Diagnostic Performance of Dengue Virus Envelope Domain III in Acute Dengue Infection

Abstract
:
1. Introduction
2. Results
2.1. Expression of Recombinant Envelope Domain III Proteins
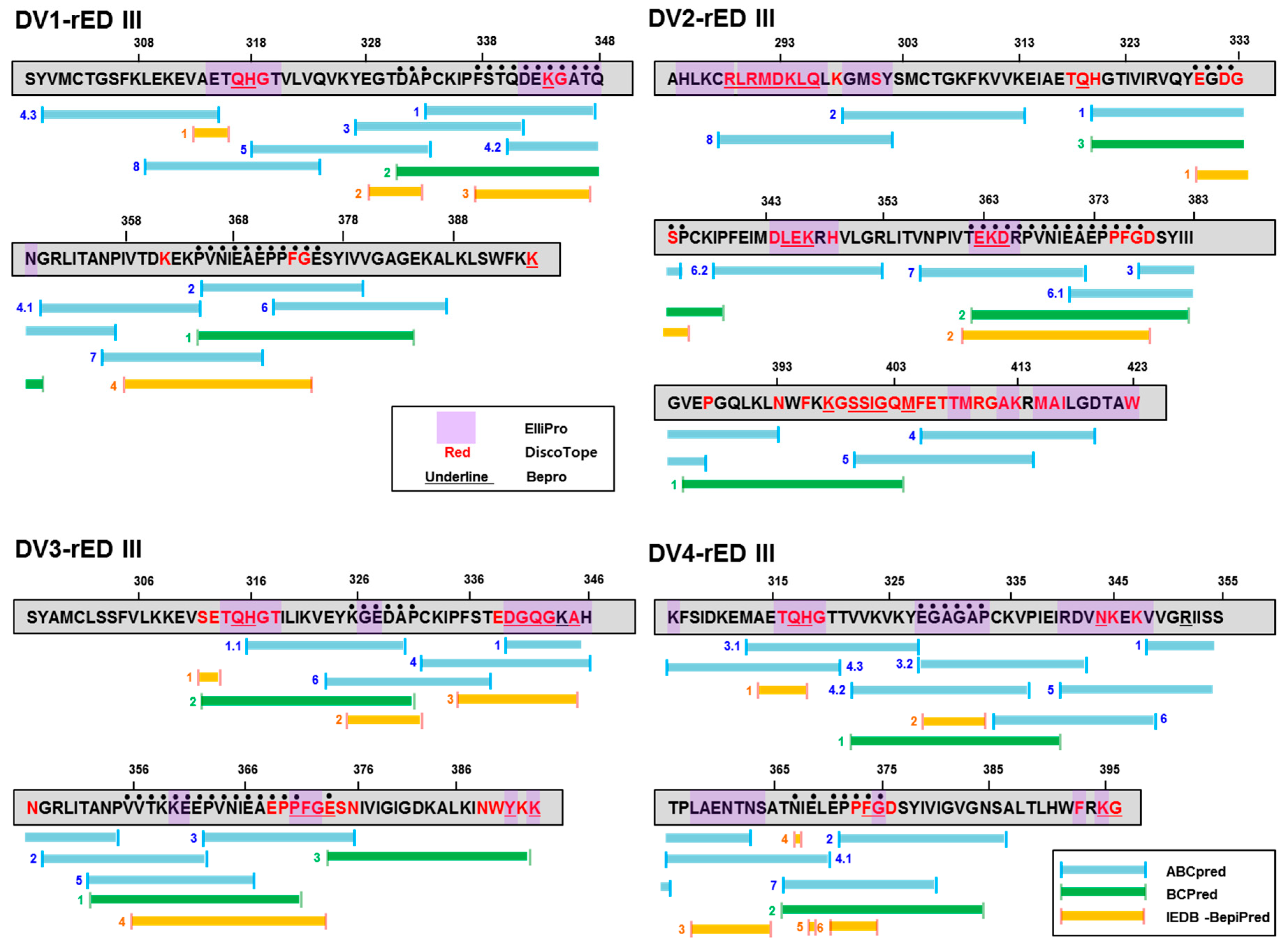
2.2. Prediction of Antigenic Epitope in Recombinant Antigens
2.3. Prediction of Position of Antigenic Epitope in Recombinant Antigens
2.4. Clinical Cohort Study
2.5. DENV-Linked ELISA to Detect IgM in Clinical Samples
2.6. Recombinant Antigen-Linked ELISA to Detect IgM in Clinical Samples
3. Discussion
4. Materials and Methods
4.1. Cell and Virus
4.2. Reagents
4.3. Expression of Recombinant Envelope Domain III Antigen
4.4. RT-PCR
4.5. ELISA
4.6. ED III Sequence Analysis and Protein Structure Graphic
4.7. Ethics Statement
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Dengue and Severe Dengue. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 15 April 2019).
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Gatell, H.; Alpuche-Aranda, C.M.; Santos-Preciado, J.I.; Hernandez-Avila, M. Dengue vaccine: Local decisions, global consequences. Bull. World Health Organ. 2016, 94, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Fry, S.R.; Meyer, M.; Semple, M.G.; Simmons, C.P.; Sekaran, S.D.; Huang, J.X.; McElnea, C.; Huang, C.Y.; Valks, A.; Young, P.R.; et al. The diagnostic sensitivity of dengue rapid test assays is significantly enhanced by using a combined antigen and antibody testing approach. PLoS Negl. Trop. Dis. 2011, 5, e1199. [Google Scholar] [CrossRef] [PubMed]
- Chatchen, S.; Sabchareon, A.; Sirivichayakul, C. Serodiagnosis of asymptomatic dengue infection. Asian Pac. J. Trop Med. 2017, 10, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.J.; Lin, P.C.; Tsai, C.Y.; Wang, Y.H.; Liu, L.T. Low frequency of asymptomatic dengue virus-infected donors in blood donor centers during the largest dengue outbreak in Taiwan. PLoS ONE 2018, 13, e0205248. [Google Scholar] [CrossRef] [PubMed]
- Halasa, Y.A.; Shepard, D.S.; Zeng, W. Economic cost of dengue in Puerto Rico. Am. J. Trop Med. Hyg 2012, 86, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Pozzetto, B.; Memmi, M.; Garraud, O. Is transfusion-transmitted dengue fever a potential public health threat? World J. Virol. 2015, 4, 113–123. [Google Scholar] [CrossRef]
- Muñoz-Jordán, J.L.; Collins, C.S.; Vergne, E.; Santiago, G.A.; Petersen, L.; Sun, W.; Linnen, J.M. Highly sensitive detection of dengue virus nucleic acid in samples from clinically ill patients. J. Clin. Microbiol. 2009, 47, 927–931. [Google Scholar] [CrossRef]
- Libraty, D.H.; Endy, T.P.; Houng, H.S.; Green, S.; Kalayanarooj, S.; Suntayakorn, S.; Chansiriwongs, W.; Vaughn, D.W.; Nisalak, A.; Ennis, F.A.; et al. Differing influences of virus burden and immune activation on disease severity in secondary dengue-3 virus infections. J. Infect. Dis. 2002, 185, 1213–1221. [Google Scholar] [CrossRef]
- Chaterji, S.; Allen, J.C., Jr.; Chow, A.; Leo, Y.S.; Ooi, E.E. Evaluation of the NS1 rapid test and the WHO dengue classification schemes for use as bedside diagnosis of acute dengue fever in adults. Am. J. Trop Med. Hyg. 2011, 84, 224–228. [Google Scholar] [CrossRef]
- Chen, W.H.; Chou, F.P.; Wang, Y.K.; Huang, S.C.; Cheng, C.H.; Wu, T.K. Characterization and epitope mapping of Dengue virus type 1 specific monoclonal antibodies. Virol J. 2017, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Parkash, O.; Shueb, R.H. Diagnosis of Dengue Infection Using Conventional and Biosensor Based Techniques. Viruses 2015, 7, 5410–5427. [Google Scholar] [CrossRef] [PubMed]
- Matheus, S.; Boukhari, R.; Labeau, B.; Ernault, V.; Bremand, L.; Kazanji, M.; Rousset, D. Specificity of Dengue NS1 Antigen in Differential Diagnosis of Dengue and Zika Virus Infection. Emerg. Infect. Dis 2016, 22, 1691–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, I.; de Puig, H.; Hiley, M.; Carré-Camps, M.; Perdomo-Celis, F.; Narváez, C.F.; Salgado, D.M.; Senthoor, D.; O’Grady, M.; Phillips, E.; et al. Rapid antigen tests for dengue virus serotypes and Zika virus in patient serum. Sci. Transl. Med. 2017, 9, 1589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jia, R.; Shen, H.; Wang, M.; Yin, Z.; Cheng, A. Structures and Functions of the Envelope Glycoprotein in Flavivirus Infections. Viruses 2017, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Prince, H.E.; Matud, J.L. Estimation of dengue virus IgM persistence using regression analysis. Clin. Vaccine. Immunol. 2011, 18, 2183–2185. [Google Scholar] [CrossRef] [PubMed]
- Shu, P.Y.; Huang, J.H. Current advances in dengue diagnosis. Clin. Diagn. Lab. Immunol. 2004, 11, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Crill, W.D.; Hughes, H.R.; Delorey, M.J.; Chang, G.J. Humoral immune responses of dengue fever patients using epitope-specific serotype-2 virus-like particle antigens. PLoS ONE 2009, 4, e4991. [Google Scholar] [CrossRef] [PubMed]
- McBurney, S.P.; Sunshine, J.E.; Gabriel, S.; Huynh, J.P.; Sutton, W.F.; Fuller, D.H.; Haigwood, N.L.; Messer, W.B. Evaluation of protection induced by a dengue virus serotype 2 envelope domain III protein scaffold/DNA vaccine in non-human primates. Vaccine 2016, 34, 3500–3507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poddar, A.; Ramasamy, V.; Shukla, R.; Rajpoot, R.K.; Arora, U.; Jain, S.K.; Swaminathan, S.; Khanna, N. Virus-like particles derived from Pichia pastoris-expressed dengue virus type 1 glycoprotein elicit homotypic virus-neutralizing envelope domain III-directed antibodies. BMC Biotechnol. 2016, 16, 50. [Google Scholar] [CrossRef] [PubMed]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.; Collins, M.; Graham, S.; Liou, G.A.; Lopez, C.A.; Jadi, R.; Balmaseda, A.; Brackbill, J.A.; Dietze, R.; Camacho, E.; et al. Development of Envelope Protein Antigens To Serologically Differentiate Zika Virus Infection from Dengue Virus Infection. J. Clin. Microbiol. 2018, 56, e01504–e01517. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.A.; Paixão, V.F.; Oliveira, M.D.; Honda, E.R.; Oliveira, L.L.; da Silva, C.C.; de Paula, S.O. Dengue-1 envelope protein domain III produced in Pichia pastoris: potential use for serological diagnosis. Protein Expr. Purif. 2013, 92, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Babu, J.P.; Pattnaik, P.; Gupta, N.; Shrivastava, A.; Khan, M.; Rao, P.V. Immunogenicity of a recombinant envelope domain III protein of dengue virus type-4 with various adjuvants in mice. Vaccine 2008, 26, 4655–4663. [Google Scholar] [CrossRef] [PubMed]
- Ponomarenko, J.; Bui, H.H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef] [PubMed]
- Sweredoski, M.J.; Baldi, P. PEPITO: Improved discontinuous B-cell epitope prediction using multiple distance thresholds and half sphere exposure. Bioinformatics 2008, 24, 1459–1460. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Liu, X.; Baker, T.; Shi, J.; Deane, C.M. Improving B-cell epitope prediction and its application to global antibody-antigen docking. Bioinformatics 2014, 30, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.M.; Sekaran, S.D. Early diagnosis of Dengue infection using a commercial Dengue Duo rapid test kit for the detection of NS1, IGM, and IGG. Am. J. Trop Med. Hyg 2010, 83, 690–695. [Google Scholar] [CrossRef]
- Bagut, E.T.; Cambier, L.; Heinen, M.P.; Cozma, V.; Monod, M.; Mignon, B. Development of an enzyme-linked immunosorbent assay for serodiagnosis of ringworm infection in cattle. Clin. Vaccine Immunol. 2013, 20, 1150–1154. [Google Scholar] [CrossRef]
- Burke, D.S.; Nisalak, A.; Johnson, D.E.; Scott, R.M. A prospective study of dengue infections in Bangkok. Am. J. Trop Med. Hyg 1988, 38, 172–180. [Google Scholar] [CrossRef]
- David, W.V.; Sharone, G.; Siripen, K.; Bruce, L.I.; Suchitra, N.; Saroj, S.; Timothy, P.E.; Boonyos, R.; Alan, L.R.; Francis, A.E.; et al. Dengue Viremia Titer, Antibody Response Pattern, and Virus Serotype Correlate with Disease Severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar]
- Wright, W.F.; Pritt, B.S. Update: The diagnosis and management of dengue virus infection in North America. Diagn Microbiol. Infect. Dis 2012, 73, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Stiasny, K. The Antigenic Structure of Zika Virus and Its Relation to Other Flaviviruses: Implications for Infection and Immunoprophylaxis. Microbiol. Mol. Biol. Rev. 2017, 81, e00055-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Testing Guidance. 2019. Available online: https://www.cdc.gov/dengue/healthcare-providers/testing/testing-guidance.html (accessed on 13 June 2019).
- Thomas, L.; Najioullah, F.; Verlaeten, O.; Martial, J.; Brichler, S.; Kaidomar, S.; Moravie, V.; Cabié, A.; Césaire, R. Relationship between nonstructural protein 1 detection and plasma virus load in Dengue patients. Am. J. Trop Med. Hyg 2010, 83, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Mat, J.T.; Shueb, R.H. Performance Evaluation of Commercial Dengue Diagnostic Tests for Early Detection of Dengue in Clinical Samples. J. Trop Med. 2017, 2017, 4687182. [Google Scholar]
- Shukla, M.K.; Singh, N.; Sharma, R.K.; Barde, P.V. Utility of dengue NS1 antigen rapid diagnostic test for use in difficult to reach areas and its comparison with dengue NS1 ELISA and qRT-PCR. J. Med. Virol. 2017, 89, 1146–1150. [Google Scholar] [CrossRef]
- Hermann, L.L.; Thaisomboonsuk, B.; Poolpanichupatam, Y.; Jarman, R.G.; Kalayanarooj, S.; Nisalak, A.; Yoon, I.K.; Fernandez, S. Evaluation of a dengue NS1 antigen detection assay sensitivity and specificity for the diagnosis of acute dengue virus infection. PLoS Negl Trop Dis. 2014, 8, e3193. [Google Scholar] [CrossRef]
- Chen, J.; Wen, K.; Li, X.; Yi, H.; Ding, X.; Huang, Y.; Pan, Y.; Hu, D.; Di, B.; Che, X.; et al. Functional properties of DENV EDIII-reactive antibodies in human DENV-1-infected sera and rabbit antiserum to EDIII. Mol. Med. Rep. 2016, 14, 1799–1808. [Google Scholar] [CrossRef]
- Niu, G.; Pang, Z.; Guan, C.; Qi, J.; Li, D. Dengue virus envelope domain III protein based on a tetravalent antigen secreted from insect cells: Potential use for serological diagnosis. Virus Res. 2015, 201, 73–78. [Google Scholar] [CrossRef]
- Batra, G.; Nemani, S.K.; Tyagi, P.; Swaminathan, S.; Khanna, N. Evaluation of envelope domain III-based single chimeric tetravalent antigen and monovalent antigen mixtures for the detection of anti-dengue antibodies in human sera. BMC Infect. Dis. 2011, 11, 64. [Google Scholar] [CrossRef]
- Murgue, B.; Roche, C.; Chungue, E.; Deparis, X. Prospective study of the duration and magnitude of viraemia in children hospitalised during the 1996-1997 dengue-2 outbreak in French Polynesia. J. Med. Virol. 2000, 60, 432–438. [Google Scholar] [CrossRef]
- Jaiswal, S.; Khanna, N.; Swaminathan, S. High-level expression and one-step purification of recombinant dengue virus type 2 envelope domain III protein in Escherichia coli. Protein Expr. Purif. 2004, 33, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.S.; Kwak, S.Y.; May, W.L.; Yang, D.J.; Nam, J.; Lim, C.S. Comparative evaluation of three dengue duo rapid test kits to detect NS1, IgM, and IgG associated with acute dengue in children in Myanmar. PLoS ONE 2019, 14, e0213451. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Russell, B.J.; Lanciotti, R.S. Serotype-specific detection of dengue viruses in a fourplex real-time reverse transcriptase PCR assay. J. Clin. Microbiol. 2005, 43, 4977–4983. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.T.H.; Bao, D.T.; Park, H.; Ngoc, N.M.; Yeo, S.J. Development of a novel peptide aptamer-based immunoassay to detect Zika virus in serum and urine. Theranostics 2018, 8, 3629–3642. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef] [PubMed]
- El-Manzalawy, Y.; Dobbs, D.; Honavar, V. Predicting linear B-cell epitopes using string kernels. J. Mol. Recognit. 2008, 21, 243–255. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Haste, A.P.; Nielsen, M.; Lund, O. Prediction of residues in discontinuous B-cell epitopes using protein 3D structures. Protein Sci. 2006, 15, 2558–2567. [Google Scholar] [CrossRef]
- Potocnakova, L.; Bhide, M.; Pulzova, L.B. An Introduction to B-Cell Epitope Mapping and In Silico Epitope Prediction. J. Immunol. Res. 2016, 2016, 6760830. [Google Scholar] [CrossRef]
- Yeo, S.J.; Huong, D.T.; Han, J.H.; Kim, J.Y.; Lee, W.J.; Shin, H.J.; Han, E.T.; Park, H. Performance of coumarin-derived dendrimer-based fluorescence-linked immunosorbent assay (FLISA) to detect malaria antigen. Malar J. 2014, 13, 266. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dengue Type | Linear | Conformational | ||
|---|---|---|---|---|
| DV1-rED III | L1-1 L1-2 L1-3 | DAP (331-333) a FSTQDEKGATQ (337-347) PVNIEAEPPFG (365-375) | C1-1 C1-2 | QH (317-318) K (344) |
| DV2-rED III | L2-1 L2-2 | EGDGSP (330-335) TEKDRPVNIEAEPPFG (362-377) | C2-1 C2-2 C2-3 | RLRMDKLQ (289-296) LEK (345-347) EKD (363-365) |
| DV3-rED III | L3-1 L3-2 L3-3 | KGEDAP (326-331) VVTKKEEPVNIEAEPP (356-371) E (374) | C3-1 C3-2 C3-3 C3-4 C3-5 C3-6 | QH (315-316) DGQG (340-343) A (345) PFG (371-373) Y (391) K (393) |
| DV4-rED III | L4-1 L4-2 L4-3 L4-4 | EGAGAP (328-333) N (367) E (369) EPPFG (371-375) | C4-1 C4-2 C4-3 C4-4 | QH (317-318) N (344) G (375) K (395) |
| Antigens | µg/well | Cut-off | Sensitivity (n = 22) | Specificity (n = 46) | ||
|---|---|---|---|---|---|---|
| % (Positive No.) | 95% CI a | % (Negative No.) | 95% CI | |||
| DV1-rED III | 0.1 | 0.269527 | 45.45 (10) | 24.39 to 67.79 | 100.00 (46) | 92.29 to 100.00 |
| 0.5 b | 0.276739 | 59.09 (13) | 36.35 to 79.29 | 100.00 (46) | 92.29 to 100.00 | |
| 1 | 0.286781 | 50.00 (11) | 28.22 to 71.78 | 100.00 (46) | 92.29 to 100.00 | |
| 5 | 0.270051 | 45.45 (10) | 24.39 to 67.79 | 100.00 (46) | 92.29 to 100.00 | |
| DV2-rED III | 0.1 | 0.314 | 27.27 (6) | 10.73 to 50.22 | 100.00 (46) | 92.29 to 100.00 |
| 0.5 | 0.282 | 31.82 (7) | 13.86 to 54.87 | 100.00 (46) | 92.29 to 100.00 | |
| 1 | 0.295 | 40.91 (9) | 20.71 to 63.65 | 100.00 (46) | 92.29 to 100.00 | |
| 5 | 0.314 | 40.91 (9) | 20.71 to 63.65 | 100.00 (46) | 92.29 to 100.00 | |
| DV3-rED III | 0.1 | 0.324308 | 54.55 (12) | 32.21 to 75.61 | 100.00 (46) | 92.29 to 100.00 |
| 0.5 | 0.434963 | 45.45 (10) | 24.39 to 67.79 | 100.00 (46) | 92.29 to 100.00 | |
| 1 | 0.481123 | 54.55 (12) | 32.21 to 75.61 | 100.00 (46) | 92.29 to 100.00 | |
| 5 | 0.483615 | 59.09 (13) | 36.35 to 79.29 | 100.00 (46) | 92.29 to 100.00 | |
| DV4-rED III | 0.1 | 0.402995 | 31.82 (7) | 13.86 to 54.87 | 100.00 (46) | 92.29 to 100.00 |
| 0.5 | 0.477119 | 50.00 (11) | 28.22 to 71.78 | 100.00 (46) | 92.29 to 100.00 | |
| 1 | 0.606018 | 36.36 (8) | 17.20 to 59.34 | 100.00 (46) | 92.29 to 100.00 | |
| 5 | 0.492167 | 45.45 (10) | 24.39 to 67.79 | 100.00 (46) | 92.29 to 100.00 | |
| Patients | Days after Onset of Disease | RT-PCR (Dengue Type/Ct) | RT-PCR (Zika Virus/Ct) | ELISA | |||
|---|---|---|---|---|---|---|---|
| DV1-rED III (0.5 µg/well) | DV2-rED III (5 µg/well) | DV3-rED III (5 µg/well) | DV4-rED III (0.5 µg/well) | ||||
| VN-1 | 3 | 1(22.72) | N | N | N | N | N |
| VN-2 | 3 | 1(25.99) | N | N | N | N | N |
| VN-3 | 2 | 1(29.15) | N | P b | N | P | P |
| VN-4 | 2 | 1(30.15) | N | P | P | P | N |
| VN-5 | 3 | 2(35.03) | N | N | N | P | N |
| VN-6 | 4 | 1(27.06) | N | P | P | P | P |
| VN-7 | 3 | 1(35.66) | N | P | N | N | N |
| VN-8 | 2 | N a | N | P | P | N | N |
| VN-9 | 3 | N | N | P | P | P | N |
| VN-10 | 3 | 1(25.17) | N | P | P | P | P |
| VN-11 | 3 | 1(35.57)/2(35.89) | N | N | N | N | N |
| VN-12 | 2 | N | N | P | N | N | P |
| VN-13 | 3 | N | N | N | N | P | P |
| VN-14 | 2 | 2(33.83) | N | P | P | P | P |
| VN-15 | 2 | 1(23.19) | N | N | N | N | P |
| VN-16 | 2 | 1(22.43) | N | N | N | N | N |
| VN-17 | 3 | 1(35.76) | N | N | N | P | P |
| VN-18 | 2 | 2(35.50) | N | N | N | P | N |
| VN-19 | 3 | 1(31.50) | N | P | N | P | P |
| VN-20 | 2 | 1(24.64) | N | P | P | P | P |
| VN-24 | 3 | 1(33.31) | N | P | P | P | P |
| VN-25 | 3 | N | N | P | P | N | N |
| rRT-PCR (Dengue) | ELISA (DV1-rED III) + ELISA (DV3-rED III) + ELISA (DV4-rED III) | Total (n = 22) | ||||||
|---|---|---|---|---|---|---|---|---|
| 2 Days after Onset of Disease | 3 Days after Onset of Disease | 4 Days after Onset of Disease | ||||||
| Neg. | Pos. | |||||||
| Negative | Positive | Negative | Positive | Negative | Positive | |||
| 20 < Ct ≤ 30 (n = 8) | 1 (VN16 1,2,3,4) | 3 (VN3 1,3,4; VN15 4; VN20 1,2,3,4) | 2 (VN1 1,2,3,4; VN2 1,2,3,4) | 1 (VN10 1,2,3,4) | 1 (VN6 1,2,3,4) | 3 | 5 | |
| 30 < Ct ≤ 35 (n = 4) | 2 (VN4 1,2,3; VN14 1,2,3,4) | 2 (VN19 1,3,4; VN24 1,2,3,4) | 4 | |||||
| 35 < Ct (n = 5) | 1 (VN18 3) | 1 (VN11 1,2,3,4) | 3 (VN5 3; VN7 1; VN17 3,4) | 1 | 4 | |||
| Negative (n = 5) | 2 (VN8 1,2; VN12 1,4) | 3 (VN9 1,2,3; VN13 3,4; VN25 1,2) | 5 | |||||
| Total (n = 22) | 1 | 8 | 3 | 9 | 1 | 4 | 18 | |
| Parameter | DV1-rED III | DV2-rED III | DV3-rED III | DV4-rED III | Combined Result of DV1-, DV3-, and DV4-rED III |
|---|---|---|---|---|---|
| Sensitivity% (95% CI) | 59.09 (13/22) (36.35 to 79.29) | 40.91 (9/22) (20.71 to 63.65) | 59.09 (13/22) (36.35 to 79.29) | 50.00 (11/22) (28.22 to 71.78) | 81.82 (18/22) (59.72 to 94.81) |
| Specificity% (95% CI) | 100.00 (46/46) (92.29 to 100.00) | 100.00 (46/46) (92.29 to 100.00) | 100.00 (46/46) (92.29 to 100.00) | 100.00 (46/46) (92.29 to 100.00) | 100.00 (46/46) (92.29 to 100.00) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, N.M.; Duong, B.T.; Azam, M.; Phuong, T.T.; Park, H.; Thuy, P.T.B.; Yeo, S.-J. Diagnostic Performance of Dengue Virus Envelope Domain III in Acute Dengue Infection. Int. J. Mol. Sci. 2019, 20, 3464. https://doi.org/10.3390/ijms20143464
Nguyen NM, Duong BT, Azam M, Phuong TT, Park H, Thuy PTB, Yeo S-J. Diagnostic Performance of Dengue Virus Envelope Domain III in Acute Dengue Infection. International Journal of Molecular Sciences. 2019; 20(14):3464. https://doi.org/10.3390/ijms20143464
Chicago/Turabian StyleNguyen, Ngoc Minh, Bao Tuan Duong, Mudsser Azam, Truong Thai Phuong, Hyun Park, Phung Thi Bich Thuy, and Seon-Ju Yeo. 2019. "Diagnostic Performance of Dengue Virus Envelope Domain III in Acute Dengue Infection" International Journal of Molecular Sciences 20, no. 14: 3464. https://doi.org/10.3390/ijms20143464