abnormal heart development from E12.5 to E17.5 | srf and cyclin d2 | [163] |
Abbreviations: NA, not available or not addressed.
Table 2.
Studies of specific miRNAs in in vitro cell culture models.
| Species | Cell | Cell Potential | miRNA | Target | Phenotype | Ref. |
|---|---|---|---|---|---|---|
| Mouse | ESC | Naïve pluripotency | miR-34a | gata2 | miR-34a restricts the acquisition of expanded cell fate potential in pluripotent stem cells. | [168] |
| Mouse | ESC | Naïve pluripotency | miR-290~295 | NA | miR-290~295 knockout ESCs are pluripotent and no obvious phenotypes were observed. | [173] |
| Mouse | ESC | Naïve pluripotency | miR-290~295 | casp2, ei24 | Overexpression of miR-294 rescues the apoptosis phenotype of dicer−/− ESCs upon genotoxic stress. Deletion of miR-295 cluster enhances the susceptibility of ESCs toward apoptosis upon DNA damage. | [176] |
| Mouse | ESC | Naïve pluripotency | miR-290, miR-183~182 | miR-290 targets rbl2, cdkn1a, lats2, mbd2, etc. miR-183~182 may have different targets but not elucidated | miR-294, miR-182 and miR-183 antagonize the differentiation of dgcr8−/− ESCs induced by let-7. miR-290−/−; miR-183−/− double knockout ESCs are susceptible for let-7 induced differentiation. | [177] |
| Mouse | ESC | Naïve pluripotency | miR-294/302 | tgfbr1, tgfbr2, gsk3b | miR-294/302 combinatorically suppress epithelial–mesenchymal transition (EMT) and apoptosis in differentiating dgcr8−/− ESCs induced by let-7. | [178] |
| Mouse | ESC | Naïve pluripotency | miR-26a, miR-99b, miR-193, miR-199a-5p, and miR-218 | NA | Similar to let-7, overexpression of these five miRNAs induces differentiation in dgcr8−/− ESCs | [179] |
| Mouse | ESC | Naïve pluripotency | miR-291a/b-3p, miR-294, miR-295, and miR-302 (embryonic stem cell specific, or ESCC miRNAs) | cdkn1a, rbl2, lats2 | Overexpression of ESCC miRNAs promotes cell cycle progression and oppose let-7-induced differentiation in dgcr8−/− ESCs. | [137,180] |
| let-7 | lin28, cmyc, nmyc, sall4 | Overexpression of let-7 induces differentiation in dgcr8−/− ESCs. | ||||
| Mouse | ESC | Naïve pluripotency | miR-320, miR-702 (microprocessor-independent miRNAs) | p57, p21 | Overexpression of miR-320 or miR-702 rescues proliferation defects of dicer−/− and dgcr8−/− ESCs. | [181] |
| Mouse | EpiLC | Primed pluripotency | miR-302 single knockout, miR-290 and miR-302 double knockout | akt1 | Double knockout of miR-290/302 leads to the improper transition from ESCs to EpiLCs; overexpression of miRNAs in two clusters rescues the phenotype of dgcr8−/− ESCs. | [175] |
| Mouse | EpiSC | Primed pluripotency | miR-127 | lefty2 | Inhibiting miR-127 expression in EpiSCs results in decreasing mesendodermal differentiation. | [182] |
| Mouse | EpiSC | Primed pluripotency | miR-20, miR-92, miR-302 | bim | Overexpression of either of three miRNAs rescues the apoptosis phenotype of dicer−/− EpiSCs. | [183] |
| Mouse | TSC | Extraembryonic | miR-290~295 | p21, p57, rbl2, and lats2 | Depletion of dicer in TSCs results in the differentiation and proliferation block of mTSCs. | [128] |
| Mouse | XEN | Extraembryonic | miR-20a, miR-30b | rasa2 | Deletion of dicer in XEN stem cells leads to differentiation and proliferation block. | [128] |
| miR-669a | dusp1 | |||||
| Mouse | PGC-like | Germline | let-7 | blimp1 | let-7 blocks the transition of ESCs to PGCs. | [184] |
| Human | ESC | Primed | miR-302~367, miR-371~373 | fas, nik, trailr4, and bim | Overexpression of either of two miRNA clusters rescues the apoptosis phenotype in DICER−/− ESCs. | [173] |
| Human | PGCLC | Germline | miR-372 | smarcc1, mecp2, cdkn1, rbl2, rhoc, and tgfbr2 | miR-372 promotes PGC specification. | [185] |
| let-7 | cmyc and nmyc | let-7 inhibits PGC specification. | [185] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Z.; Zhuang, L.; Lin, C.-P. Roles of MicroRNAs in Establishing and Modulating Stem Cell Potential. Int. J. Mol. Sci. 2019, 20, 3643. https://doi.org/10.3390/ijms20153643
AMA Style
Zhang Z, Zhuang L, Lin C-P. Roles of MicroRNAs in Establishing and Modulating Stem Cell Potential. International Journal of Molecular Sciences. 2019; 20(15):3643. https://doi.org/10.3390/ijms20153643
Chicago/Turabian StyleZhang, Zhenwu, Lili Zhuang, and Chao-Po Lin. 2019. "Roles of MicroRNAs in Establishing and Modulating Stem Cell Potential" International Journal of Molecular Sciences 20, no. 15: 3643. https://doi.org/10.3390/ijms20153643
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.