2,4-Dinitrotoluene (DNT) Perturbs Yolk Absorption, Liver Development and Lipid Metabolism/Oxygen Transport Gene Expression in Zebrafish Embryos and Larvae

Abstract
:
1. Introduction
2. Results
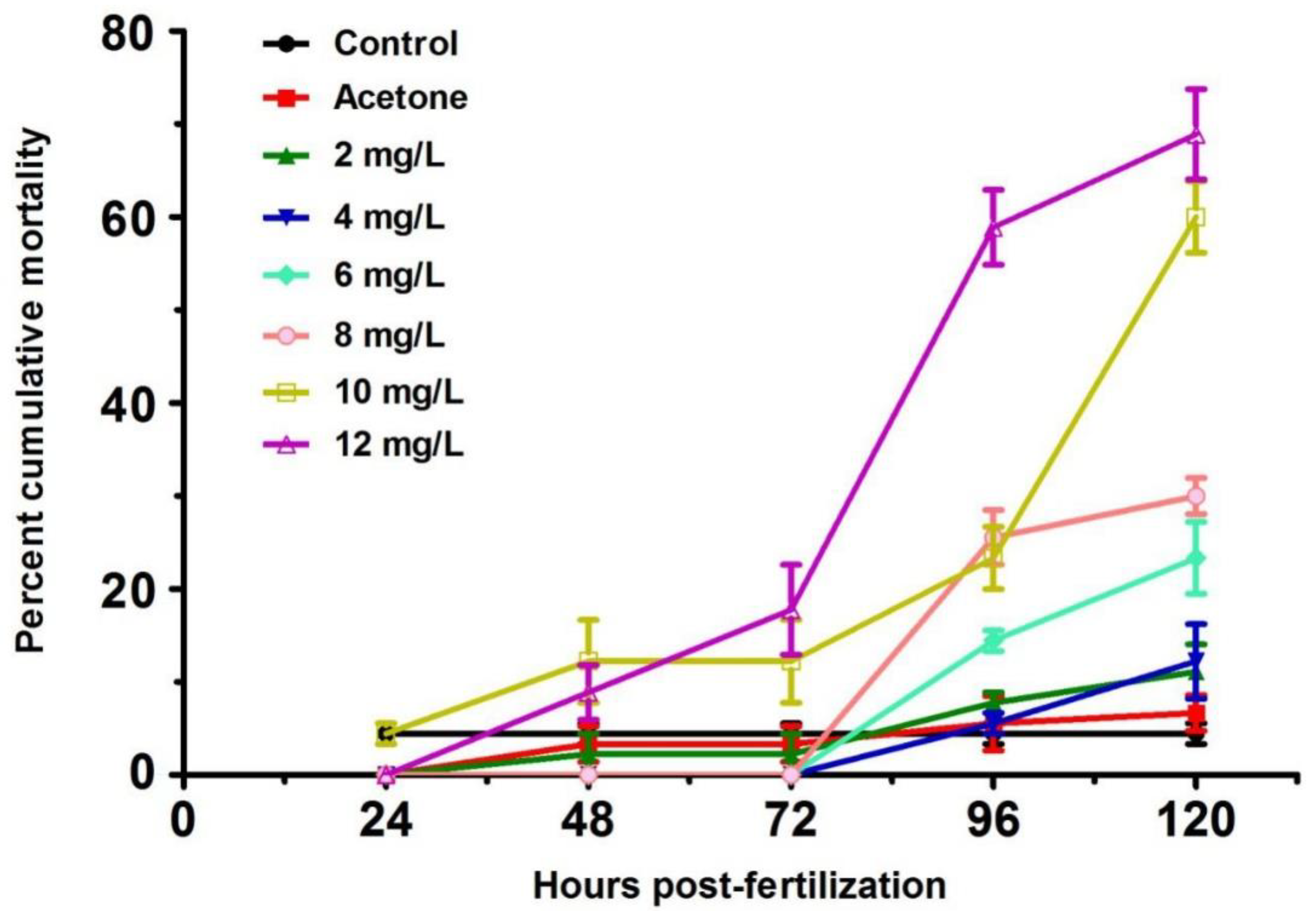
2.1. Mortality of Zebrafish Larvae from 24 to 120 h Post-Fertilization (hpf)
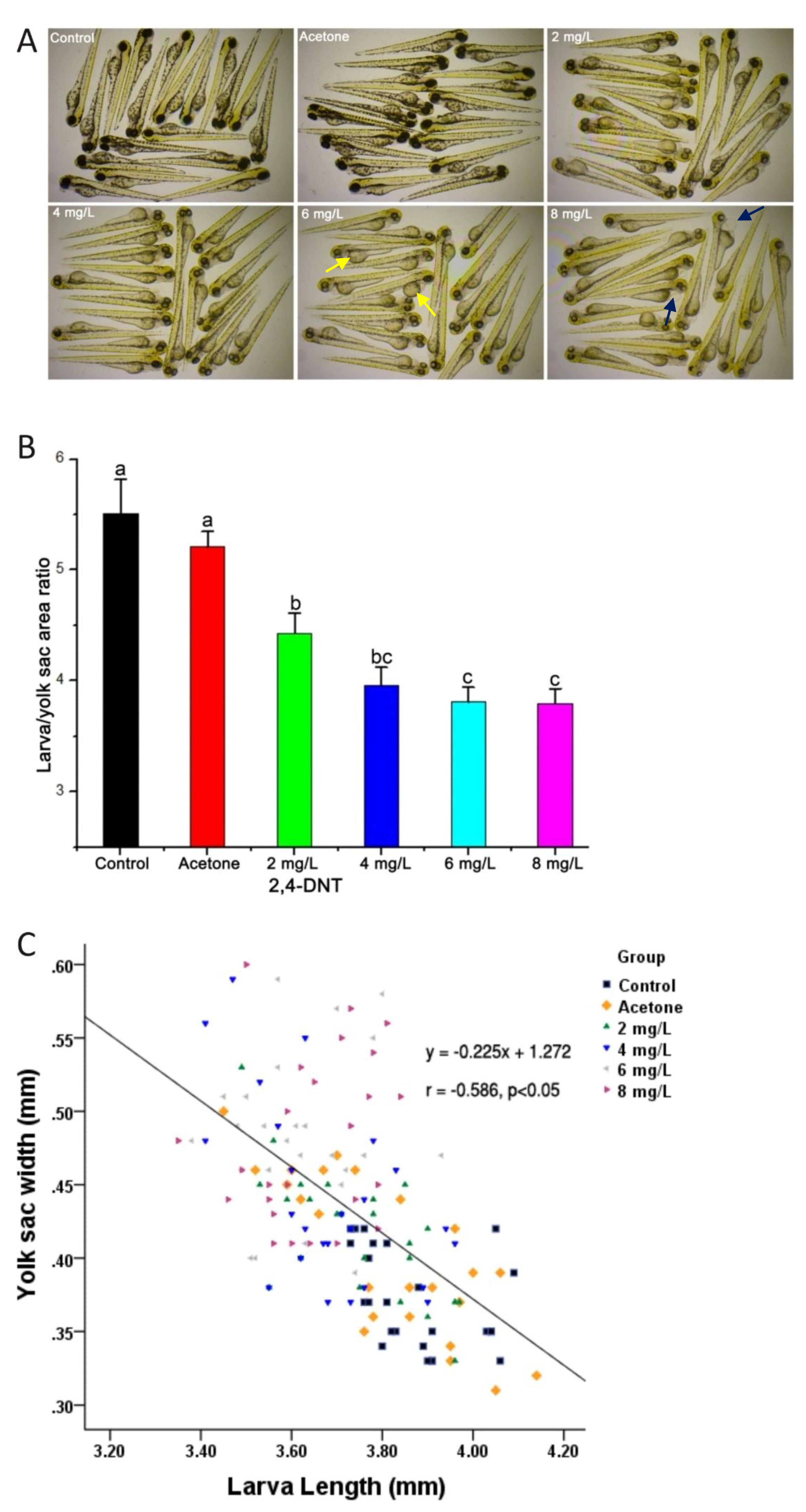
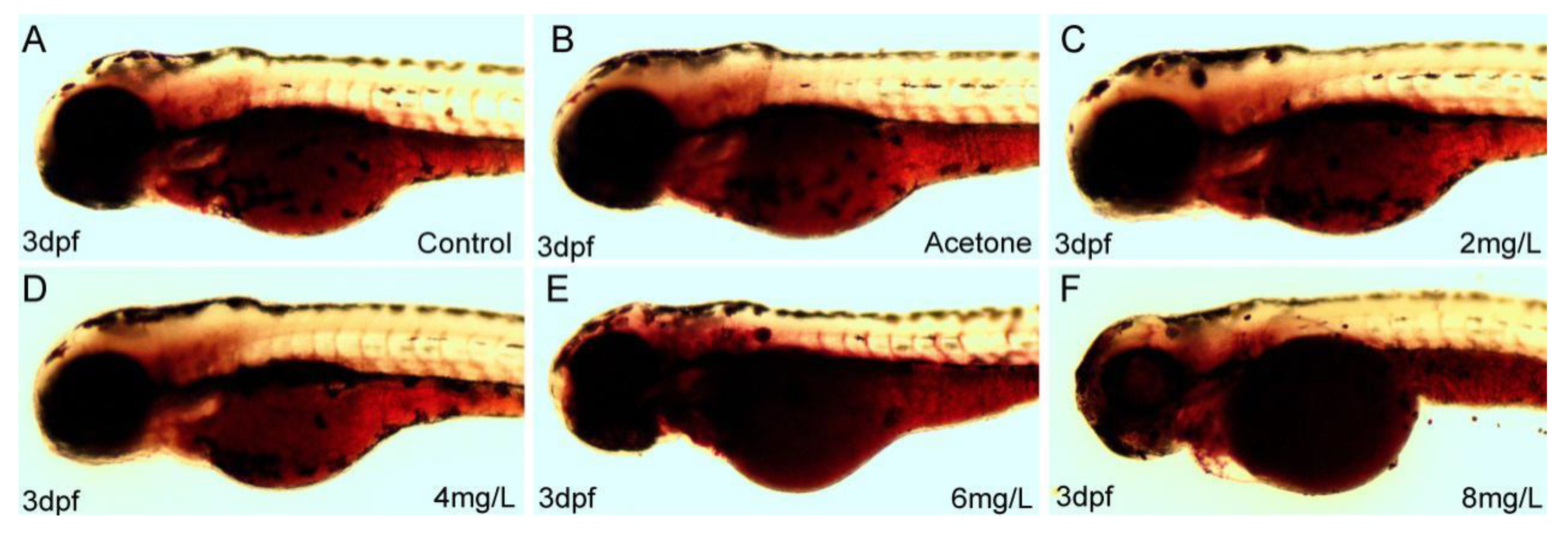
2.2. 2,4-DNT Induced Embryonic Malabsorption Syndrome at 3 Days Post-Fertilization (dpf)
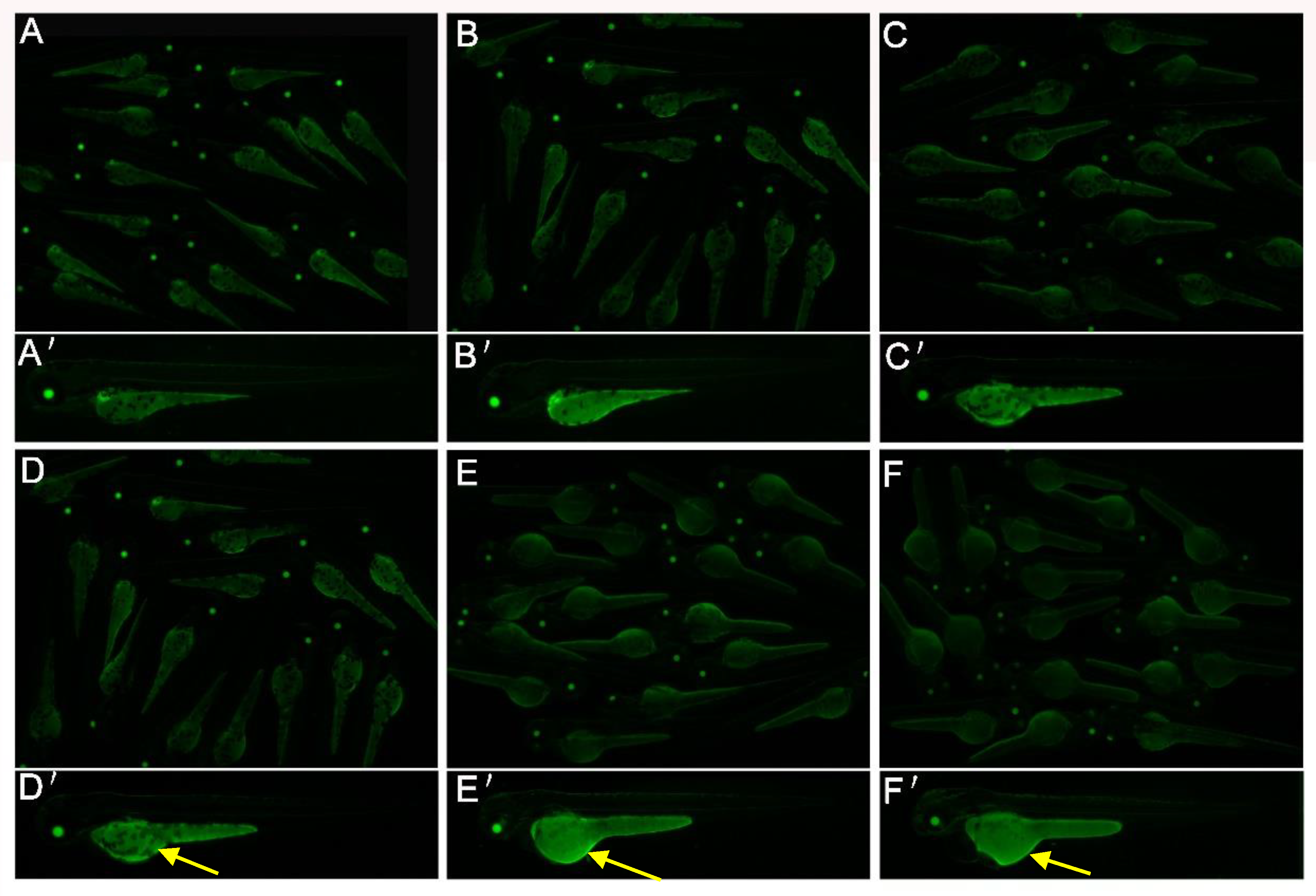
2.3. 2,4-DNT Impaired Endotrophic Lipid Consumption in Zebrafish Larvae at 3 dpf
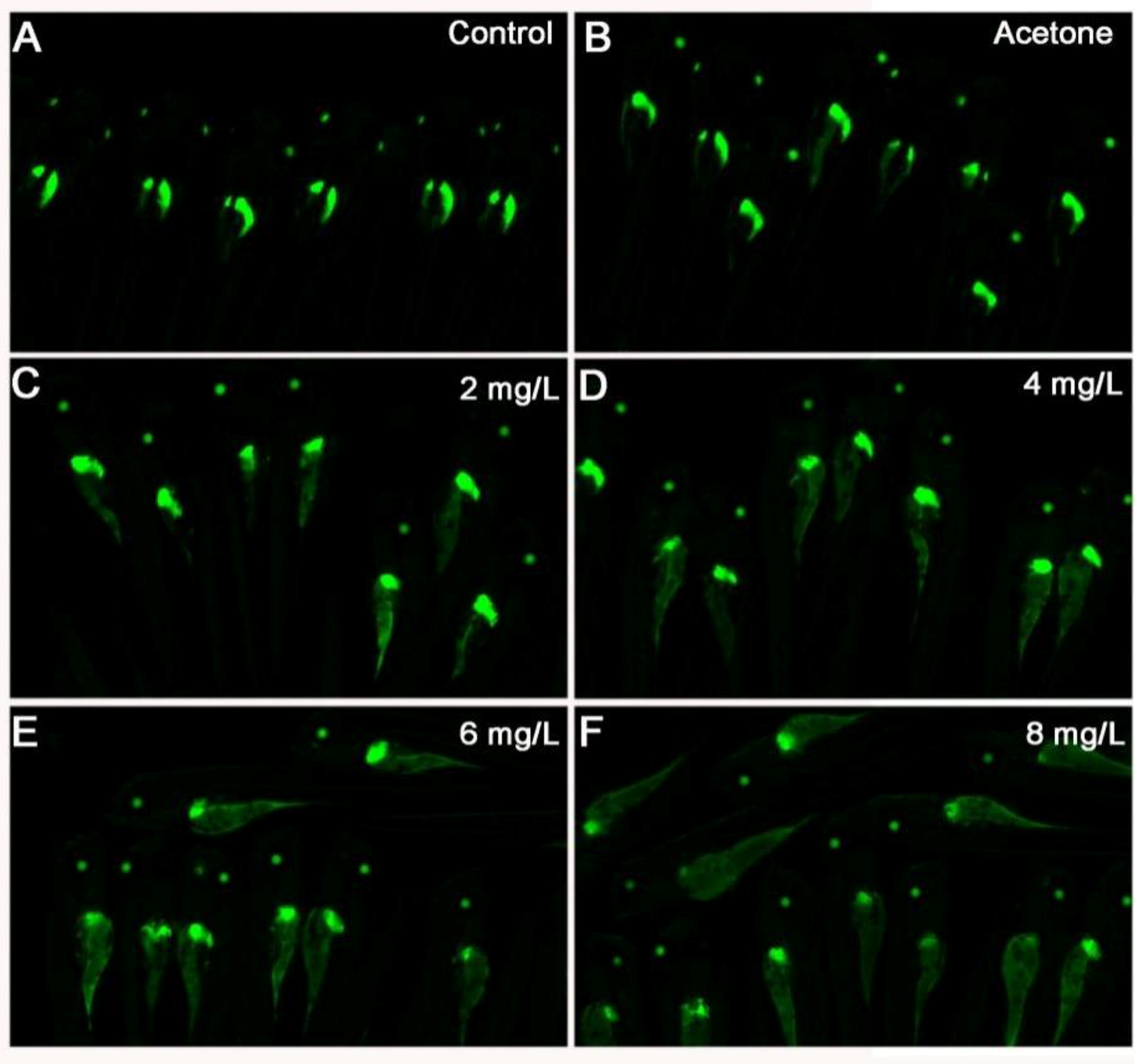
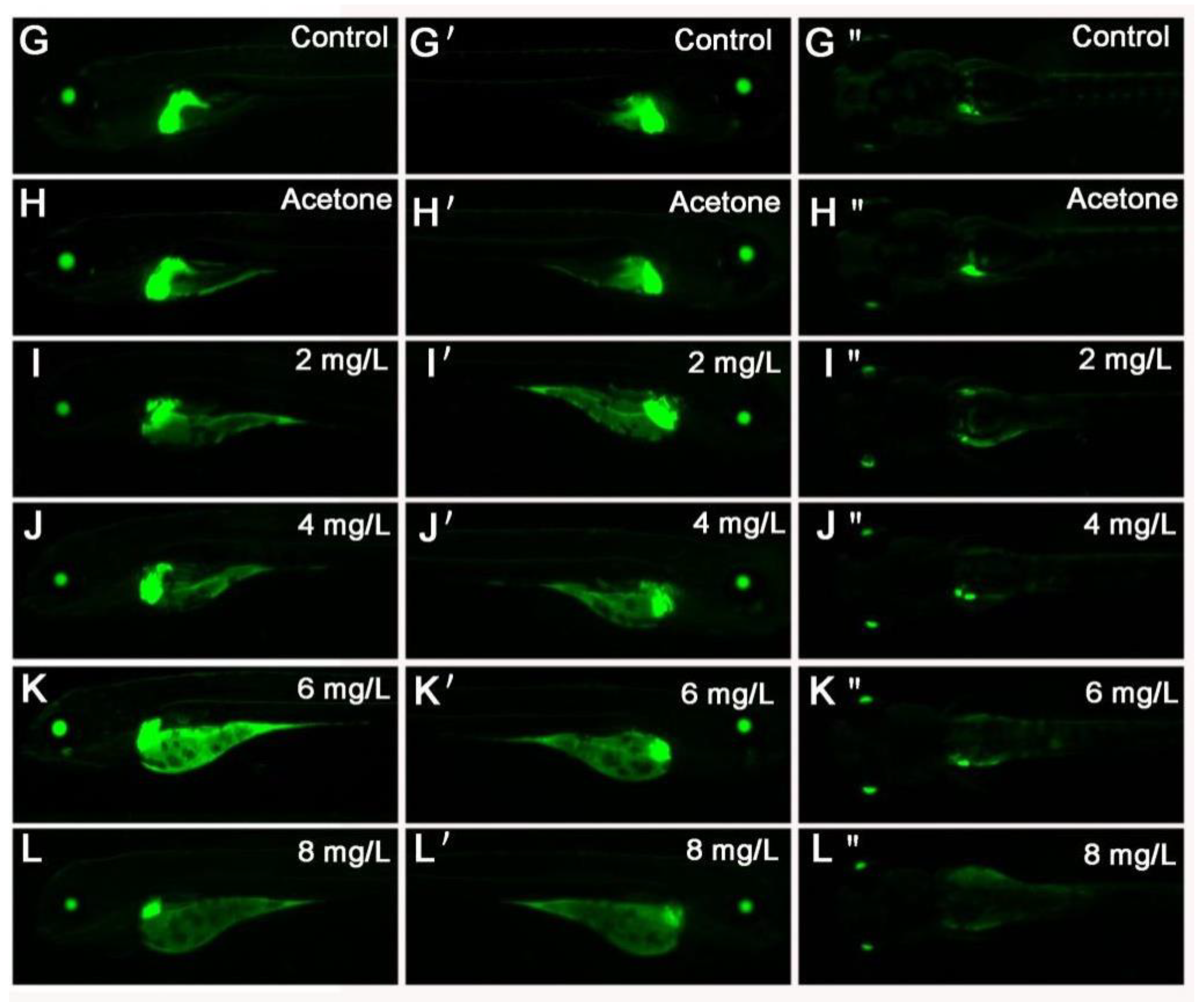
2.4. 2,4-DNT Inhibited Liver Organogenesis in 5 dpf Larvae
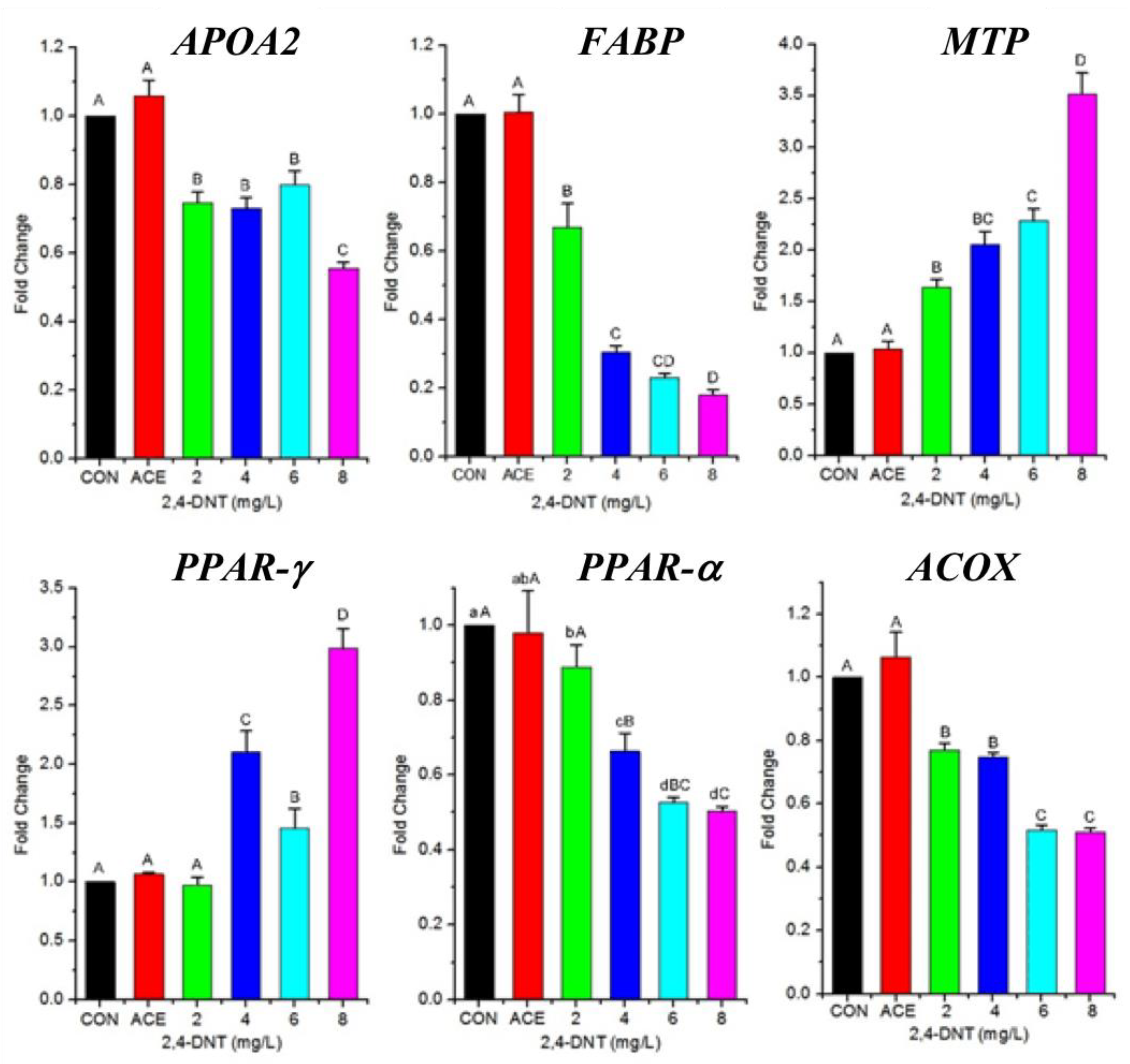
2.5. Effects of 2,4-DNT on the Expression of Genes Related to Lipid Transport and Lipolysis
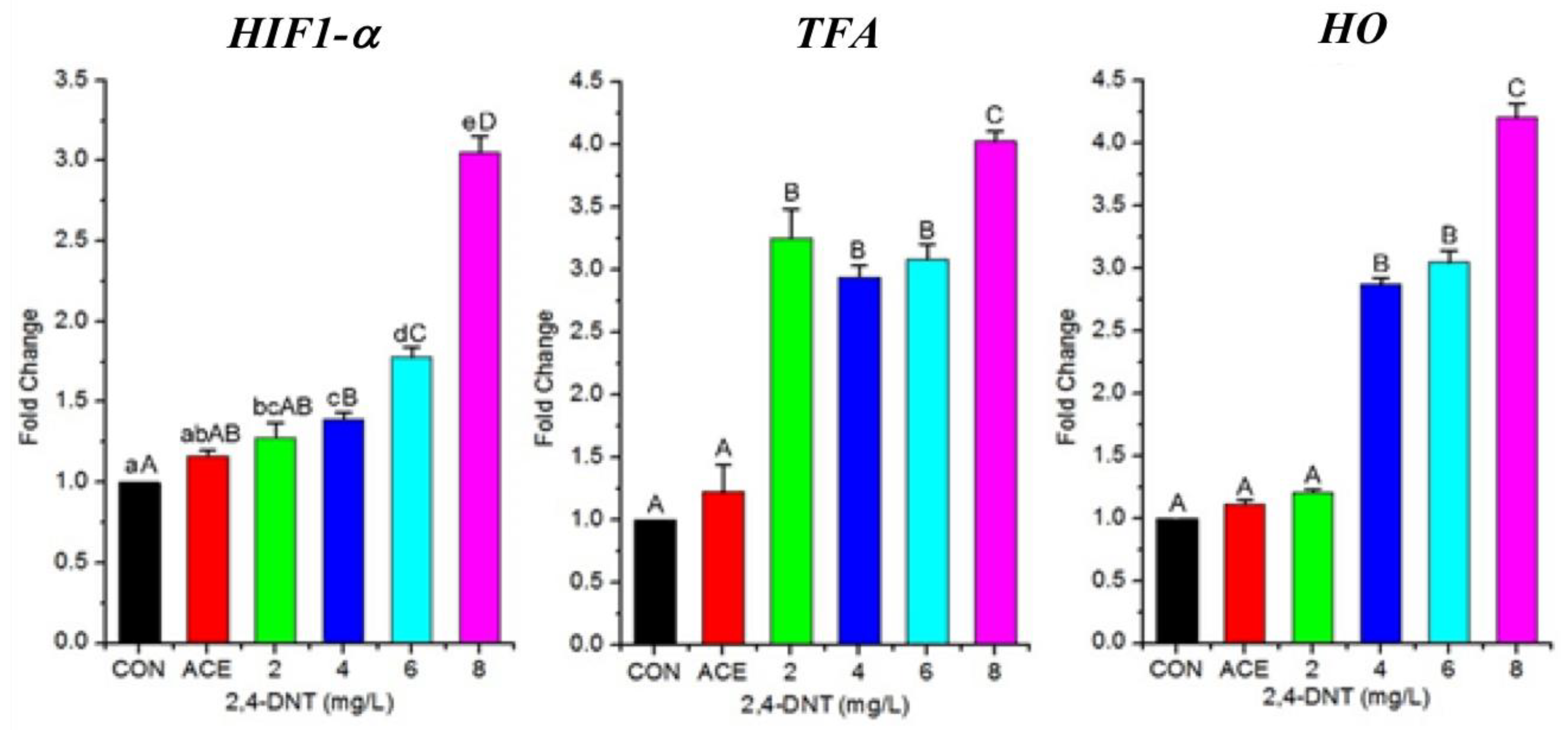
2.6. 2,4-DNT Up-Regulated Transcription of Genes Involved in Respiration
3. Discussion
4. Materials and Methods
4.1. Fish Breeding
4.2. Preparation of Chemicals and Solutions for Exposure Treatments
4.3. Exposure Procedures
4.4. Larvae Monitoring and Morphometric Analyses
4.5. ORO Staining
4.6. Isolation of Total Larval RNA and Quantitative RT-PCR
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tchounwou, P.B.; Wilson, B.A.; Ishaque, A.B.; Schneider, J. Transcriptional activation of stress genes and cytotoxicity in human liver carcinoma cells (hepg2) exposed to 2,4,6-trinitrotoluene, 2,4-dinitrotoluene, and 2,6-dinitrotoluene. Environ. Toxicol. 2001, 16, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Ishaque, A.B.; Timmons, C.; Ballard, F.V.; Hupke, C.; Dulal, K.; Johnson, L.R.; Gerald, T.M.; Boucaud, D.; Tchounwou, P.B. Cytotoxicity of dinitrotoluenes (2,4-DNT, 2,6-DNT) to MCF-7 and MRC-5 cells. Int. J. Environ. Res. Public Health 2005, 2, 304–307. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). List of Classifications. In IARC Monographs on the Evaluation of Carcinogenic Risk to Humans; IARC: Lyon, France, 1996; Available online: https://monographs.iarc.fr/agents-classified-by-the-iarc (accessed on 23 May 2019).
- U.S. Environmental Protection Agency (EPA). Drinking Water Health Advisory for 2,4-Dinitrotoluene and 2,6-Dinitrotoluene. In EPA Compliance Policy Guides; Document Number: 822-R-08-010; EPA: Washington, DC, USA, 2008. [Google Scholar]
- Shukla, N.; Gupta, V.; Rawat, A.S.; Gahlot, V.K.; Rai, P.K. 2,4-Dinitrotoluene (DNT) and 2,4,6-Trinitrotoluene (TNT) removal kinetics and degradation mechanism using zero valent iron-silica nanocomposite. J. Environ. Chem. Eng. 2018, 6, 5196–5203. [Google Scholar] [CrossRef]
- Jenkins, T.F.; Hewitt, A.D.; Grant, C.L.; Thiboutot, S.; Ampleman, G.; Walsh, M.E.; Ranney, T.A.; Ramsey, C.A.; Palazzo, A.J.; Pennington, J.C. Identity and distribution of residues of energetic compounds at army live-fire training ranges. Chemosphere 2006, 63, 1280–1290. [Google Scholar] [CrossRef] [Green Version]
- Lang, P.Z. Reaction and Tocixity of Organic Pollutants in the Songhua River; Jilin Science and Technology Press: Changchun, China, 1998. [Google Scholar]
- Pang, Z.Y.; Feng, L.; Zhou, J.L.; Liu, Z.T. Ecological risk assessment of substituted aromatic hydrocarbons in water of yangtze river estuary. Environ. Chem. 2011, 30, 430–434. [Google Scholar]
- Tchounwou, P.B.; Newsome, D.; Glass, K.; Centeno, J.A.; Lesczynski, J.; Bryant, J.; Okoh, J.; Ishaque, A.; Broker, M. Environmental toxicology and health effects associated with dinitrotoluene exposure. Rev. Environ. Health 2003, 18, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Jing, N. Effect of 2,4–dinitrotoluene exposure on enzyme activity, energy reserves and condition factors in common carp (Cyprinus carpio). J. Hazard. Mater. 2012, 203, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Lent, E.M. Chapter 6—Wildlife Toxicity Assessment for 2,4-Dinitrotoluene and 2,6-Dinitrotoluene. In Wildlife Toxicity Assessments for Chemicals of Military Concern; Williams, M.A., Reddy, G., Quinn, M.J., Jr., Johnson, M.S., Eds.; Elsevier Inc. Press: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Wintz, H.; Yoo, L.J.; Loguinov, A.; Wu, Y.Y.; Steevens, J.A.; Holland, R.D.; Beger, R.D.; Perkins, E.J.; Hughes, O.; Vulpe, C.D. Gene expression profiles in fathead minnow exposed to 2,4-DNT: Correlation with toxicity in mammals. Toxicol. Sci. 2006, 94, 71–82. [Google Scholar] [CrossRef]
- Poon, K.L.; Wang, X.G.; Lee, S.G.P.; Ng, A.S.; Goh, W.H.; Zhao, Z.H.; Al-Haddaw, M.; Wang, H.S.; Mathavan, S.; Ingham, P.W.; et al. Transgenic Zebrafish Reporter Lines as Alternative In Vivo Organ Toxicity Models. Toxicol. Sci. 2017, 156, 133–148. [Google Scholar]
- U.S. Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological Profile for 2,4- and 2,6-Dinitrotoluene; ATSDR: Atlanta, GA, USA, 1998.
- Bugel, S.M.; Tanguay, R.L.; Planchart, A. Zebrafish: A marvel of high-throughput biology for 21(st) century toxicology. Curr. Environ. Health Rep. 2014, 1, 341–352. [Google Scholar] [CrossRef]
- Paden, N.E.; Smith, E.E.; Maul, J.D.; Kendall, R.J. Effects of chronic 2,4,6,-trinitrotoluene, 2,4-dinitrotoluene, and 2,6-dinitrotoluene exposure on developing bullfrog (rana catesbeiana) tadpoles. Ecotox. Environ. Saf. 2011, 74, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Spanggord, R.J.; Suta, B.E. Effluent analysis of wastewater generated in the manufacture of 2,4,6-trinitrotoluene. Determination of a representative discharge of ether extractable components. Environ. Sci. Technol. 1982, 16, 233–236. [Google Scholar] [CrossRef]
- Escorcia, W.; Ruter, D.L.; Nhan, J.; Curran, S.P. Quantification of lipid abundance and evaluation of lipid distribution in caenorhabditis elegans by Nile Red and Oil Red O staining. J. Vis. Exp. 2018, 133, e57352. [Google Scholar] [CrossRef] [PubMed]
- Patiño, R.; Sullivan, C.V. Ovarian follicle growth, maturation, and ovulation in teleost fish. Fish Physiol. Biochem. 2002, 26, 57–70. [Google Scholar] [CrossRef]
- Lang, P.Z.; Wang, Y.; Chen, D.B.; Wang, N.; Ding, Y.Z. Bioconcentration, elimination and metabolism of 2,4-dinitrotoluene in carps (Cyprinus Carpio L.). Chemosphere 1997, 35, 1799–1815. [Google Scholar] [CrossRef]
- Deng, Y.; Meyer, S.A.; Guan, X.; Escalon, B.L.; Ai, J.; Wilbanks, M.S.; Welti, R.; Garcia-Reyero, N.; Perkins, E.J. Analysis of common and specific mechanisms of liver function affected by nitrotoluene compounds. PLoS ONE 2011, 6, e14662. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M.; Hennuyer, N.; Staels, B.; Fruchart, J.C.; Fievet, C.; Gonzalez, F.J.; Auwerx, J. Alterations in lipoprotein metabolism in peroxisome proliferator-activated receptor alpha-deficient mice. J. Biol. Chem. 1997, 272, 27307–27312. [Google Scholar] [CrossRef]
- Wilbanks, M.S.; Gust, K.A.; Atwa, S.; Sunesara, I.; Johnson, D.; Ang, C.Y.; Meyer, S.A.; Perkins, E.J. Validation of a genomics-based hypothetical adverse outcome pathway: 2,4-dinitrotoluene perturbs PPAR signaling thus impairing energy metabolism and exercise endurance. Toxicol. Sci. 2014, 141, 44–58. [Google Scholar] [CrossRef]
- Quinn, M.J.; Bazar, M.A.; Mcfarl, C.A.; Perkins, E.J.; Gust, K.A.; Gogal, R.M.; Johnson, M.S. Effects of subchronic exposure to 2,6-dinitrotoluene in the northern bobwhite (Colinus virginianus). Environ. Toxicol. Chem. 2007, 26, 2202–2207. [Google Scholar] [CrossRef]
- Kadegowda, A.K.; Bionaz, M.; Piperova, L.S.; Erdman, R.A.; Loor, J.J. Peroxisome proliferator-activated receptor-gamma activation and long-chain fatty acids alter lipogenic gene networks in bovine mammary epithelial cells to various extents. J. Dairy Sci. 2009, 92, 4276–4289. [Google Scholar] [CrossRef]
- Zhang, M.Q.; Gao, J.L.; Liao, X.D.; Huang, T.H.; Zhang, M.N.; Wang, M.Q.; Tian, Y.; Bai, J.; Zhou, C.H. miR-454 regulates triglyceride synthesis in bovine mammary epithelial cells by targeting PPAR-γ. Gene 2019, 691, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Hengbo, S.; Jun, L.; Jun, L.; Wangsheng, Z.; Huibin, T.; Huaiping, S. PPARG modulated lipid accumulation in dairy GMEC via regulation of ADRP gene. J. Cell. Biochem. 2015, 116, 192–201. [Google Scholar] [CrossRef]
- Jiang, C.; Ting, A.T.; Seed, B. PPAR-gamma agonists inhibit production of monocyte inflammatory cytokines. Nature 1998, 391, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, C.; Cai, H.; Xu, Y.; Lan, X.; Lei, C.; Chen, H. Novel polymorphisms of the APOA2 gene and its promoter region affect body traits in cattle. Gene 2013, 531, 288–293. [Google Scholar] [CrossRef] [PubMed]
- McKillop, I.H.; Girardi, C.A.; Thompson, K.J. Role of fatty acid binding proteins (FABPs) in cancer development and progression. Cell. Signal. 2019, 62, e109336. [Google Scholar] [CrossRef] [PubMed]
- Navasa, M.; Gordon, D.A.; Hariharan, N.; Jamil, H.; Shigenaga, J.K.; Moser, A.; Fiers, W.; Pollock, A.; Grunfeld, C.; Feingold, K.R. Regulation of microsomal triglyceride transfer protein mRNA expression by endotoxin and cytokines. J. Lipid Res. 1998, 39, 1220–1230. [Google Scholar]
- Lee, P.J.; Jiang, B.H.; Chin, B.Y.; Iyer, N.V.; Alam, J.; Semenza, G.L.; Choi, A.M.K. Hypoxia-inducible factor-1 mediates transcriptional activation of the heme oxygenase-1 gene in response to hypoxia. J. Biol. Chem. 1997, 272, 5375–5381. [Google Scholar] [CrossRef]
- Tacchini, L.; Bianchi, L.; Bernelli-Zazzera, A.; Cairo, G. Transferrin receptor induction by hypoxia. HIF-1-mediated transcriptional activation and cell-specific post-transcriptional regulation. J Biol Chem. 1999, 274, 24142–24146. [Google Scholar] [CrossRef]
- Rolfs, A.; Kvietikova, I.; Gassmann, M.; Wenger, R.H. Oxygen-regulated Transferrin Expression Is Mediated by Hypoxia-inducible Factor-1. J. Biol. Chem. 1997, 272, 20055–20062. [Google Scholar] [CrossRef] [Green Version]
- Semenza, G.L. Chairman’s Summary: Mechanisms of Oxygen Homeostasis, Circa 1999. In Oxygen Sensing Advances in Experimental Medicine and Biology; Lahiri, S., Prabhakar, N.R., Forster, R.E., Eds.; Springer Press: Boston, MA, USA, 2002. [Google Scholar]
- Beauchamp, R.O., Jr.; Irons, R.D.; Rickert, D.E.; Couch, D.B.; Hamm, T.E., Jr. A critical review of the literature on nitrobenzene toxicity. Crit. Rev. Toxicol. 1982, 11, 33–84. [Google Scholar] [CrossRef]
- Wang, R.; Li, Z.; Wang, Y.; Gui, J.F. An Apo-14 Promoter-Driven Transgenic Zebrafish That Marks Liver Organogenesis. PLoS ONE 2011, 6, e22555. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of zebrafish. Dev. Dynam. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Schlombs, K.; Wagner, T.; Scheel, J. Site-1 protease is required for cartilage development in zebrafish. Proc. Natl. Acad. Sci. USA 2003, 100, 14024–14029. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) |
|---|---|
| ZPPARGF | CGCAGGCTGAGAAGGAGAAGC |
| ZPPARGR | CATGTATCTGCAGTTGATCATC |
| ZPPARAF | CATCACCAGAGAGTTTCTGAAG |
| ZPPARAR | GCGGCGTTCACACTTATCGTAC |
| ZAOXF | AAGGACATCGAGCGAATGATG |
| ZAOXR | ACTATAAAAGAGTGGAGGCCG |
| APOF | ATGAAGCTGACATTCGCTCTC |
| APOR | TAGTGCTGGCTCAACTGCAG |
| ZFABPF | ATGGCCTTCAGCGGGACGTGG |
| ZFABPR | TGAGCTTCTTGCCGTCCATAG |
| ZMTPF | ATGAACATTTACGGTCAGAGC |
| ZMTPR | CACCACATTGATAGGATCTCC |
| ZHIF1AF | GTCAGCAAGAGCATGGGCCTC |
| ZHIF1AR | GAAGAACCTTCCACGTCGCAG |
| ZHOF | GCGGCAGAGAACACTGGCAGT |
| ZHOR | CTGCACTGCTGGGTGGTCTGC |
| ZTFAF | GAAGGTCCTGCTCATCTCTTTG |
| ZTFAR | CAGATAATTATTTAGTCCACCAG |
| ZHBAF | GAGTCTCTCTGCCAAAGACAAAG |
| ZHBAR | CGATTTTGCTGACAGCCTCAGC |
| ZACTINBF | CATGGATGAGGAAATCGCTGC |
| ZACTINBR | GTTAGTCACAATACCGTGCTC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, J.; Sha, H.; Zhou, H.; Peng, L.; Wu, L.; Qiu, Y.; Wang, R.; Hu, X. 2,4-Dinitrotoluene (DNT) Perturbs Yolk Absorption, Liver Development and Lipid Metabolism/Oxygen Transport Gene Expression in Zebrafish Embryos and Larvae. Int. J. Mol. Sci. 2019, 20, 3632. https://doi.org/10.3390/ijms20153632
Xiong J, Sha H, Zhou H, Peng L, Wu L, Qiu Y, Wang R, Hu X. 2,4-Dinitrotoluene (DNT) Perturbs Yolk Absorption, Liver Development and Lipid Metabolism/Oxygen Transport Gene Expression in Zebrafish Embryos and Larvae. International Journal of Molecular Sciences. 2019; 20(15):3632. https://doi.org/10.3390/ijms20153632
Chicago/Turabian StyleXiong, Jianglin, Hang Sha, Hualin Zhou, Lijuan Peng, Lingying Wu, Yinsheng Qiu, Rui Wang, and Xianqin Hu. 2019. "2,4-Dinitrotoluene (DNT) Perturbs Yolk Absorption, Liver Development and Lipid Metabolism/Oxygen Transport Gene Expression in Zebrafish Embryos and Larvae" International Journal of Molecular Sciences 20, no. 15: 3632. https://doi.org/10.3390/ijms20153632
APA StyleXiong, J., Sha, H., Zhou, H., Peng, L., Wu, L., Qiu, Y., Wang, R., & Hu, X. (2019). 2,4-Dinitrotoluene (DNT) Perturbs Yolk Absorption, Liver Development and Lipid Metabolism/Oxygen Transport Gene Expression in Zebrafish Embryos and Larvae. International Journal of Molecular Sciences, 20(15), 3632. https://doi.org/10.3390/ijms20153632