Aluminum Alters the Histology and Pectin Cell Wall Composition of Barley Roots

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Root Histology is Altered by Al Treatment
2.2. Al Treatment Affects the Distribution of the Pectic Epitopes in Barley Roots
2.2.1. LM5
2.2.2. LM6
2.2.3. LM19
2.2.4. LM20
2.3. Summary
3. Discussion
4. Materials and Methods
4.1. Plant Material Growth and Treatment
4.2. Histological Procedures
4.3. Immunohistochemistry

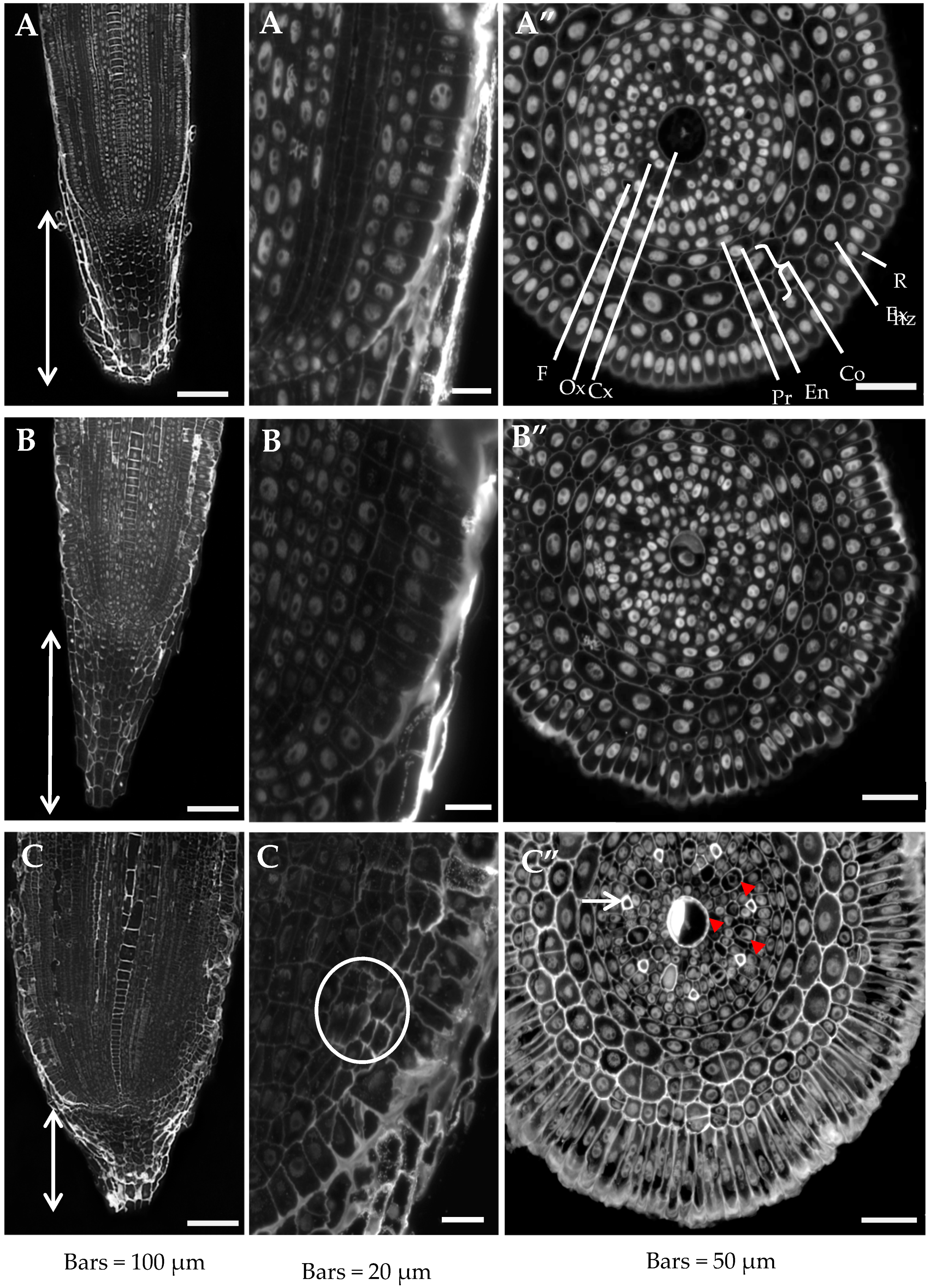{kind=link}
{kind=link}
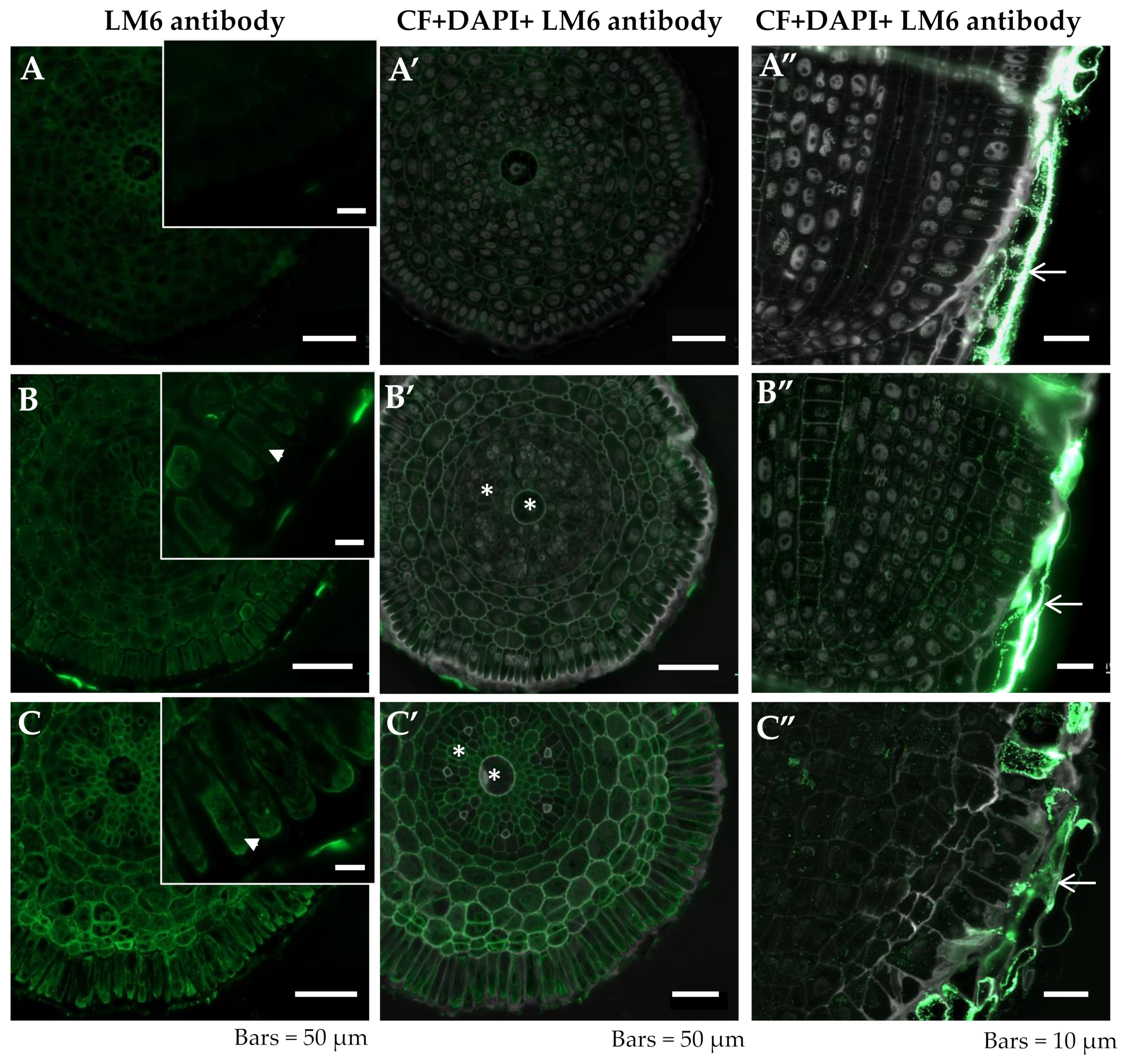{kind=link}
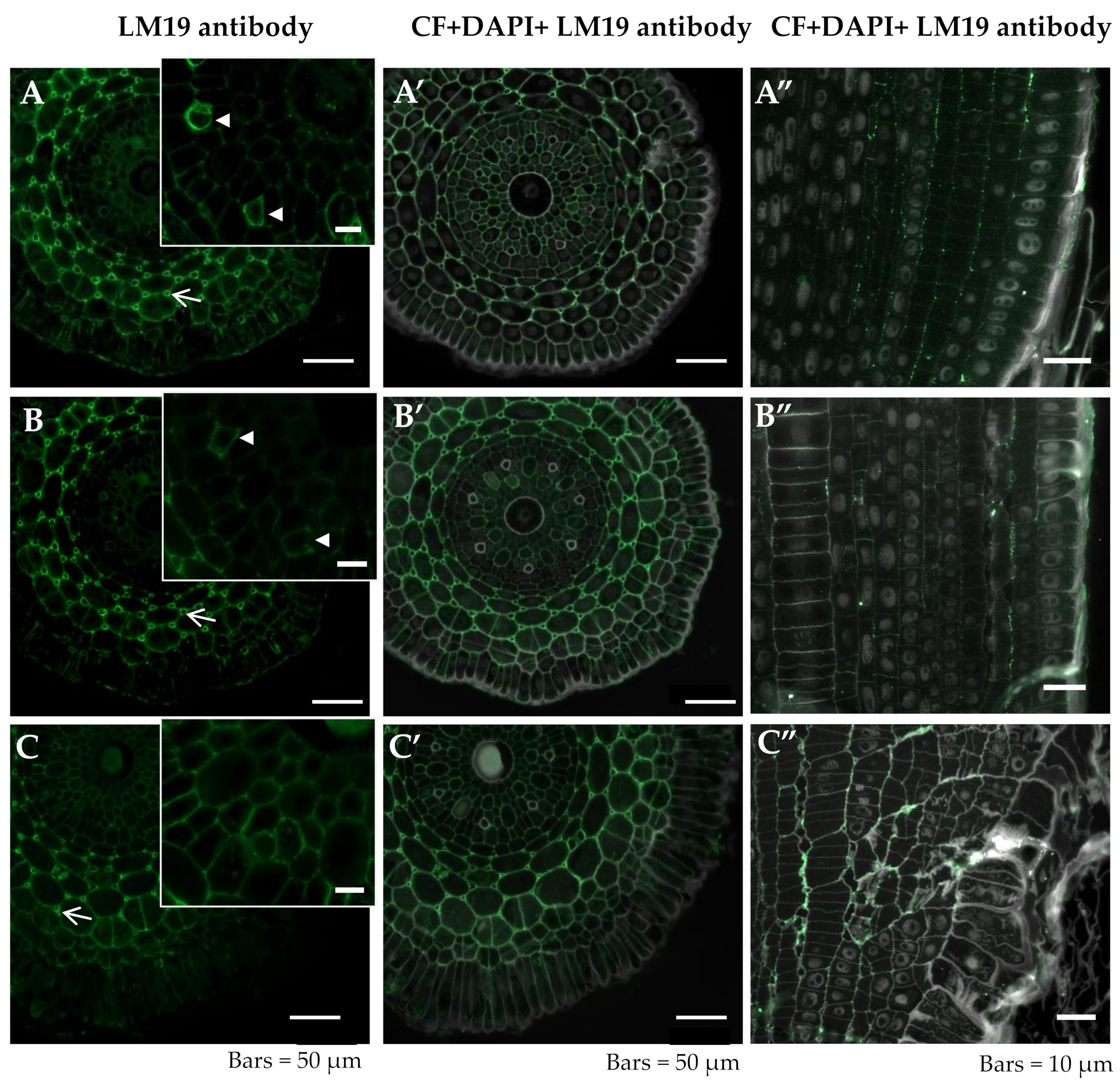{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Recognized Epitope | Reference |
|---|---|---|
| Pectins—rhamnogalacturonan I (RG I) and homogalacturonan (HG) | ||
| LM5 | linear tetrasaccharide in (1-4)-β-D-galactans (RG I side chain) | [58] |
| LM6 | linear pentasaccharide in (1-5)-α-L-arabinans (RG I side chain) | [59] |
| LM19 | unmethyl-esterified, partially methyl-esterified HG | [51] |
| LM20 | methyl-esterified HG | [51] |
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A


References
- Kochian, L.V.; Hoekenga, O.A.; Pineros, M.A. How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 2004, 55, 459–493. [Google Scholar] [CrossRef] [PubMed]
- Foy, C.D. Soil chemical factors limiting plant root growth. In limitations to plant root growth. In Advances in Soil Science; Hatfield, J.L., Stewart, B.A., Eds.; Springer: New York, NY, USA, 1992. [Google Scholar]
- Bot, A.J.; Nachtergaele, F.O.; Young, A. Land Re-source Potential and Constraints at Regional and Country Levels; Food and Agricultural Organization of the United Nations: Rome, Italy, 2000; p. 114. [Google Scholar]
- Gupta, N.; Gaurav, S.S.; Kumar, A. Molecular basis of aluminum toxicity in plants: A review. Am. J. Plant Sci. 2013, 4, 21–37. [Google Scholar] [CrossRef]
- Silva, S. Aluminium toxicity targets in plants. J. Bot. 2012, 2012. [Google Scholar] [CrossRef]
- Rounds, M.A.; Larsen, P.B. Aluminum-dependent root-growth inhibition in Arabidopsis results from AtATR-regulated cell-cycle arrest. Curr. Biol. 2008, 18, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Sjogren, C.A.; Larsen, P.B.; Schnittger, A. A multi-level response to DNA damage induced by Aluminium. Plant J. 2019, 93, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, A.; Matsumoto, H. Changes in cell-wall properties of wheat (Triticum aestivum) roots during aluminum-induced growth inhibition. Physiol. Plant. 2001, 112, 353–358. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, J.L.; He, L.S.; Li, Y.Y.; Zheng, S.J. Effect of aluminum on cell wall, plasma membrane, antioxidants and root elongation in triticale. Biol. Plant. 2008, 52, 87–92. [Google Scholar] [CrossRef]
- Wang, J.; Raman, H.; Zhang, G.; Mendham, N.; Zhou, M. Aluminum tolerance in barley (Hordeum vulgare L.) physiological mechanisms, genetics and screening methods. J. Zhejiang Univ. Sci. B 2006, 7, 769–787. [Google Scholar] [CrossRef]
- Jaskowiak, J.; Tkaczyk, O.; Slota, M.; Kwasniewska, J.; Szarejko, I. Analysis of aluminum toxicity in Hordeum vulgare roots with an emphasis on DNA integrity and cell cycle. PLoS ONE 2018, 13, e0193156. [Google Scholar] [CrossRef]
- Zobel, R.W.; Kinraide, T.B.; Baligar, V.C. Fine root diameters can change in response to changes in nutrient concentrations. Plant Soil 2007, 297, 243–254. [Google Scholar] [CrossRef]
- Horst, W.J. The role of the apoplast in aluminium toxicity and resistance of higher plants: A review. J. Plant Nutr. Soil Sci. 1995, 158, 419–428. [Google Scholar] [CrossRef]
- Ma, J.F.; Shen, R.F.; Nagao, S.; Tanimoto, E. Aluminum targets elongating cells by reducing cell wall extensibility in wheat roots. Plant Cell Physiol. 2004, 45, 583–589. [Google Scholar] [CrossRef]
- Jones, D.L.; Blancaflor, E.B.; Kochian, L.V.; Gilroy, S. Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant Cell Environ. 2006, 29, 1309–1318. [Google Scholar] [CrossRef]
- Ma, J.F. Syndrome of aluminum toxicity and diversity of aluminum resistance in higher plants. Int. Rev. Cytol. 2007, 264, 225–252. [Google Scholar]
- Yang, Z.-B.; Horst, W.J. Aluminum-Induced Inhibition of Root Growth: Roles of Cell Wall Assembly, Structure and Function. In Aluminum Stress Adaptation in Plants, Signalling and Communication in Plants 24; Panda, S.K., Baluska, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Schmohl, N.; Horst, W.J. Cell wall pectin content modulates aluminum sensitivity of Zea mays (L.) cells grown in suspension culture. Plant Cell Environ. 2000, 23, 735–742. [Google Scholar] [CrossRef]
- Schmohl, N.; Pilling, J.; Fisahn, J.; Horst, W.J. Pectin methylesterase modulates aluminum sensitivity in Zea mays and Solanum tuberosum. Physiol. Plant. 2000, 109, 419–427. [Google Scholar] [CrossRef]
- Hossain, A.K.M.Z.; Koyama, H.; Hara, T. Growth and cell wall properties of two wheat cultivars differing in their sensitivity to aluminum stress. J. Plant Physiol. 2006, 163, 39–47. [Google Scholar] [CrossRef]
- Sun, C.h.; Lu, L.; Yu, Y.; Liu, L.; Hu, Y.; Ye, Y.; Jin, C.; Lin, X. Decreasing methylation of pectin caused by nitric oxide leads to higher aluminium binding in cell walls and greater aluminium sensitivity of wheat roots. J. Exp. Bot. 2015, 67, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef]
- Milewska-Hendel, A.; Baczewska, A.H.; Sala, K.; Dmuchowski, W.; Bragoszewska, P.; Gozdowski, D.; Jozwiak, A.; Chojnacki, T.; Swiezewska, E.; Kurczynska, E. Quantitative and qualitative characteristics of cell wall components and prenyl lipids in the leaves of Tilia x euchlora trees growing under salt stress. PLoS ONE 2017, 12, e0172682. [Google Scholar] [CrossRef]
- Matsumoto, H.; Riechers, D.E.; Lygin, A.V.; Baluska, F.; Sivaguru, M. Aluminum Signaling and Potential Links with Safener-Induced Detoxification in Plants. In Aluminum Stress Adaptation in Plants, Signaling and Communication in Plants; Panda, S.K., Baluska, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Horst, W.J.; Wang, Y.; Eticha, D. The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: A review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef]
- Geng, X.; Horst, W.J.; Golz, J.F.; Lee, J.E.; Ding, Z.; Yang, Z.-B. LEUNIG_HOMOLOG transcriptional co-repressor mediates aluminium sensitivity through PECTIN METHYLESTERASE46-modulated root cell wall pectin methylesterification in Arabidopsis. Plant J. 2017, 90, 491–504. [Google Scholar] [CrossRef]
- Eticha, D.; Stass, A.; Horst, W.J. Cell-wall pectin and its degree of methylation in the maize root-apex: Significance for genotypic differences in aluminium resistance. Plant Cell Environ. 2005, 28, 1410–1420. [Google Scholar] [CrossRef]
- Baldwin, L.; Domon, J.-M.; Klimek, J.F.; Fournet, F.; Sellier, H.; Gillet, F.; Pelloux, J.; Lejeune-Hénaut, I.; Carpita, N.C.; Rayon, C. Structural alteration of cell wall pectins accompanies pea development in response to cold. Phytochemistry 2014, 104, 37–47. [Google Scholar] [CrossRef]
- Muschitz, A.; Riou, C.; Mollet, J.-C.; Gloaguen, V.; Faugeron, C. Modifications of cell wall pectin in tomato cell suspension in response to cadmium and zinc. Acta Physiol. Plant. 2015, 37, 245. [Google Scholar] [CrossRef]
- Knox, J.P. Revealing the structural and functional diversity of plant cell walls. Curr. Opin. Plant Biol. 2008, 11, 308–313. [Google Scholar] [CrossRef]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef]
- Sjogren, C.A.; Larsen, P.B. Aluminum-dependent root growth inhibition as mediated by DNA damage responses. In Aluminum Stress Adaptation in Plants, Signaling and Communication in Plants 24; Panda, S.K., Baluska, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Foy, C.D. Plant adaption to acid, aluminum-toxic soils. Commun. Soil Sci. Plant Anal. 1988, 19, 959–987. [Google Scholar] [CrossRef]
- Horst, W.J.; Puschel, A.-K.; Schmohl, N. Induction of callose formation is a sensitive marker for genotypic aluminium sensitivity in maize. Plant Soil. 1997, 192, 23–30. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Tolra, R.; Hajiboland, R.; Arroyave, C.; Barcelo, J. Mechanisms of hyper-resistance and hyper-tolerance to aluminum in plants. In Aluminum Stress Adaptation in Plants, Signaling and Communication in Plants; Panda, S.K., Baluska, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Blancaflor, E.B.; Jones, D.L.; Gilroy, S. Alterations in the cytoskeleton accompany aluminium-induced growth inhibition and morphological changes in primary roots of maize. Plant Physiol. 1998, 118, 159–172. [Google Scholar] [CrossRef]
- Ryan, P.R.; Ditomaso, J.M.; Kochian, L.V. Aluminium toxicity in roots: And investigation of spatial sensitivity and the role of root cap. J. Exp. Bot. 1993, 44, 437–446. [Google Scholar] [CrossRef]
- Bennet, R.J.; Breen, C.M.; Fey, M.V. The effects of aluminium on root cap function and root development in Zea mays L. Environ. Exp. Bot. 1987, 27, 21–27. [Google Scholar] [CrossRef]
- He, H.-Y.; Gu, M.-H.; He, L.-F. Signaling Pathways of Aluminum-Induced Programmed Cell Death in Higher Plants. In Aluminum Stress Adaptation in Plants, Signaling and Communication in Plants; Panda, S.K., Baluska, F., Eds.; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar]
- Yamamoto, Y.; Matsumoto, H.; Hachiya, A. Oxidative Damage to Membranes by a Combination of Aluminum and Iron in Suspension-Cultured Tobacco Cells. Plant Cell Physiol. 1997, 38, 1333–1339. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Zhu, M.; Chen, H. Aluminum-induced cell death in root-tip cells of barley. Environ. Exp. Bot. 2001, 46, 71–79. [Google Scholar] [CrossRef]
- Boscolo, P.R.S.; Menossi, M.; Jorge, R.A. Aluminum-induced oxidative stress in maize. Phytochemistry 2003, 62, 181–189. [Google Scholar] [CrossRef]
- Matsumoto, H. Cell biology of aluminum toxicity and tolerance in higher plants. Int. Rev. Cytol. 2000, 200, 1–46. [Google Scholar]
- Doncheva, S.; Amenos, M.; Poschenrieder, C.; Barcelo, J. Root cell patterning: A primary target for aluminium toxicity in maize. J. Exp. Bot. 2005, 56, 1213–1230. [Google Scholar] [CrossRef]
- Amenos, M.; Corrales, I.; Poschenrieder, C.; Illes, P.; Baluska, F.; Barcelo, J. Different effects of aluminum on the actin cytoskeleton and brefeldin A-sensitive vesicle recycling in root apex cells of two maize varieties differing in root elongation rate and aluminum tolerance. Plant Cell Physiol. 2009, 50, 528–540. [Google Scholar] [CrossRef]
- Miyasaka, S.C.; Hawes, M.C. Possible role of root border cells in detection and avoidance of aluminum toxicity. Plant Physiol. 2001, 125, 1978–1987. [Google Scholar] [CrossRef]
- Larsen, F.H.; Byg, I.; Damager, I.; Diaz, J.; Engelsen, S.B.; Ulvskov, P. Residue specific hydration of primary cell wall potato pectin identified by solid-state 13C single-pulse MAS and CP/MAS NMR spectroscopy. Biomacromolecules 2011, 12, 1844–1850. [Google Scholar] [CrossRef]
- Willats, W.G.; Steele-King, C.G.; Marcus, S.E.; Knox, J.P. Side chains of pectic polysaccharides are regulated in relation to cell proliferation and cell differentiation. Plant J. 1999, 20, 619–628. [Google Scholar] [CrossRef]
- Willats, W.FG.; Orfila, C.; Limberg, G.; Buchholt, H.C.; van Alebeek, G.J.W.M.; Voragen, A.G.J.; Marcus, S.E.; Christensen, T.M.I.E.; Mikkelsen, J.D.; Murray, B.S.; et al. Modulation of the degree and pattern of methyl-esterification of pectic homogalacturonan in plant cell walls—Implications for pectin methyl esterase action, matrix properties and cell adhesion. J. Biol. Chem. 2001, 276, 19404–19413. [Google Scholar] [CrossRef]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Verhoef, R.; Schols, H.A.; McCleary, B.V.; McKee, L.; Gilbert, H.J.; Knox, J.P. Developmental complexity of arabinan polysaccharides and their processing in plant cell walls. Plant J. 2009, 59, 413–425. [Google Scholar] [CrossRef]
- Micheli, F. Pectin methylesterases: Cell wall enzymes with important roles in plant physiology. Trends Plant Sci. 2001, 6, 414–419. [Google Scholar] [CrossRef]
- Wolf, S.; Mouille, G.; Pelloux, J. Homogalacturonan methyl-esterification and plant development. Mol. Plant 2009, 5, 851–860. [Google Scholar] [CrossRef]
- Wu, H.-Ch.; Bulgakov, V.P.; Jinn, T.L. Pectin Methylesterases: Cell Wall Remodeling Proteins Are Required for Plant Response to Heat Stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [Green Version]
- Carpita, N.; McCann, M. The Cell Wall; Buchanan, B., Gruissem, W., Jones, R., Buchanan, B., Gruissem, W., Jones, R., Eds.; Biochemistry and Molecular Biology of Plants, American Society of Plant Physiologists: Rockwell, MD, USA, 2000; pp. 52–108. [Google Scholar]
- Ryden, P.; Sugimoto-Shirasu, K.; Smith, A.C.; Findlay, K.; Reiter, W.-D.; McCann, M. Tensile properties of Arabidopsis cell walls depend on both a xylogucan cross-linked microfibrillar network and rhamnogalacturonan II-borate complexes. Plant Physiol. 2003, 132, 1033–1040. [Google Scholar] [CrossRef]
- Steedman, H.F. Polyester wax; a new ribboning embedding medium for histology. Nature 1957, 179, 1345. [Google Scholar] [CrossRef]
- Jones, L.; Seymour, G.B.; Knox, J.P. Localization of pectic galactan in tomato cell walls using a monoclonal antibody specific to (1-4)-β-D-Galactan. Plant Physiol. 1997, 113, 1405–1412. [Google Scholar] [CrossRef]
- Willats, W.G.; Marcus, S.E.; Knox, J.P. Generation of monoclonal antibody specific to (1—>5)-alphaL-arabinan. Carbohydr. Res. 1998, 308, 149–152. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaskowiak, J.; Kwasniewska, J.; Milewska-Hendel, A.; Kurczynska, E.U.; Szurman-Zubrzycka, M.; Szarejko, I. Aluminum Alters the Histology and Pectin Cell Wall Composition of Barley Roots. Int. J. Mol. Sci. 2019, 20, 3039. https://doi.org/10.3390/ijms20123039
Jaskowiak J, Kwasniewska J, Milewska-Hendel A, Kurczynska EU, Szurman-Zubrzycka M, Szarejko I. Aluminum Alters the Histology and Pectin Cell Wall Composition of Barley Roots. International Journal of Molecular Sciences. 2019; 20(12):3039. https://doi.org/10.3390/ijms20123039
Chicago/Turabian StyleJaskowiak, Joanna, Jolanta Kwasniewska, Anna Milewska-Hendel, Ewa Urszula Kurczynska, Miriam Szurman-Zubrzycka, and Iwona Szarejko. 2019. "Aluminum Alters the Histology and Pectin Cell Wall Composition of Barley Roots" International Journal of Molecular Sciences 20, no. 12: 3039. https://doi.org/10.3390/ijms20123039