Ocular-Component-Specific miRNA Expression in a Murine Model of Lens-Induced Myopia

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
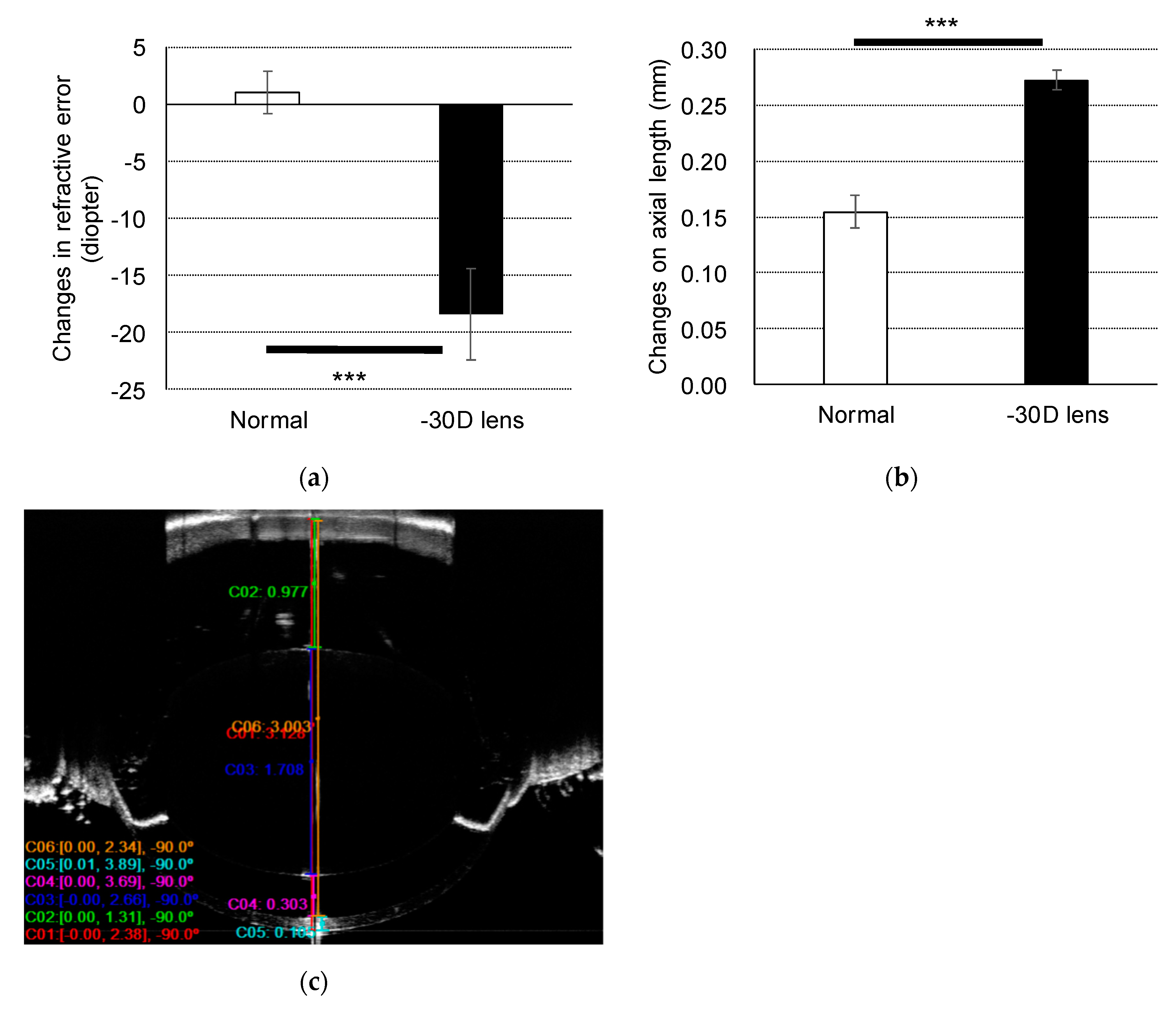
2.1. Refractive Error and Axial Length Changes in the Murine Model of Myopia
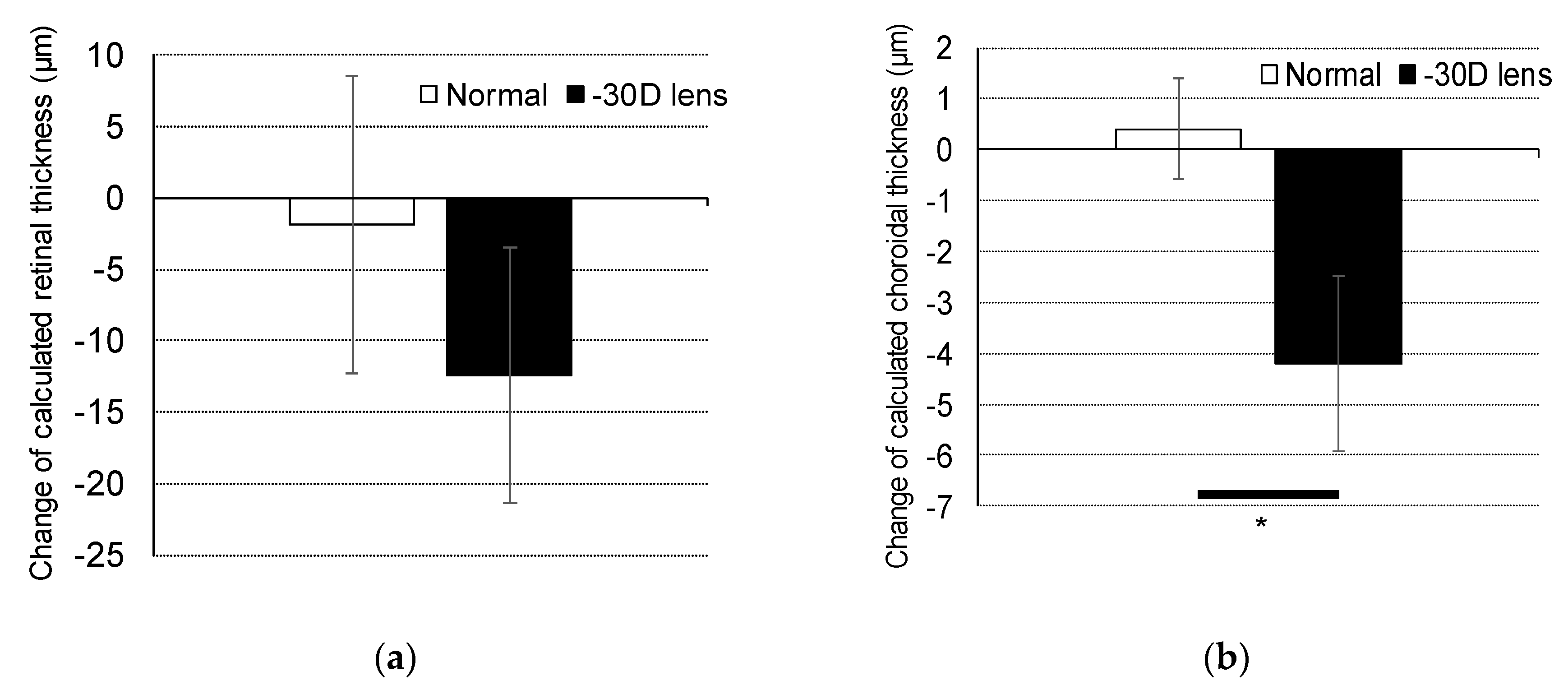
2.2. Change in Retinal and Choroidal Thickness
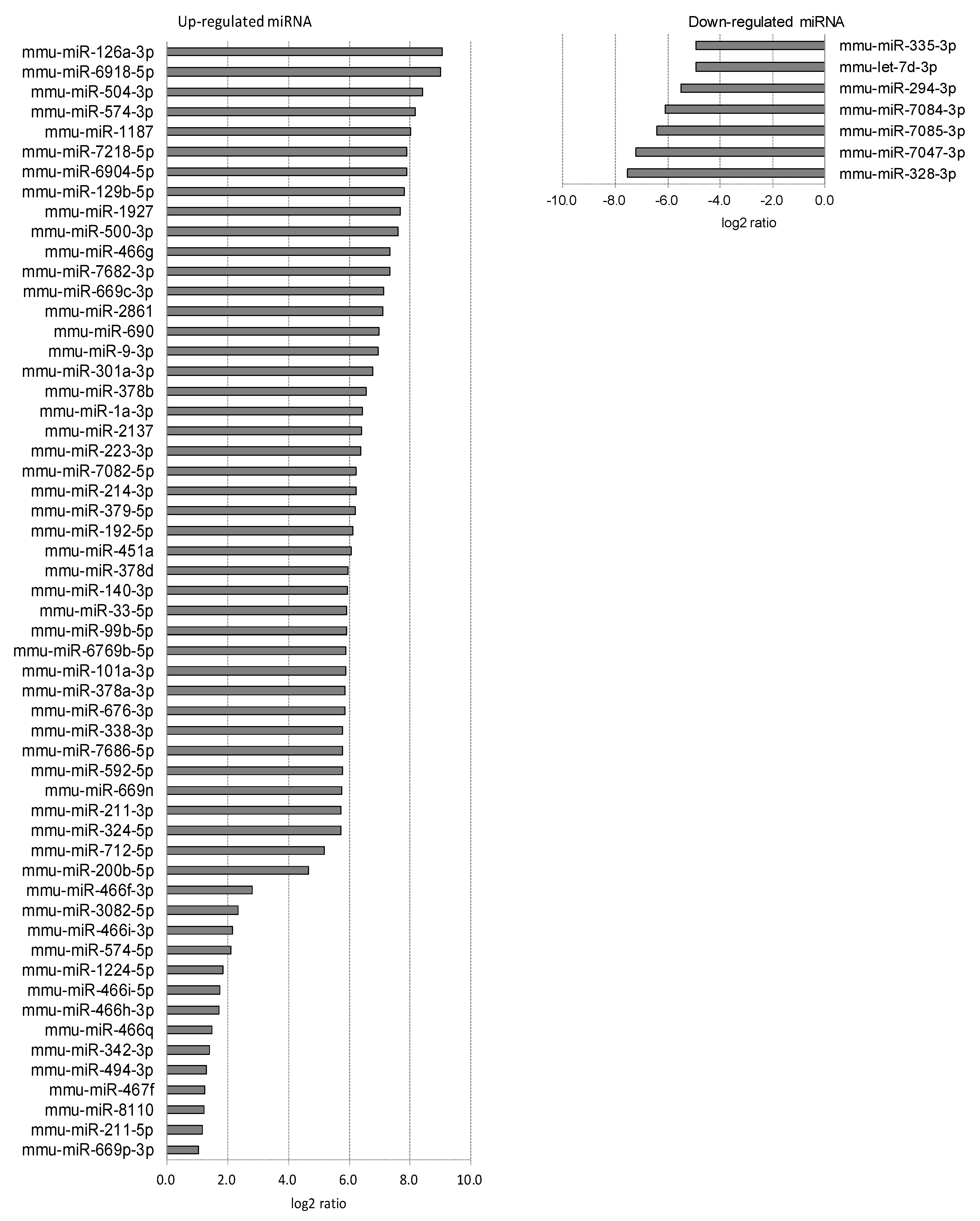
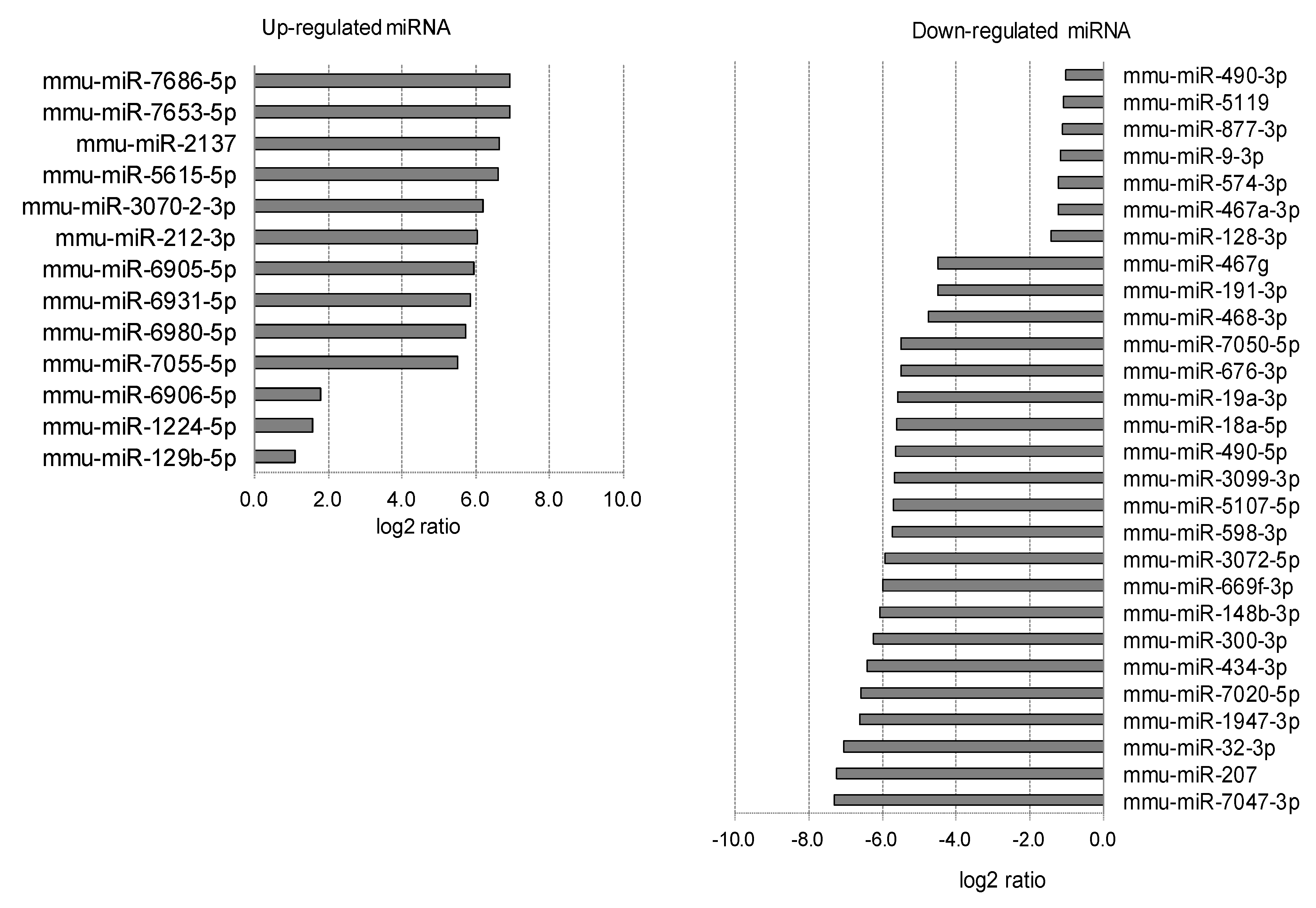
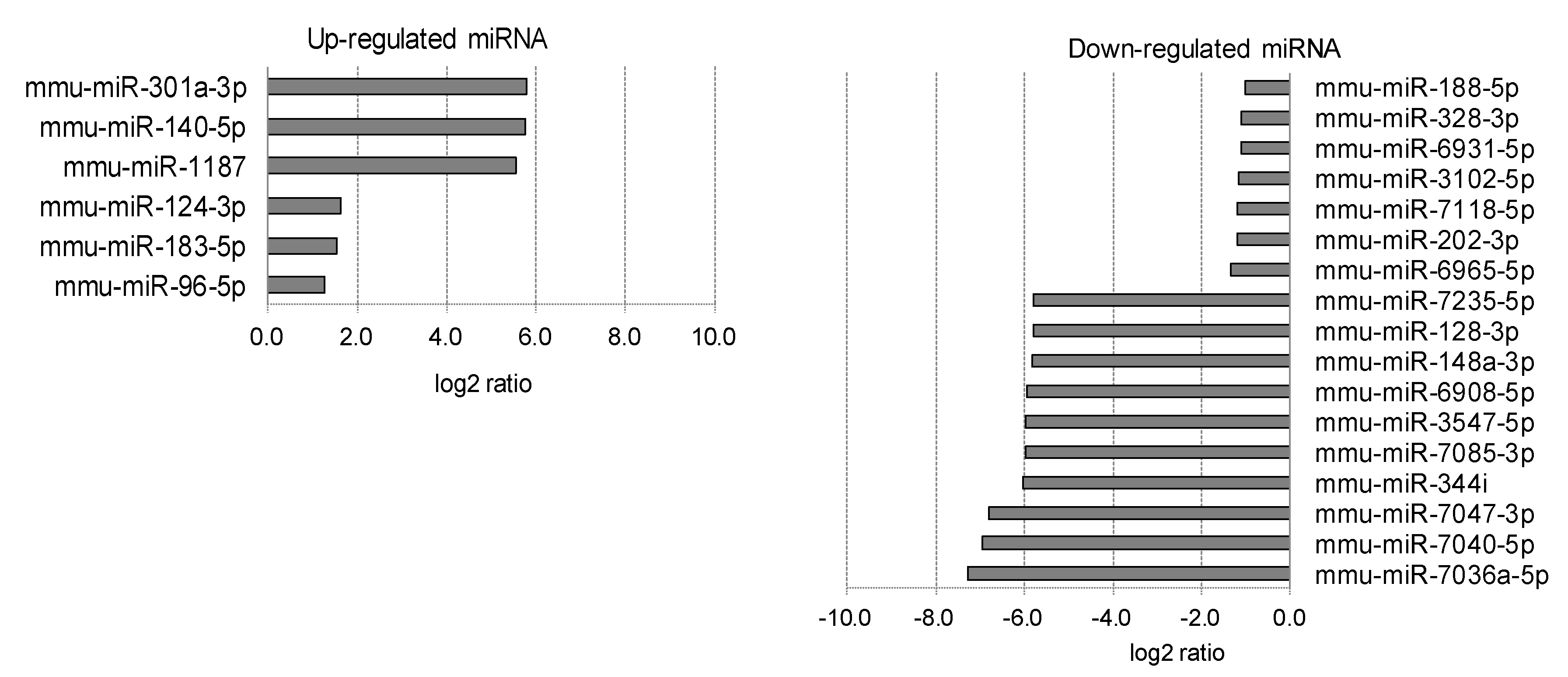
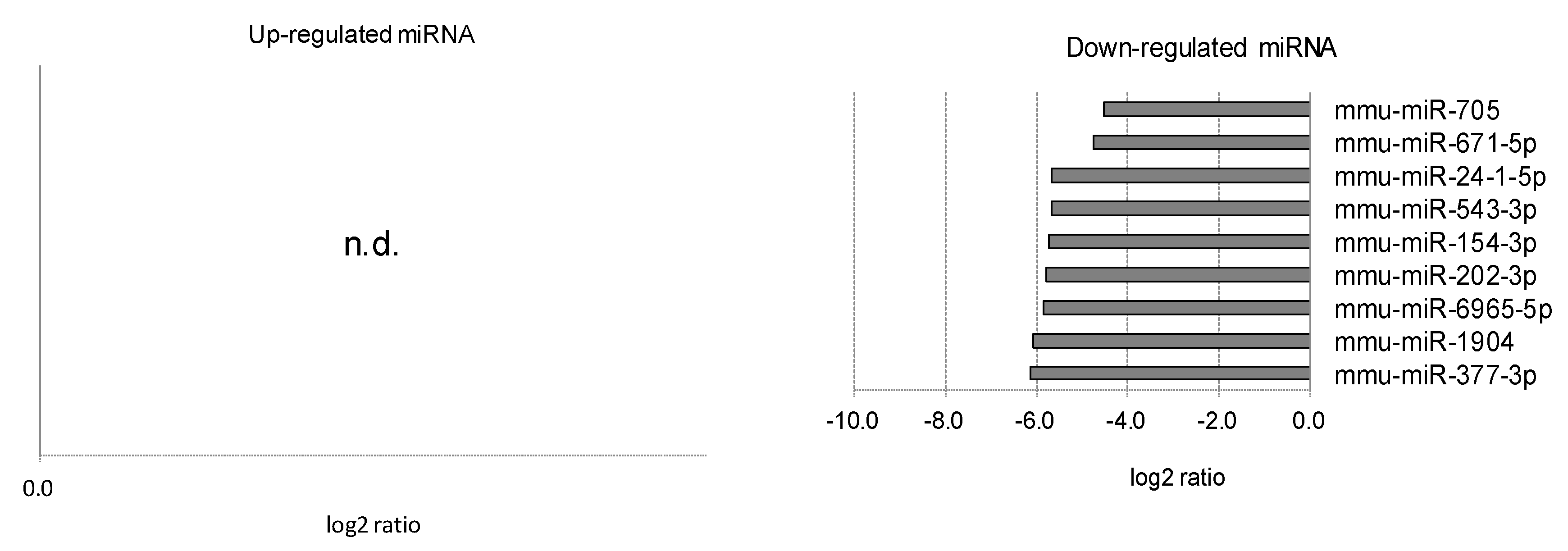
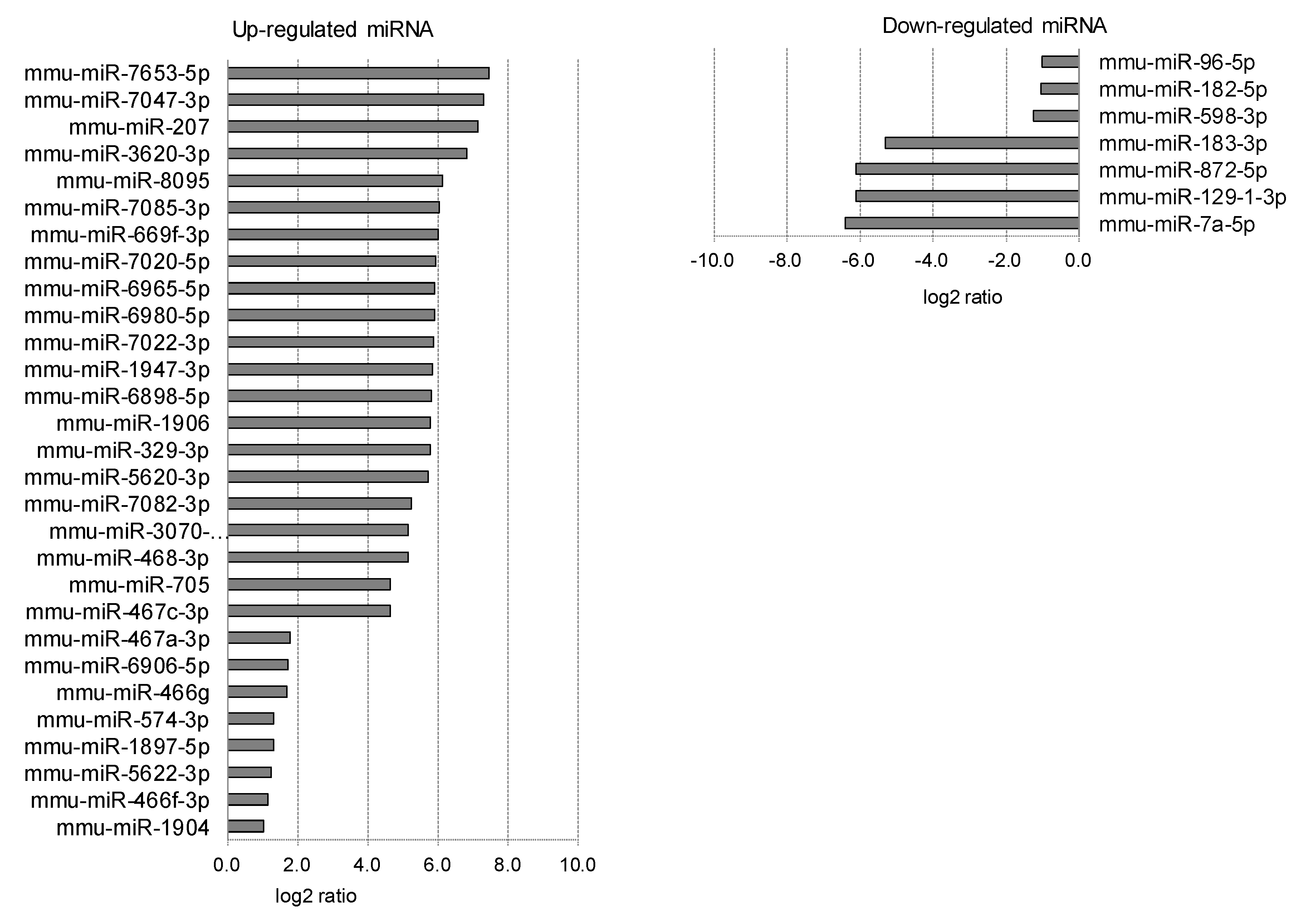
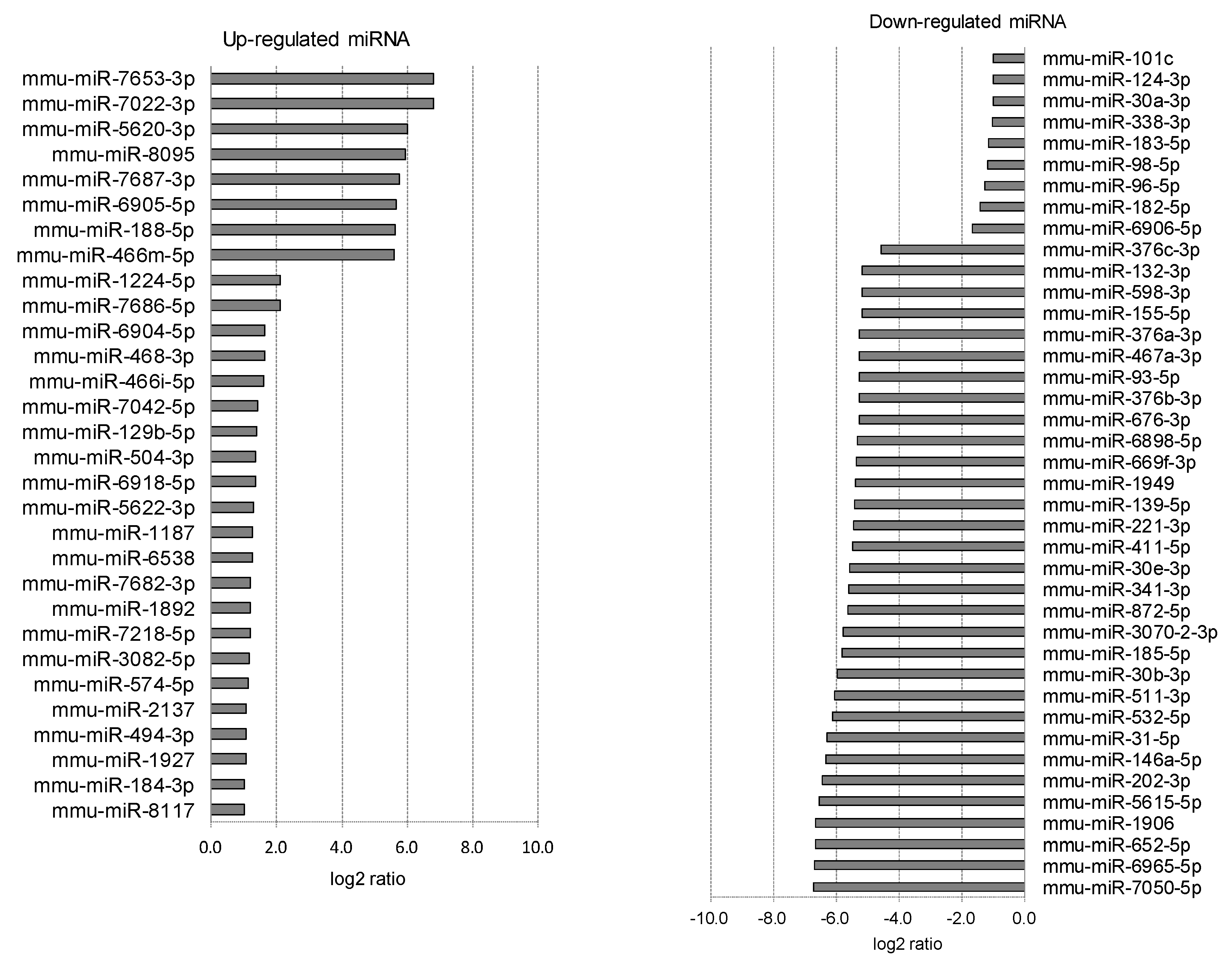
2.3. Differentially Expressed miRNAs in Individual Ocular Components During Myopia Progression
2.4. Overlapping Expression Changes in miRNAs in Different Ocular Component Tissues
2.5. Affected miRNAs and Their Predicted Target mRNA in Different Ocular Component Tissues
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Myopia Induction
4.3. miRNA Extraction
4.4. miRNA Microarray
4.5. Data Analysis
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global prevalence of myopia and high myopia and temporal trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. The myopia boom. Nature 2015, 519, 276–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, M.; Yamashiro, K.; Miyake, M.; Oishi, M.; Akagi-Kurashige, Y.; Kumagai, K.; Nakata, I.; Nakanishi, H.; Oishi, A.; Gotoh, N.; et al. Comprehensive replication of the relationship between myopia-related genes and refractive errors in a large Japanese cohort. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7343–7354. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, V.J.; Hysi, P.G.; Wojciechowski, R.; Fan, Q.; Guggenheim, J.A.; Höhn, R.; Mac Gregor, S.; Hewitt, A.W.; Nag, A.; Cheng, C.Y.; et al. Genome-wide meta-analyses of multi-ethnic cohorts identify multiple new susceptibility loci for refractive error and myopia jointly conceived the project and supervised the work NIH public access author manuscript. Veluchamy A. Barathi 2013, 29, 314–318. [Google Scholar]
- Jiang, X.; Kurihara, T.; Kunimi, H.; Miyauchi, M.; Ikeda, S.-I.; Mori, K.; Tsubota, K.; Torii, H.; Tsubota, K. A highly efficient murine model of experimental myopia. Sci. Rep. 2018, 8, 2026. [Google Scholar] [CrossRef] [PubMed]
- Ashby, R.S.; Schaeffel, F. The effect of bright light on lens compensation in Chicks. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5247–5253. [Google Scholar] [CrossRef] [PubMed]
- Ashby, R.; Kozulin, P.; Megaw, P.L.; Morgan, I.G. Alterations in ZENK and glucagon RNA transcript expression during increased ocular growth in chickens. Mol. Vis. 2010, 50, 639–649. [Google Scholar]
- Hui Xiao, Z.Y.F. Comparison of form-deprived myopia and lens-induced myopia in guinea pigs. Int J. Ophthalmol. 2014, 7, 245–250. [Google Scholar] [Green Version]
- Li, M.; Yuan, Y.; Chen, Q.; Me, R.; Gu, Q.; Yu, Y.; Sheng, M.; Ke, B. Expression of Wnt/β-catenin signaling pathway and its regulatory role in type I collagen with TGF-β1 in scleral fibroblasts from an experimentally induced myopia guinea pig model. J. Ophthalmol. 2016, 2016, 5126560. [Google Scholar] [CrossRef]
- He, L.; Frost, M.R.; Siegwart, J.T.; Norton, T.T. Gene expression signatures in tree shrew choroid in response to three myopiagenic conditions. Vision Res. 2014, 102, 52–63. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Liu, Q.; Ma, P.; Zhong, X.; Wu, J.; Ge, J. Effects of direct intravitreal dopamine injections on the development of lid-suture induced myopia in rabbits. Graefe’s Arch. Clin. Exp. Ophthalmol. 2006, 244, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hung, L.F.; Iii, E.L.S. Recovery of peripheral refractive errors and ocular shape in rhesus monkeys (Macaca mulatta) with experimentally induced myopia. Vis. Res. 2012, 73, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Riddell, N.; Faou, P.; Murphy, M.; Giummarra, L.; Downs, R.A.; Rajapaksha, H.; Crewther, S.G. The retina/RPE proteome in chick myopia and hyperopia models: Commonalities with inherited and age-related ocular pathologies. Mol. Vis. 2017, 23, 872–888. [Google Scholar] [PubMed]
- Metlapally, R.; Park, H.N.; Chakraborty, R.; Wang, K.K.; Tan, C.C.; Light, J.G.; Pardue, M.T.; Wildsoet, C.F. Genome-wide Scleral Micro-and messenger-RNA regulation during myopia development in the mouse. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6089–6097. [Google Scholar] [CrossRef] [PubMed]
- Tarver, J.E.; Donoghue, P.C.J.; Peterson, K.J. Do miRNAs have a deep evolutionary history? Bioessays 2012, 34, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Bhayani, M.K.; Calin, G.A.; Lai, S.Y. Functional relevance of miRNA * sequences in human disease. Mutat. Res. 2011, 731, 14–19. [Google Scholar] [CrossRef]
- Sethupathy, P. The promise and challenge of therapeutic MicroRNA silencing in diabetes and metabolic diseases. Curr. Diab. Rep. 2016, 16, 52. [Google Scholar] [CrossRef] [PubMed]
- Chim, S.S.C.; Shing, T.K.F.; Hung, E.C.W.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.K.; Lo, Y.M.D. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum microRNAs are promising novel biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef]
- Chen, K.C.; Hsi, E.; Hu, C.Y.; Chou, W.W.; Liang, C.L.; Juo, S.H.H. MicroRNA-328 may influence myopia development by mediating the PAX6 Gene. Investig. Opthalmol. Vis. Sci. 2012, 53, 2732. [Google Scholar] [CrossRef]
- Xie, M.; Li, Y.; Wu, J.; Wu, J. Genetic variants in MiR-29a associated with high myopia. Ophthalmic Genet. 2016, 37, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Hsi, E.; Chen, K.C.; Chang, W.S.; Yu, M.L.; Liang, C.L.; Juo, S.H.H. A Functional polymorphism at the FGF10 gene is associated with extreme myopia. Investig. Opthalmol. Vis. Sci. 2013, 54, 3265. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Zhang, Z.; Du, E.; Zheng, W.; Gu, Q.; Xu, X.; Ke, B. Wnt signaling in form deprivation myopia of the mice retina. PLoS ONE 2014, 9, e91086. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Mohamed, O.; Dufort, D.; Wallace, V.A. Characterization of Wnt signaling components and activation of the Wnt canonical pathway in the murine retina. Dev. Dyn. 2003, 227, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Schippert, R.; Burkhardt, E.; Feldkaemper, M.; Schaeffel, F. Relative axial myopia in Egr-1 (ZENK) knockout mice. Investig. Opthalmol. Vis. Sci. 2007, 48, 11. [Google Scholar] [CrossRef] [PubMed]
- Barathi, V.A.; Kwan, J.L.; Tan, Q.S.W.; Weon, S.R.; Seet, L.F.; Goh, L.K.; Vithana, E.N.; Beuerman, R.W. Muscarinic cholinergic receptor (M2) plays a crucial role in the development of myopia in mice. Dis. Model. Mech. 2013, 6, 1146–1158. [Google Scholar] [CrossRef] [PubMed]
- Cases, O.; Joseph, A.; Obry, A.; Santin, M.D.; Ben-Yacoub, S.; Pâques, M.; Amsellem-Levera, S.; Bribian, A.; Simonutti, M.; Augustin, S.; et al. Foxg1-Cre mediated Lrp2 inactivation in the developing mouse neural retina, ciliary and retinal pigment epithelia models congenital high myopia. PLoS ONE 2015, 10, e0129518. [Google Scholar] [CrossRef]
- Tkatchenko, A.V.; Tkatchenko, T.V.; Guggenheim, J.A.; Verhoeven, V.J.; Hysi, P.G.; Wojciechowski, R.; Singh, P.K.; Kumar, A.; Thinakaran, G.; Consortium for Refractive Error and Myopia (CREAM); et al. APLP2 regulates refractive error and myopia development in mice and humans. PLoS Genet. 2015, 11, e1005432. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, F.; Zhao, Y.; Sun, M.; Tao, J.; Liang, Y.; Ma, L.; Yu, Y.; Wang, J.; Hao, J. Enlargement of the axial length and altered ultrastructural features of the sclera in a mutant lumican transgenic mouse model. PLoS ONE 2016, 11, e0163165. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Schache, M.; Ikram, M.K.; Young, T.L.; Guggenheim, J.A.; Vitart, V.; Mac Gregor, S.; Verhoeven, V.J.M.; Barathi, V.A.; Liao, J.; et al. Nine loci for ocular axial length identified through genome-wide association studies, including shared loci with refractive error. Am. J. Hum. Genet. 2013, 93, 264–277. [Google Scholar] [CrossRef]
- Kito, N.; Endo, K.; Ikesue, M.; Weng, H.; Iwai, N. miRNA profiles of tubular cells: Diagnosis of kidney injury. Biomed. Res. Int. 2015, 2015, 465479. [Google Scholar] [CrossRef] [PubMed]
- Mei, F.; Wang, J.; Chen, Z.; Yuan, Z. Potentially important MicroRNAs in form-deprivation myopia revealed by bioinformatics analysis of MicroRNA profiling. Ophthalmic Res. 2017, 57, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Tkatchenko, A.V.; Luo, X.; Tkatchenko, T.V.; Vaz, C.; Tanavde, V.M.; Maurer-Stroh, S.; Zauscher, S.; Gonzalez, P.; Young, T.L. Large-scale microRNA expression profiling identifies putative retinal miRNA-mRNA signaling pathways underlying form-deprivation myopia in mice. PLoS ONE 2016, 11, e0162541. [Google Scholar] [CrossRef] [PubMed]
- Zorc, M.; Obsteter, J.; Dovc, P.; Kunej, T. Genetic variability of MicroRNA genes in 15 animal species. J. Genom. 2015, 3, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Mori, K.; Kurihara, T.; Miyauchi, M.; Ishida, A.; Jiang, X.; Ikeda, S.I.; Torii, H.; Tsubota, K. Oral crocetin administration suppressed refractive shift and axial elongation in a murine model of lens-induced myopia. Sci. Rep. 2019, 9, 295. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Combination | 2 Times Over or 0.5 Times Less *1 | 5 Times Over *2 or 0.2 Times Less *3 | Total | ||||
|---|---|---|---|---|---|---|---|
| 1 | Cornea | Numerator | Lens-induced | Up | 14 | 42 | 56 |
| Denominator | Normal | Down | 0 | 7 | 7 | ||
| 2 | Iris | Numerator | Lens-induced | Up | 3 | 10 | 13 |
| Denominator | Normal | Down | 7 | 21 | 28 | ||
| 3 | Lens | Numerator | Lens-induced | Up | 3 | 3 | 6 |
| Denominator | Normal | Down | 7 | 10 | 17 | ||
| 4 | Retina | Numerator | Lens-induced | Up | 0 | 0 | 0 |
| Denominator | Normal | Down | 0 | 9 | 9 | ||
| 5 | RPE/choroid | Numerator | Lens-induced | Up | 8 | 21 | 29 |
| Denominator | Normal | Down | 3 | 4 | 7 | ||
| 6 | Sclera | Numerator | Lens-induced | Up | 22 | 8 | 30 |
| Denominator | Normal | Down | 9 | 31 | 40 | ||
| Cornea | |||
|---|---|---|---|
| Up Each Eye-Tissues | Down Each Eye-Tissues | ||
| up | down | up | down |
| Iris | Iris | ||
| mmu-miR-7686-5p | mmu-miR-676-3p | - | mmu-miR-7047-3p |
| mmu-miR-2137 | mmu-miR-574-3p | Lens | |
| mmu-miR-1224-5p | mmu-miR-9-3p | - | mmu-miR-7047-3p |
| mmu-miR-129b-5p | mmu-miR-7085-3p | ||
| Lens | mmu-miR-328-3p | ||
| mmu-miR-301a-3p | - | Retina | |
| mmu-miR-1187 | - | - | |
| Retina | RPE/choroid | ||
| - | - | mmu-miR-7085-3p | - |
| RPE/choroid | mmu-miR-7047-3p | ||
| mmu-miR-466f-3p | - | Sclera | |
| mmu-miR-574-3p | - | - | |
| mmu-miR-466g | |||
| Sclera | |||
| mmu-miR-1927 | mmu-miR-676-3p mmu-miR-338-3p | ||
| mmu-miR-494-3p | |||
| mmu-miR-2137 | |||
| mmu-miR-574-5p | |||
| mmu-miR-3082-5p | |||
| mmu-miR-7218-5p | |||
| mmu-miR-7682-3p | |||
| mmu-miR-1187 | |||
| mmu-miR-6918-5p | |||
| mmu-miR-504-3p | |||
| mmu-miR-129b-5p | |||
| mmu-miR-466i-5p | |||
| mmu-miR-6904-5p | |||
| mmu-miR-7686-5p | |||
| mmu-miR-1224-5p | |||
| Iris | |||
|---|---|---|---|
| Up Each Eye-Tissues | Down Each Eye-Tissues | ||
| up | down | up | down |
| Lens | Lens | ||
| mmu-miR-6931-5p | - | mmu-miR-128-3p | |
| Retina | Retina | ||
| - | - | - | - |
| RPE/choroid | RPE/choroid | ||
| mmu-miR-7653-5p | - | mmu-miR-207 | mmu-miR-598-3p |
| mmu-miR-3070-2-3p | mmu-miR-1947-3p | ||
| mmu-miR-6980-5p | mmu-miR-7020-5p | ||
| mmu-miR-6906-5p | mmu-miR-669f-3p | ||
| Sclera | mmu-miR-468-3p | ||
| mmu-miR-6905-5p | mmu-miR-5615-5p | mmu-miR-467a-3p | |
| mmu-miR-3070-2-3p | Sclera | ||
| mmu-miR-6906-5p | mmu-miR-468-3p | mmu-miR-598-3p | |
| mmu-miR-7050-5p | |||
| mmu-miR-467a-3p | |||
| mmu-miR-669f-3p | |||
| Lens | |||
|---|---|---|---|
| Up Each Eye-Tissues | Down Each Eye-Tissues | ||
| up | down | up | down |
| Retina | Retina | ||
| - | - | - | mmu-miR-6965-5p |
| RPE/choroid | mmu-miR-202-3p | ||
| mmu-miR-96-5p | - | RPE/choroid | |
| Sclera | mmu-miR-6965-5p | - | |
| - | mmu-miR-96-5p | Sclera | |
| mmu-miR-183-5p | mmu-miR-188-5p | mmu-miR-6965-5p | |
| mmu-miR-124-3p | mmu-miR-202-3p | ||
| Retina | |||
|---|---|---|---|
| Up Each Eye-Tissues | DownEach Eye-Tissues | ||
| up | down | up | down |
| RPE/choroid | RPE/choroid | ||
| - | - | mmu-miR-1904 | - |
| Sclera | mmu-miR-705 | ||
| - | - | Sclera | |
| - | - | ||
| RPE/Choroid | |||
|---|---|---|---|
| Up Each Eye-Tissues | Down Each Eye-Tissues | ||
| up | down | up | down |
| Sclera | Sclera | ||
| mmu-miR-5622-3p | mmu-miR-1906 mmu-miR-6898-5p | - | mmu-miR-872-5p |
| mmu-miR-8095 | mmu-miR-182-5p | ||
| mmu-miR-5620-3p | |||
| mmu-miR-7022-3p | |||
| miRNAs | Overlapping Tissues | |||
|---|---|---|---|---|
| mmu-miR-1187 | Cornea | Lens | Sclera | |
| mmu-miR-1224-5p | Cornea | Iris | Sclera | |
| mmu-miR-129b-5p | Cornea | Iris | Sclera | |
| mmu-miR-202-3p | Lens | Retina | Sclera | |
| mmu-miR-2137 | Cornea | Iris | Sclera | |
| mmu-miR-3070-2-3p | Iris | RPE/choroid | Sclera | |
| mmu-miR-467a-3p | Iris | RPE/choroid | Sclera | |
| mmu-miR-468-3p | Iris | RPE/choroid | Sclera | |
| mmu-miR-574-3p | Cornea | Iris | RPE/choroid | |
| mmu-miR-598-3p | Iris | RPE/choroid | Sclera | |
| mmu-miR-669f-3p | Iris | RPE/choroid | Sclera | |
| mmu-miR-676-3p | Cornea | Iris | Sclera | |
| mmu-miR-6906-5p | Iris | RPE/choroid | Sclera | |
| mmu-miR-6965-5p | Lens | RPE/choroid | Sclera | |
| mmu-miR-7047-3p | Cornea | Iris | Lens | RPE/choroid |
| mmu-miR-7085-3p | Cornea | Lens | RPE/choroid | |
| mmu-miR-7686-5p | Cornea | Iris | Sclera | |
| mmu-miR-96-5p | Lens | RPE/choroid | Sclera | |
| Cornea | |||
|---|---|---|---|
| Upregulated miRNAs | Target gene | Downregulated miRNAs | Target gene |
| mmu-miR-211-5p | PPM1K | mmu-miR-328-3p | BACE1 |
| mmu-miR-494-3p | FGFR2, ROCK1 | mmu-miR-294-3p | CDKN1A |
| mmu-miR-200b-5p | ARHGDIA | ||
| mmu-miR-669n | SENP6 | ||
| mmu-miR-338-3p | FGFR2, Runx2, TNFSF11 | ||
| mmu-miR-378a-3p | Nrf1, GLI3, MAPK1 | ||
| mmu-miR-101a-3p | COX2, DUSP1, ACKR3, NeuroD1 | ||
| mmu-miR-99b-5p | MFGE8 | ||
| mmu-miR-33-5p | ABCG1, ABCA1, NRIP1, CCL2, HMGA2 | ||
| mmu-miR-451a | MYC, YWHAZ, ANKRD46 | ||
| mmu-miR-192-5p | H3F3BH | ||
| mmu-miR-214-3p | POU4F2, SP7, DIO3, Atg12, PTEN | ||
| mmu-miR-223-3p | MEF2C, STAT3, GRIA2, GRIN2B, FOXO3, IGF1R, LIF, IL6 | ||
| mmu-miR-1a-3p | CALM2, CALM1, MEF2A, IGF1, HAND2, IRX5, GJA1, KLF4, CDC42, TLX2, ANXA5, MKL1, SMARCD2, CLCN3, SMARCB1, MAP4K3, FZD7, MEOX2, NFAT5, RARB, SH3BGR1, MYOCD, ADAR, BDNF | ||
| mmu-miR-301a-3p | PIAS3, SOCS5, IRF1, PTEN | ||
| mmu-miR-9-3p | SCN2B | ||
| mmu-miR-690 | VCAN, CTNNB1 | ||
| mmu-miR-126a-3p | PIK3R2, IRS1, ITGA11, ERRFI1, FZD7 | ||
| Iris | |||
|---|---|---|---|
| Upregulated miRNAs | Target gene | Downregulated miRNAs | Target gene |
| mmu-miR-212-3p | MMP9, HMGB1 | mmu-miR-434-3p | VCAN, CTNNB1, EIF5A |
| mmu-miR-148b-3p | CAMK2A, DNMT1 | ||
| mmu-miR-18a-5p | HSF2, HIF1A | ||
| mmu-miR-19a-3p | ZFPM2, PTEN, TNF, FZD4, LRP6 | ||
| mmu-miR-468-3p | HELLS | ||
| mmu-miR-467g | RUNX2 | ||
| mmu-miR-128-3p | POU4F2, NF1, PPARA, RUNX1, PAX3, ABHD5, MAPK14, DCX | ||
| mmu-miR-9-3p | SCN2B | ||
| mmu-miR-877-3p | SMAD7 | ||
| mmu-miR-490-3p | NANOG | ||
| Lens | |||
|---|---|---|---|
| Upregulated miRNAs | Target gene | Downregulated miRNAs | Target gene |
| mmu-miR-301a-3p | PIAS3, SOCS5, IRF1, PTEN | mmu-miR-148a-3p | CAMK2A, KDM6B, ROCK1, MET |
| mmu-miR-140-5p | HDAC4, ASP1, TGFBR1, WNT11, OSTM1 | mmu-miR-128-3p | POU4F2, NF1, PPARA, RUNX1, PAX3, ABHD5, MAPK14, DCX |
| mmu-miR-124-3p | ITGB1, FOXA2, SYCP1, PTBP1, CTDSP1 | mmu-miR-328-3p | BACE1 |
| mmu-miR-183-5p | ZEB2, LRP6 | ||
| mmu-miR-96-5p | CLIC5, INSIG2, AKT1S1 | ||
| RPE/choroid | |||
|---|---|---|---|
| Upregulated miRNAs | Target gene | Downregulated miRNAs | Target gene |
| mmu-miR-468-3p | HELLS | mmu-miR-7a-5p | HELLS, EIF4E, RPS6KB1, MAPKAP1, MKNK2, MKNK1, Sp1, PARP1, NLRP3, HERPUD2, MYRIP, PAX6, KLF4, PTK2 |
| mmu-miR-329-3p | DLK1 | ||
| mmu-miR-182-5p | CLIC5, FBXW7, MYOD1, TLR4 | ||
| mmu-miR-96-5p | CLIC5, INSIG2, AKT1S1 | ||
| Sclera | |||
|---|---|---|---|
| Upregulated miRNAs | Target gene | Downregulated miRNAs | Target gene |
| mmu-miR-184-3p | FZD4, NUMBL, SLC25A22 | mmu-miR-146a-5p | IRAK2, TRAF6, VSIVGP2, NOTCH1, MED1, RELB, IRAK1, MAP1B, RNF11, WNT1, WNT5A, EGR1, PPP3R2, TGIF1, CAMK2D |
| mmu-miR-494-3p | FGFR2, ROCK1 | ||
| mmu-miR-468-3p | HELLS | ||
| mmu-miR-31-5p | HIF1A, PDGFB, FZD4, TGFB2, VAV3 | ||
| mmu-miR-30b-3p | PPIF | ||
| mmu-miR-185-5p | KDM6B, VCAN, CTNNB1 | ||
| mmu-miR-30e-3p | SPATA19, NPFFR2, MRPS30 | ||
| mmu-miR-221-3p | DDIT4, KIT, ARNT | ||
| mmu-miR-139-5p | FOXO1, NOTCH1, IRS1 | ||
| mmu-miR-376b-3p | NFKBIZ, STAT3, HOXD10 | ||
| mmu-miR-93-5p | STAT3, VSIVGP2, SQSTM1, NFE2L2, BMPR2 | ||
| mmu-miR-376a-3p | PCNA | ||
| mmu-miR-155-5p | RHOA, RHEB, SOCS1, AICDA, PEA15A, MAF, SPI1, FGF7, INPP5D, FOS, PMAIP1, CLDN1, HDAC4, NR1H3, CISH, GSK3B, CSNK1A1 | ||
| mmu-miR-132-3p | MECP2, EP300, KDM5A, BTG2, PAIP2, MMP9, NR4A2, ARHGAP32, MAPT, SOX4 | ||
| mmu-miR-182-5p | CLIC5, FBXW7, MYOD1, TLR4 | ||
| mmu-miR-96-5p | CLIC5, INSIG2, AKT1S1 | ||
| mmu-miR-98-5p | ACVR1B, MMP11, IL6, WNT1 | ||
| mmu-miR-183-5p | ZEB2, LRP6 | ||
| mmu-miR-338-3p | FGFR2, RUNX2, TNFSF11 | ||
| mmu-miR-124-3p | ITGB1, FOXA2, SYCP1, PTBP1, PTBP2, CTDSP1, CEBPA, DLX2, SOX9, PTBP2, DLX5, CCNA2, NR3C2, PIM1, CAV1 | ||
| Overlapping Expression Changes of miRNAs in Two Tissues and Target Genes | |
|---|---|
| Upregulated in both cornea and lens | Target gene |
| mmu-miR-301a-3p | FGFR2, RUNX2, TNFSF11 |
| Upregulated in both cornea and sclera | |
| mmu-miR-494-3p | FGFR2, ROCK1 |
| Downregulated in both cornea and lens | |
| mmu-miR-328-3p | BACE1 |
| Upregulated in cornea and downregulated in iris | |
| mmu-miR-9-3p | SCN2B |
| Upregulated in cornea and downregulated in sclera | |
| mmu-miR-338-3p | PIAS3, SOCS5, IRF1, PTEN |
| Downregulated in both iris and lens | |
| mmu-miR-128-3p | POU4F2, NF1, PPARA, RUNX1, PAX3, ABHD5, MAPK14, DCX |
| Upregulated in lens and downregulated in sclera | |
| mmu-miR-124-3p | ITGB1, FOXA2, SYCP1, PTBP1, CTDSP1 |
| mmu-miR-183-5p | ZEB2, LRP6 |
| Downregulated in both RPE/choroid and sclera | |
| mmu-miR-182-5p | CLIC5, FBXW7, MYOD1, TLR4 |
| Overlapping Expression Changes of miRNAs in Three Tissues and Target Gene | |
|---|---|
| Upregulated in lens and RPE/choroid, downregulated in sclera | Target gene |
| mmu-miR-96-5p | CLIC5, INSIG2, AKT1S1 |
| Upregulated in iris, RPE/choroid and sclera | |
| mmu-miR-468-3p | HELLS |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, Y.; Kurihara, T.; Hagiwara, Y.; Ikeda, S.-i.; Mori, K.; Jiang, X.; Torii, H.; Tsubota, K. Ocular-Component-Specific miRNA Expression in a Murine Model of Lens-Induced Myopia. Int. J. Mol. Sci. 2019, 20, 3629. https://doi.org/10.3390/ijms20153629
Tanaka Y, Kurihara T, Hagiwara Y, Ikeda S-i, Mori K, Jiang X, Torii H, Tsubota K. Ocular-Component-Specific miRNA Expression in a Murine Model of Lens-Induced Myopia. International Journal of Molecular Sciences. 2019; 20(15):3629. https://doi.org/10.3390/ijms20153629
Chicago/Turabian StyleTanaka, Yasuhisa, Toshihide Kurihara, Yumi Hagiwara, Shin-ichi Ikeda, Kiwako Mori, Xiaoyan Jiang, Hidemasa Torii, and Kazuo Tsubota. 2019. "Ocular-Component-Specific miRNA Expression in a Murine Model of Lens-Induced Myopia" International Journal of Molecular Sciences 20, no. 15: 3629. https://doi.org/10.3390/ijms20153629