The Role of Prep1 in the Regulation of Mesenchymal Stromal Cells

 ,
,  and
and
Abstract
:1. Introduction
2. Results
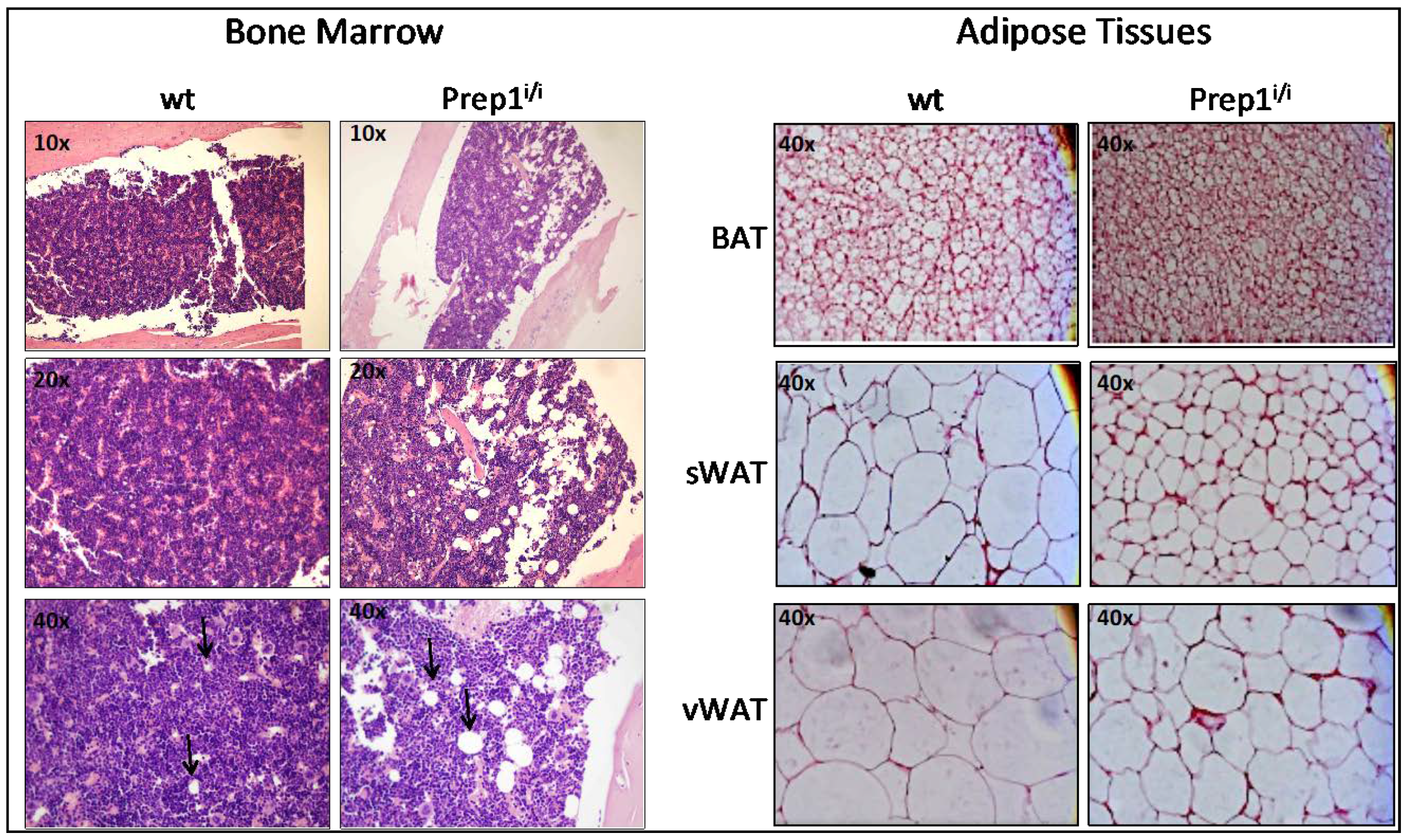
2.1. Histologic Analysis of BM and Fat Tissues
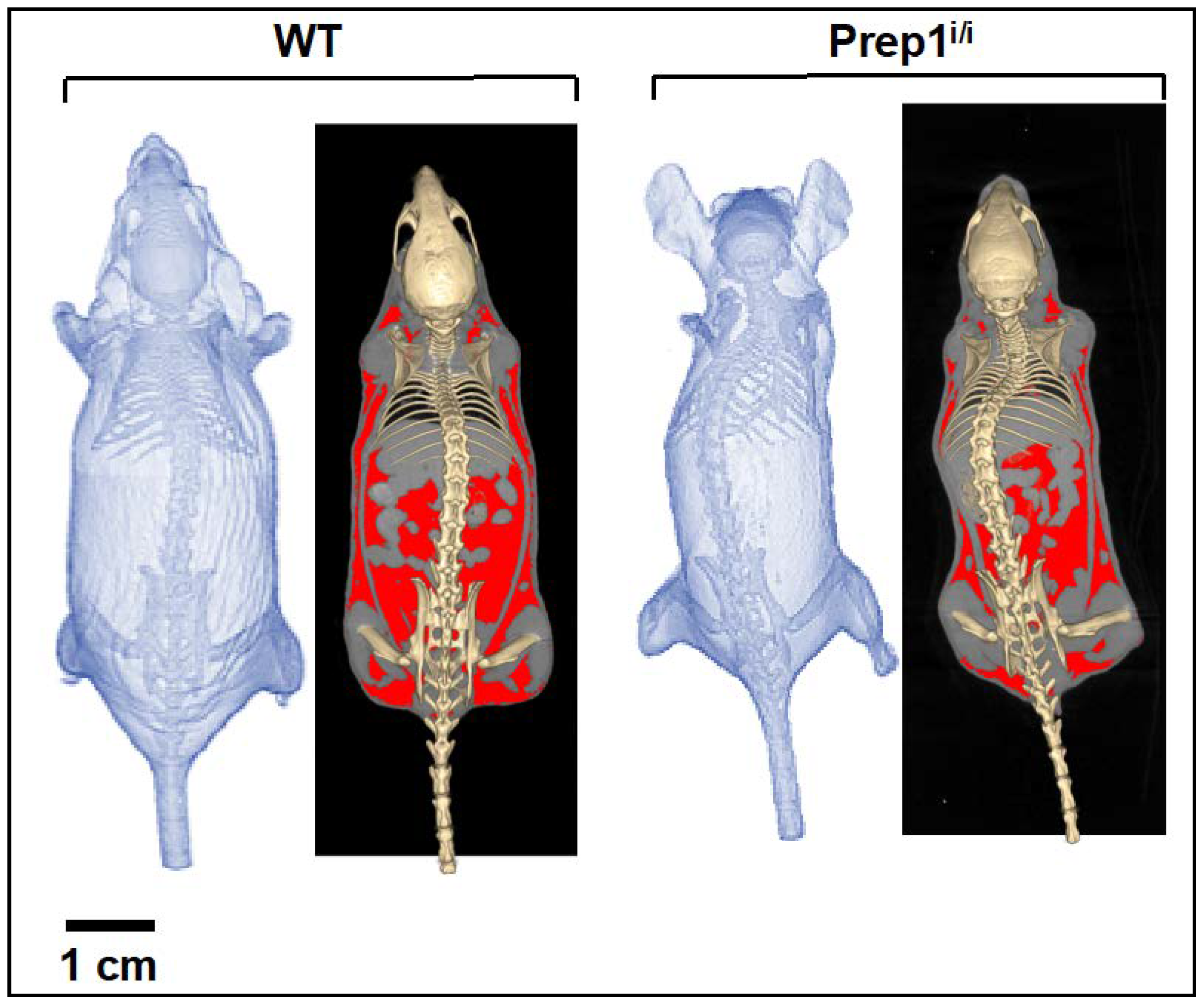
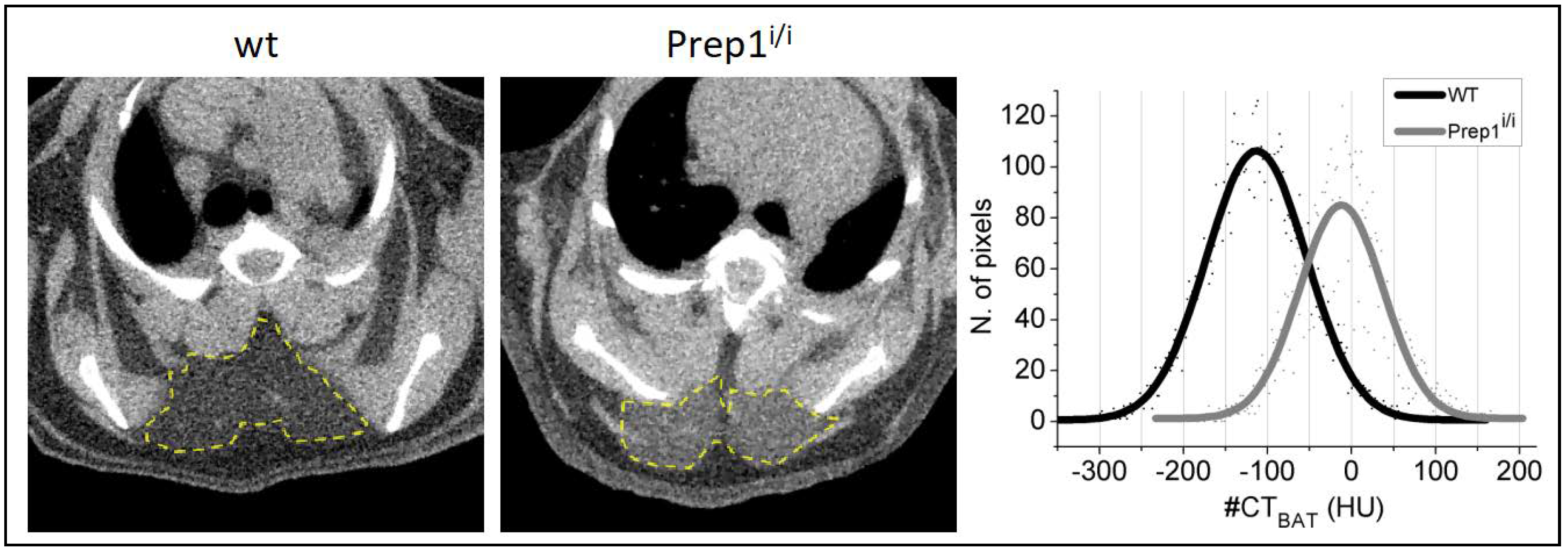
2.2. Whole Body Analysis by Micro-CT Imaging Techniques of WT and Prep1i/i Mice
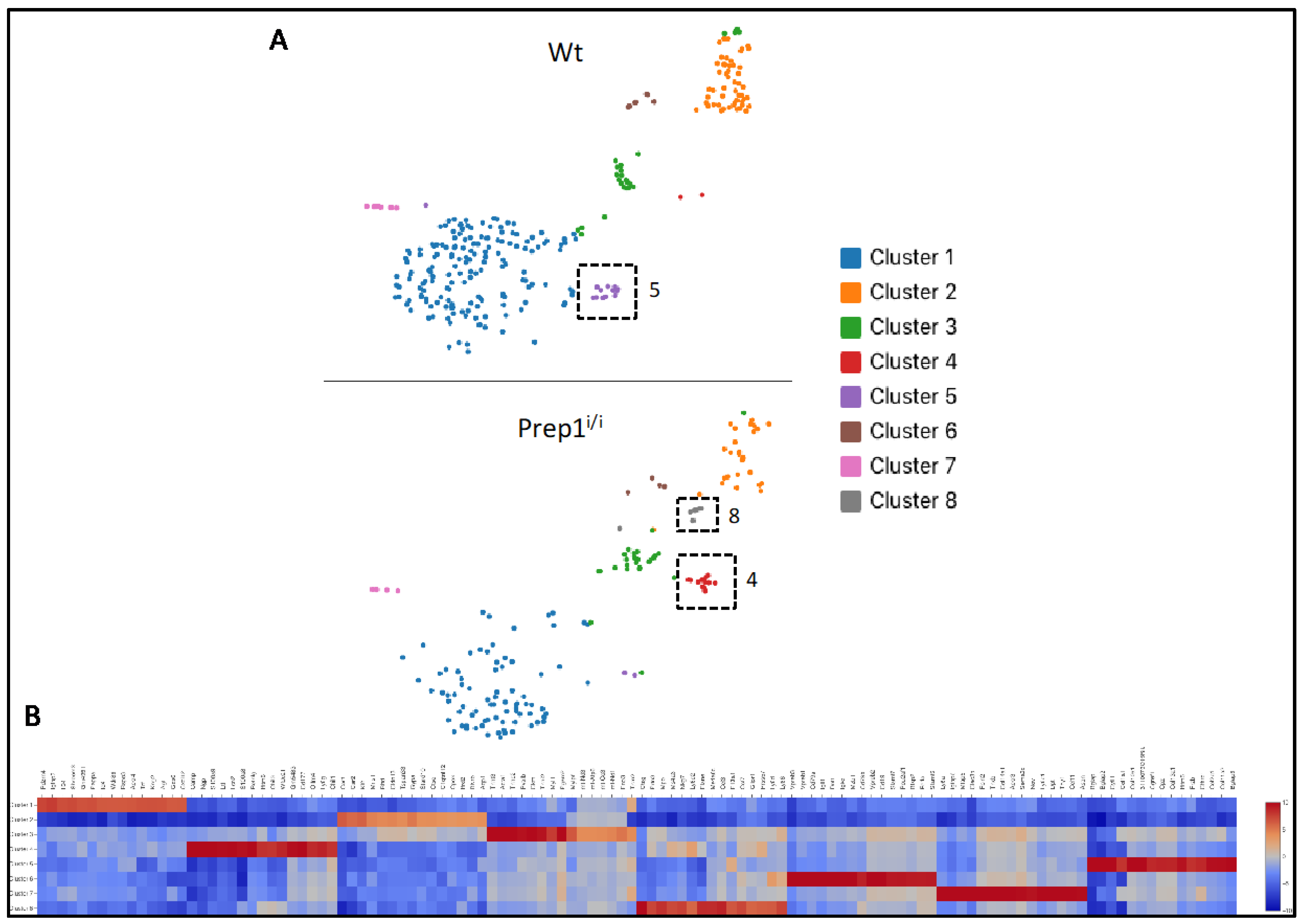
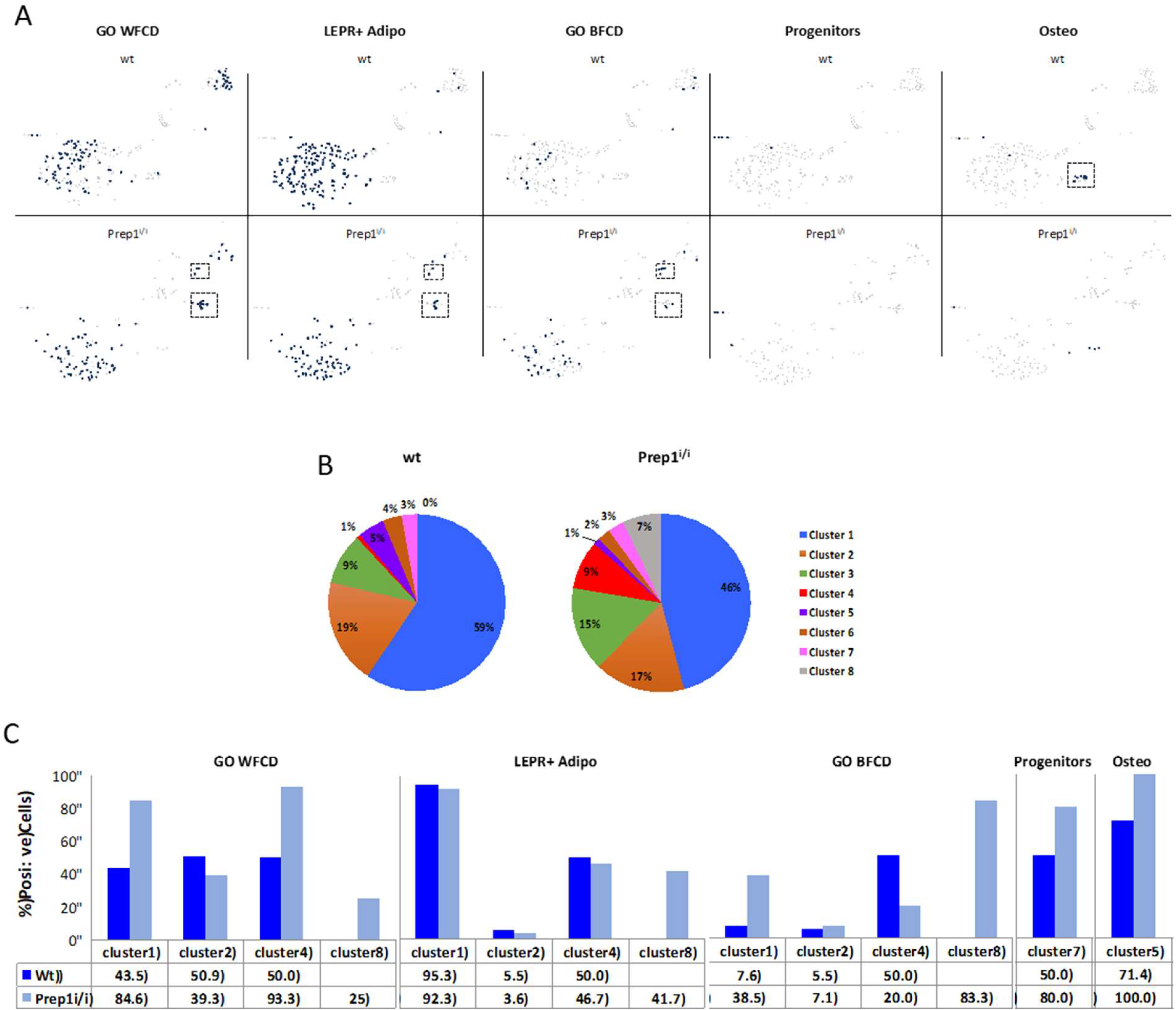
2.3. Single Cell Transcriptomics of BM Mesenchymal Stromal Cells
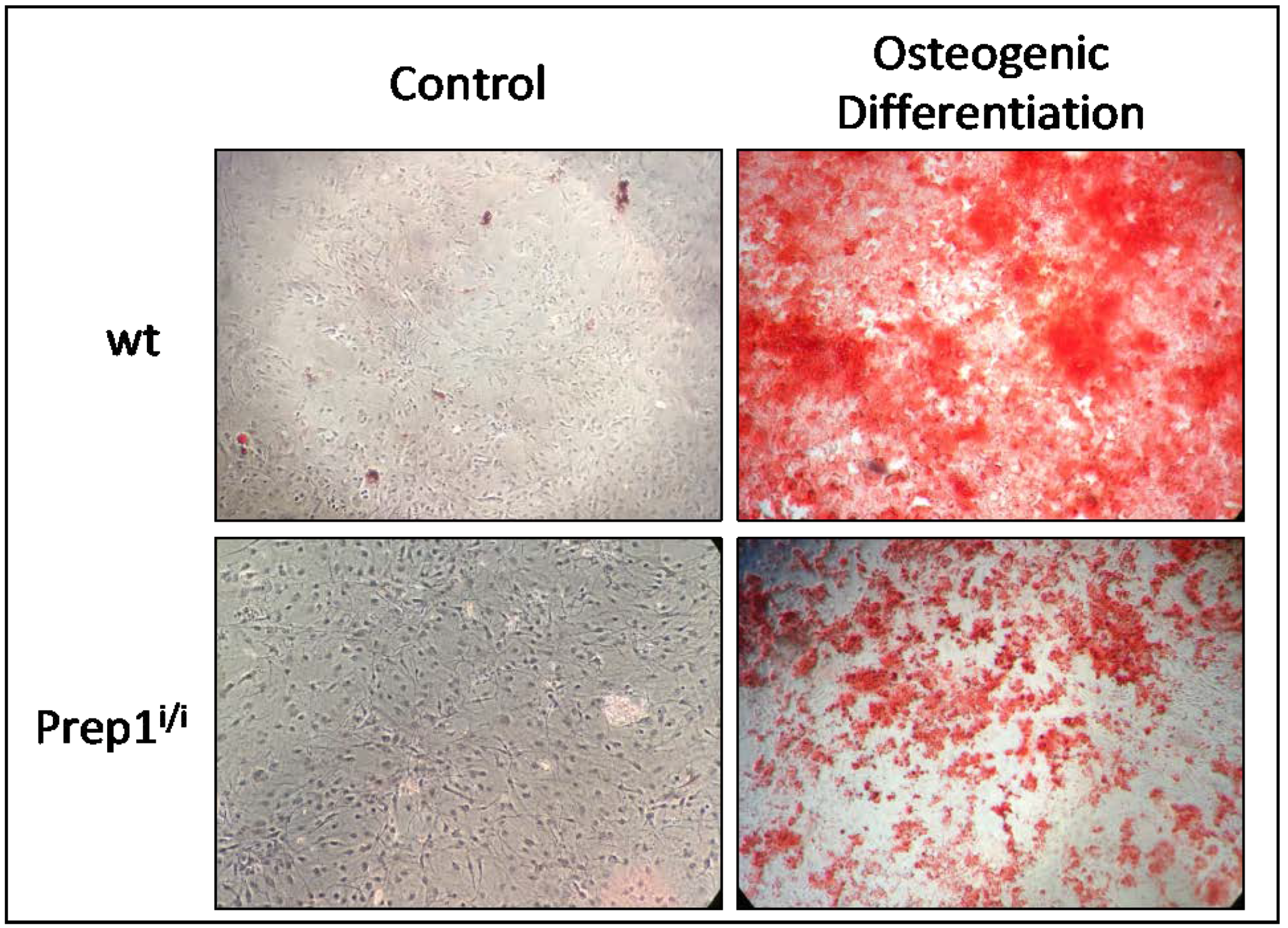
2.4. Comparative In Vitro Studies on Osteogenic Differentiation of wt and Prep1i/i MSCs
3. Discussion
4. Materials and Methods
4.1. Histological Analysis of Bone Marrow and Adipose Tissues
4.1.1. Bone Marrow
4.1.2. Adipose Tissues
4.2. In Vitro Cultures of Mesenchymal Stromal Cells (MSCs)
4.3. Osteogenic Differentiation
4.4. Single Cell RNA Sequencing and Bioinformatics Analysis
4.5. Whole-Body Micro-CT Imaging and Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fernandez-Diaz, L.C.; Laurent, A.; Girasoli, S.; Turco, M.; Longobardi, E.; Iotti, G.; Jenkins, N.A.; Fiorenza, M.T.; Copeland, N.G.; Blasi, F. The absence of Prep1 causes p53-dependent apoptosis of mouse pluripotent epiblast cells. Development 2010, 137, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, E.; Villaescusa, J.C.; Di Rosa, P.; Fernandez-Diaz, L.C.; Longobardi, E.; Mazzieri, R.; Miccio, A.; Micali, N.; Selleri, L.; Ferrari, G.; et al. Hypomorphic mutation of the TALE gene Prep1 (pKnox1) causes a major reduction of Pbx and Meis proteins and a pleiotropic embryonic phenotype. Mol. Cell Biol. 2006, 26, 5650–5662. [Google Scholar] [CrossRef] [PubMed]
- Penkov, D.; Di Rosa, P.; Fernandez Diaz, L.; Basso, V.; Ferretti, E.; Grassi, F.; Mondino, A.; Blasi, F. Involvement of Prep1 in the alphabeta T-cell receptor T-lymphocytic potential of hematopoietic precursors. Mol. Cell Biol. 2005, 25, 10768–10781. [Google Scholar] [CrossRef] [PubMed]
- Longobardi, E.; Iotti, G.; Di Rosa, P.; Mejetta, S.; Bianchi, F.; Fernandez-Diaz, L.C.; Micali, N.; Nuciforo, P.; Lenti, E.; Ponzoni, M.; et al. Prep1 (pKnox1)-deficiency leads to spontaneous tumor development in mice and accelerates EmuMyc lymphomagenesis: A tumor suppressor role for Prep1. Mol. Oncol. 2010, 4, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, P.; Villaescusa, J.C.; Longobardi, E.; Iotti, G.; Ferretti, E.; Diaz, V.M.; Miccio, A.; Ferrari, G.; Blasi, F. The homeodomain transcription factor Prep1 (pKnox1) is required for hematopoietic stem and progenitor cell activity. Dev. Biol. 2007, 311, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Modica, L.; Iotti, G.; D’Avola, A.; Blasi, F. Prep1 (pKnox1) regulates mouse embryonic HSC cycling and self-renewal affecting the Stat1-Sca1 IFN-dependent pathway. PLoS ONE 2014, 9, e107916. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, K.; Oda, A.; Notsu, C.; Ohtsuka, T.; Kawai, Y.; Suzuki, S.; Nakamura, T.; Mabuchi, Y.; Matsuzaki, Y.; Goitsuka, R. Loss of the Homeodomain Transcription Factor Prep1 Perturbs Adult Hematopoiesis in the Bone Marrow. PLoS ONE 2015, 10, e0136107. [Google Scholar] [CrossRef]
- Friedenstein, A.J. Stromal mechanisms of bone marrow: Cloning in vitro and retransplantation in vivo. Haematol Blood Transfus. 1980, 25, 19–29. [Google Scholar]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.Y. The meaning, the sense and the significance: Translating the science of mesenchymal stem cells into medicine. Nat. Med. 2013, 19, 35–42. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. Adult Mesenchymal Stem Cells: When, Where, and How. Stem Cells Int. 2015, 2015, 628767. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Ferrer, S.; Scadden, D.T.; Sanchez-Aguilera, A. Bone marrow stem cells: Current and emerging concepts. Ann. N. Y. Acad. Sci. 2015, 1335, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Frenette, P.S. Niches for Hematopoietic Stem Cells and Their Progeny. Immunity 2018, 48, 632–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahara, F.; Borger, D.K.; Wei, Q.; Pinho, S.; Maryanovich, M.; Zahalka, A.H.; Suzuki, M.; Cruz, C.D.; Wang, Z.; Xu, C.; et al. Engineering a haematopoietic stem cell niche by revitalizing mesenchymal stromal cells. Nat. Cell Biol. 2019, 21, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Maroni, G.; Tkachuk, V.A.; Egorov, A.; Morelli, M.J.; Luongo, R.; Levantini, E.; Blasi, F.; Magli, M.C.; Penkov, D. Prep1 prevents premature adipogenesis of mesenchymal progenitors. Sci Rep. 2017, 7, 15573. [Google Scholar] [CrossRef] [PubMed]
- Oriente, F.; Perruolo, G.; Cimmino, I.; Cabaro, S.; Liotti, A.; Longo, M.; Miele, C.; Formisano, P.; Beguinot, F. Prep1, A Homeodomain Transcription Factor Involved in Glucose and Lipid Metabolism. Front. Endocrinol. (Lausanne) 2018, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Borga, M.; Virtanen, K.A.; Romu, T.; Leinhard, O.D.; Persson, A.; Nuutila, P.; Enerback, S. Brown adipose tissue in humans: Detection and functional analysis using PET (positron emission tomography), MRI (magnetic resonance imaging), and DECT (dual energy computed tomography). Methods Enzymol. 2014, 537, 141–159. [Google Scholar] [PubMed]
- Tikhonova, A.N.; Dolgalev, I.; Hu, H.; Sivaraj, K.K.; Hoxha, E.; Cuesta-Dominguez, A.; Pinho, S.; Akhmetzyanova, I.; Gao, J.; Witkowski, M.; et al. The bone marrow microenvironment at single-cell resolution. Nature 2019, 569, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Houlihan, D.D.; Mabuchi, Y.; Morikawa, S.; Niibe, K.; Araki, D.; Suzuki, S.; Okano, H.; Matsuzaki, Y. Isolation of mouse mesenchymal stem cells on the basis of expression of Sca-1 and PDGFR-alpha. Nat. Protoc. 2012, 7, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, E.; Schulz, H.; Talarico, D.; Blasi, F.; Berthelsen, J. The PBX-regulating protein PREP1 is present in different PBX-complexed forms in mouse. Mech. Dev. 1999, 83, 53–64. [Google Scholar] [CrossRef]
- Horowitz, M.C.; Berry, R.; Holtrup, B.; Sebo, Z.; Nelson, T.; Fretz, J.A.; Lindskog, D.; Kaplan, J.L.; Ables, G.; Rodeheffer, M.S.; et al. Bone marrow adipocytes. Adipocyte 2017, 6, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez-Preitner, M.; Berney, X.; Uldry, M.; Vitali, A.; Cinti, S.; Ledford, J.G.; Thorens, B. Plac8 is an inducer of C/EBPbeta required for brown fat differentiation, thermoregulation, and control of body weight. Cell Metab. 2011, 14, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Wijayatunga, N.N.; Pahlavani, M.; Kalupahana, N.S.; Kottapalli, K.R.; Gunaratne, P.H.; Coarfa, C.; Ramalingam, L.; Moustaid-Moussa, N. An integrative transcriptomic approach to identify depot differences in genes and microRNAs in adipose tissues from high fat fed mice. Oncotarget 2018, 9, 9246–9261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchamp de Lageneste, O.; Julien, A.; Abou-Khalil, R.; Frangi, G.; Carvalho, C.; Cagnard, N.; Cordier, C.; Conway, S.J.; Colnot, C. Periosteum contains skeletal stem cells with high bone regenerative potential controlled by Periostin. Nat. Commun. 2018, 9, 773. [Google Scholar] [CrossRef] [PubMed]
- Baryawno, N.; Przybylski, D.; Kowalczyk, M.S.; Kfoury, Y.; Severe, N.; Gustafsson, K.; Kokkaliaris, K.D.; Mercier, F.; Tabaka, M.; Hofree, M.; et al. A Cellular Taxonomy of the Bone Marrow Stroma in Homeostasis and Leukemia. Cell 2019, 177, 1915–1932. [Google Scholar] [CrossRef] [PubMed]
- Kayaba, A.; Itoh-Nakadai, A.; Niibe, K.; Shirota, M.; Funayama, R.; Sugahara-Tobinai, A.; Wong, Y.L.; Inui, M.; Nakayama, K.; Takai, T. Bone marrow PDGFRalpha+Sca-1+-enriched mesenchymal stem cells support survival of and antibody production by plasma cells in vitro through IL-6. Int. Immunol. 2018, 30, 241–253. [Google Scholar] [CrossRef]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef]
- Liotti, A.; Cabaro, S.; Cimmino, I.; Ricci, S.; Procaccini, C.; Paciello, O.; Raciti, G.A.; Spinelli, R.; Iossa, S.; Matarese, G.; et al. Prep1 deficiency improves metabolic response in white adipose tissue. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 515–525. [Google Scholar] [CrossRef]
- Giralt, M.; Villarroya, F. Mitochondrial Uncoupling and the Regulation of Glucose Homeostasis. Curr. Diabetes Rev. 2017, 13, 386–394. [Google Scholar] [CrossRef]
- Oriente, F.; Fernandez Diaz, L.C.; Miele, C.; Iovino, S.; Mori, S.; Diaz, V.M.; Troncone, G.; Cassese, A.; Formisano, P.; Blasi, F.; et al. Prep1 deficiency induces protection from diabetes and increased insulin sensitivity through a p160-mediated mechanism. Mol. Cell Biol. 2008, 28, 5634–5645. [Google Scholar] [CrossRef]
- Schneider, W.; Bortfeld, T.; Schlegel, W. Correlation between CT numbers and tissue parameters needed for Monte Carlo simulations of clinical dose distributions. Phys. Med. Biol. 2000, 45, 459–478. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT (n = 16) | Prep1i/i (n = 14) | p-Value | |
|---|---|---|---|
| Weight (g) | |||
| mean (sd) | 32.232 ± 5.759 | 27.479 ± 4.022 | 0.029 (*) |
| min | 19.89 | 21.24 | |
| max | 40.21 | 32.78 | |
| Total volume (cm3) | |||
| mean (sd) | 31.421 ± 5.794 | 26.496 ± 4.132 | 0.026 (*) |
| min | 19.261 | 20.065 | |
| max | 39.37 | 32.23 | |
| Total fat volume (cm3) | |||
| mean (sd) | 7.641 ± 3.487 | 4.655 ± 2.367 | 0.029 (*) |
| min | 1.948 | 1.281 | |
| max | 14.330 | 7.724 | |
| Total bone volume (cm3) | |||
| mean (sd) | 1.411 ± 0.164 | 1.386 ± 0.169 | 1 |
| min | 1.155 | 1.112 | |
| max | 1.728 | 1.644 | |
| Length (mm) | |||
| mean (sd) | 96.511 ± 2.763 | 94.561 ± 2.505 | 0.044 (*) |
| min | 90.26 | 90.17 | |
| max | 101.80 | 98.54 | |
| % Fat | |||
| mean (sd) | 21.724 ± 7.614 | 15.540 ± 6.843 | 0.021 (*) |
| min | 6.74 | 5.72 | |
| max | 35.13 | 27.42 | |
| BAT volume (cm3) | |||
| mean (sd) | 0.106 ± 0.055 | 0.088 ± 0.042 | 0.57 |
| min | 0.043 | 0.045 | |
| max | 0.229 | 0.208 | |
| Bone radiodensity (HU) | |||
| mean (sd) | 1461 ± 113 | 1400 ± 165 | 0.22 |
| min | 1244 | 1204 | |
| max | 1661 | 1661 | |
| Fat radiodensity (HU) | |||
| mean (sd) | −260 ± 29 | −248 ± 25 | 0.41 |
| min | −335 | −278 | |
| max | −214 | −196 | |
| BAT radiodensity (HU) | |||
| mean (sd) | −180 ± 46 | −132 ± 36 | 0.013 (*) |
| min | −236 | −182 | |
| max | −91 | −54 | |
| GO WFCD | LEPR+ Adipo | GO BFCD | Progenitors | Osteo | |
|---|---|---|---|---|---|
| Cebpα | Hp | Adipoq | Mecom | PDGFRα | Cd200 |
| Ctbp1 | Lpl | Adrb1 | Metrnl | Ly6a | Col1a1 |
| Ctbp2 | Adipoq | Adrb2 | Mrap | Col1a2 | |
| Fabp4 | Slc1a5 | Adrb3 | Mtor | Alpl | |
| Fgf10 | Cd302 | Aldh6a1 | Napepld | Spp1 | |
| Ncor2 | Gas6 | Arl4a | Nudt7 | Sparc | |
| Per2 | Apoe | Bnip3 | Pex11a | Msx2 | |
| Pparγ | Lepr | Cebpα | Plac8 | Bglap | |
| Prdm16 | Cebpβ | Pparγ | Hox10 | ||
| Scd1 | Dusp10 | Pparγc1a | Sp7 | ||
| Sirt1 | Ebf2 | Prdm16 | Runx2 | ||
| Snai2 | Ero1l | Ptgs2 | |||
| Tbl1xr1 | Fabp4 | Rarres2 | |||
| Wfdc21 | Fndc5 | Rgs2 | |||
| Cebpδ | Fto | Scd1 | |||
| Cebpβ | Hnrnpu | Selenbp1 | |||
| Adipoq | Itga6 | Sh2b2 | |||
| Srebf1 | Lama4 | Sirt1 | |||
| Lamb3 | Slc2a4 | ||||
| Lep | Trpv4 | ||||
| Lrg1 | Vstm2a | ||||
| Mapk14 | Zbtb7b | ||||
| Mb | Zfp516 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maroni, G.; Panetta, D.; Luongo, R.; Krishnan, I.; La Rosa, F.; Campani, D.; Salvadori, P.; Iozzo, P.; Blasi, F.; Penkov, D.; et al. The Role of Prep1 in the Regulation of Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2019, 20, 3639. https://doi.org/10.3390/ijms20153639
Maroni G, Panetta D, Luongo R, Krishnan I, La Rosa F, Campani D, Salvadori P, Iozzo P, Blasi F, Penkov D, et al. The Role of Prep1 in the Regulation of Mesenchymal Stromal Cells. International Journal of Molecular Sciences. 2019; 20(15):3639. https://doi.org/10.3390/ijms20153639
Chicago/Turabian StyleMaroni, Giorgia, Daniele Panetta, Raffaele Luongo, Indira Krishnan, Federica La Rosa, Daniela Campani, Piero Salvadori, Patricia Iozzo, Francesco Blasi, Dmitry Penkov, and et al. 2019. "The Role of Prep1 in the Regulation of Mesenchymal Stromal Cells" International Journal of Molecular Sciences 20, no. 15: 3639. https://doi.org/10.3390/ijms20153639