Fine Mapping of Glutamate Decarboxylase 65 Epitopes Reveals Dependency on Hydrophobic Amino Acids for Specific Interactions

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Screening of Overlapping Peptides Identifying Antigenic Regions
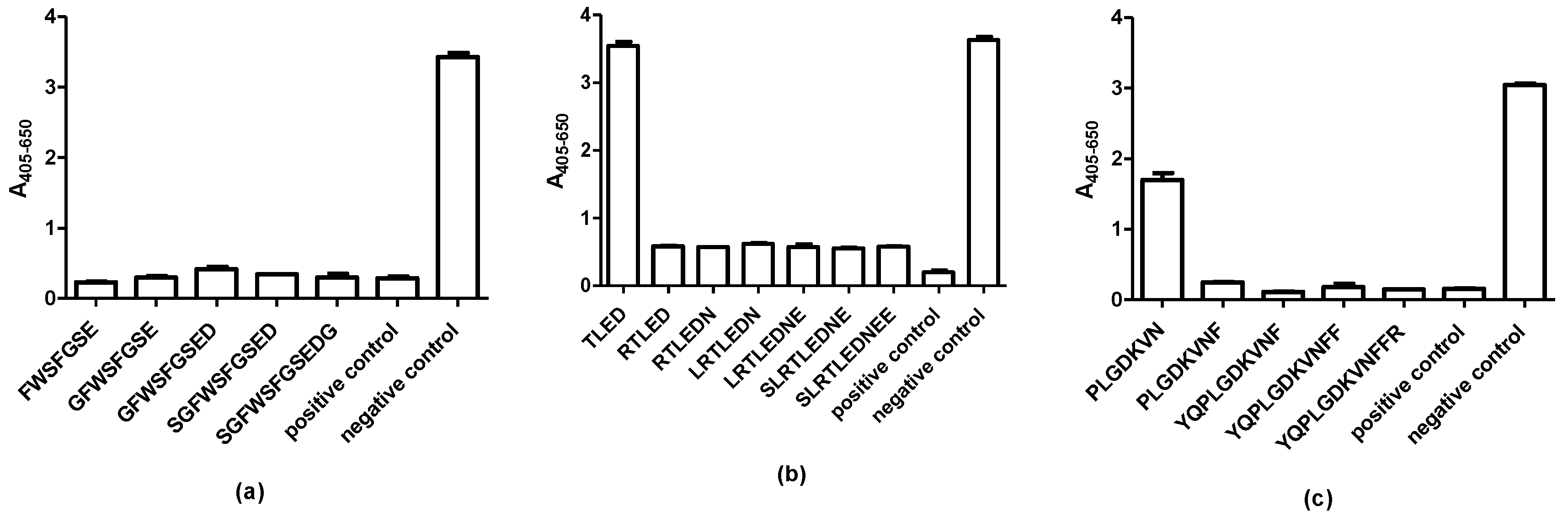
2.2. Screening of N-Terminally and C-Terminally Truncated Peptides
2.3. Final Identification of Complete Epitopes
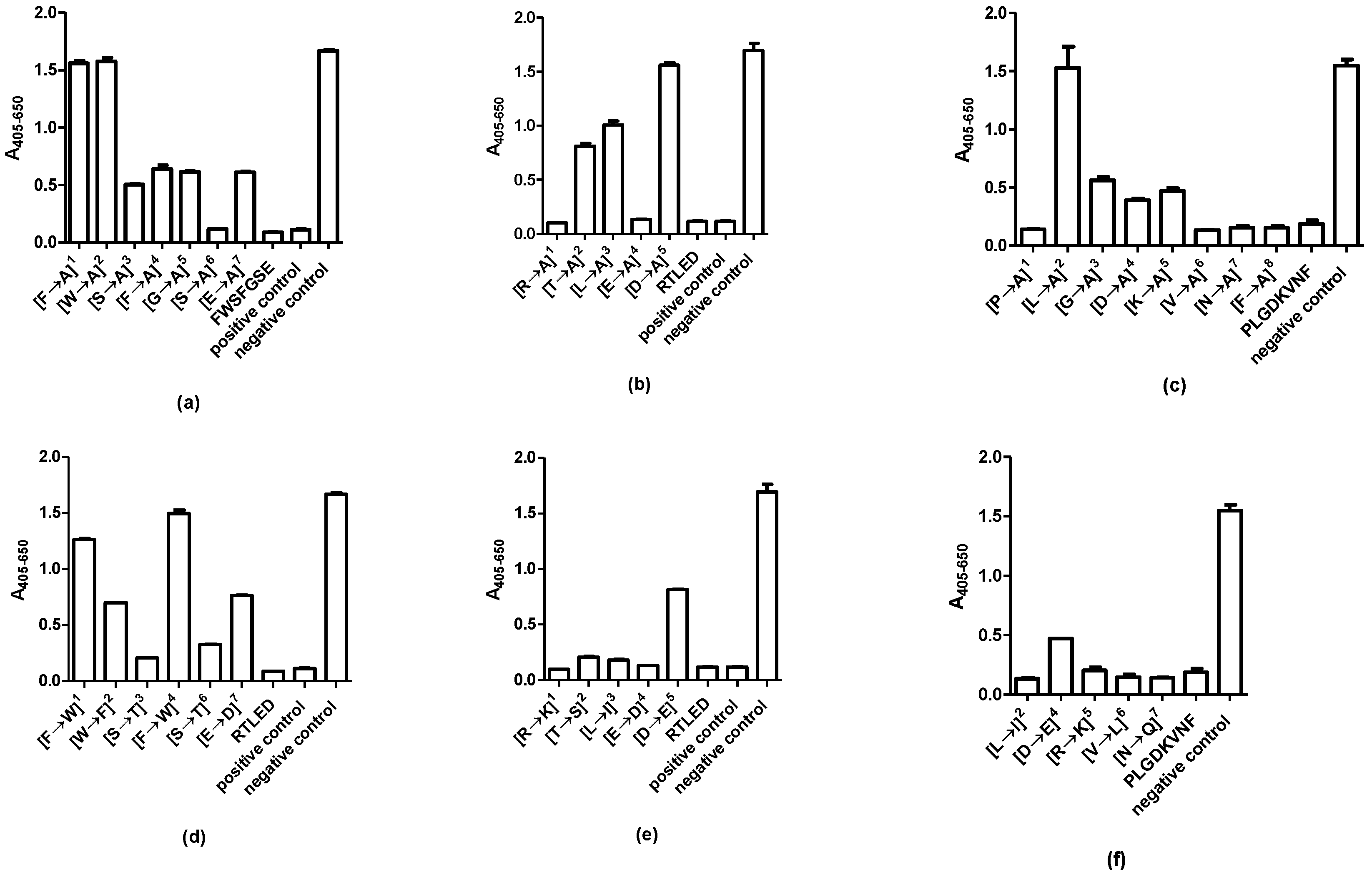
2.4. Identification of Essential Amino Acids
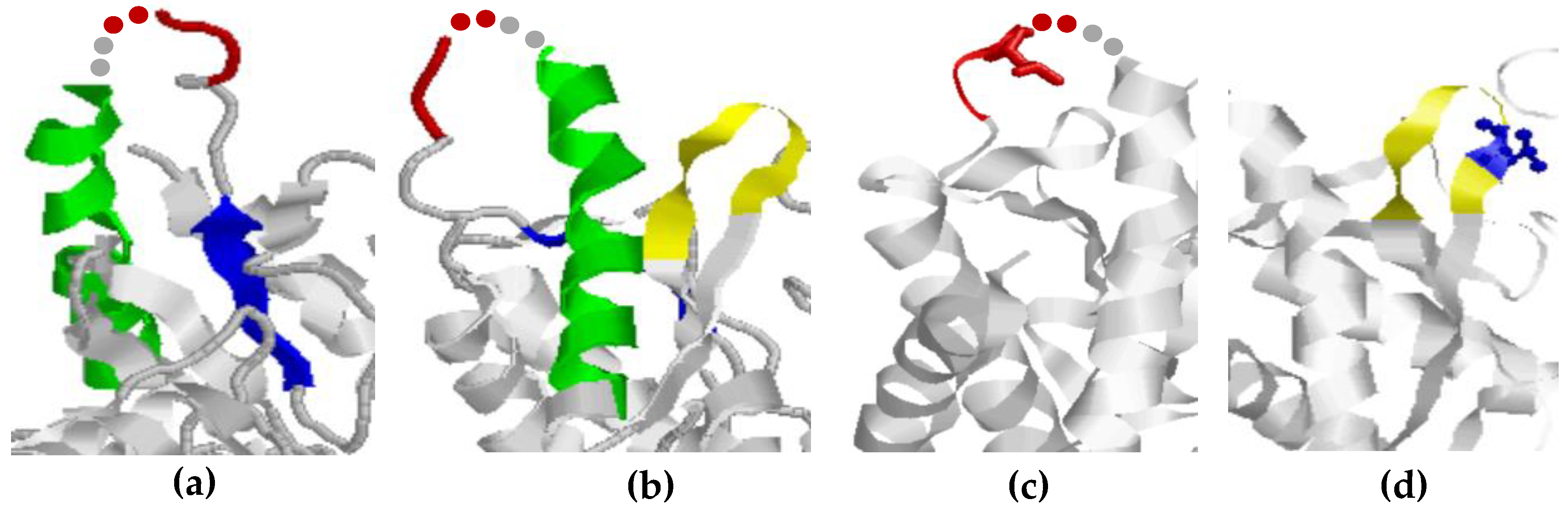
2.5. Epitope Structures
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Peptides
4.3. Modified Enzyme-Linked Immunosorbent Assay
4.4. Competitive Inhibition Assay
4.5. Structural Analysis of GAD65 Epitopes
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| aa | Amino acid |
| AP | Alkaline phosphatase |
| ELISA | Enzyme-linked immunosorbent assay |
| GABA | Γ-aminobutyric acid |
| GAD | Glutamate decarboxylase |
| PLP | Pyridoxal phosphate |
| pNPP | Para-nitrophenylphosphate |
| T1D | Type 1 diabetes |
| TTN | Tris-Tween-NaCl |
| SPS | Stiff-person syndrome |
References
- Rubinstein, N.D.; Mayrose, I.; Halperin, D.; Yekutieli, D.; Gershoni, J.M.; Pupko, T. Computational characterization of b-cell epitopes. Mol. Immunol. 2008, 45, 3477–3489. [Google Scholar] [CrossRef] [PubMed]
- Agca, S.; Houen, G.; Trier, N.H. Characterization of continuous b-cell epitopes in the n-terminus of glutamate decarboxylase67 using monoclonal antibodies. J. Pept. Sci. 2014, 20, 928–934. [Google Scholar] [CrossRef]
- Dam, C.E.; Houen, G.; Hansen, P.R.; Trier, N.H. Identification and fine mapping of a linear b cell epitope of human vimentin. Scand. J. Clin. Lab. Investig. 2014, 74, 506–514. [Google Scholar] [CrossRef]
- Haste Andersen, P.; Nielsen, M.; Lund, O. Prediction of residues in discontinuous b-cell epitopes using protein 3D structures. Protein Sci. 2006, 15, 2558–2567. [Google Scholar] [CrossRef] [PubMed]
- Trier, N.H. Characterization of peptide antibodies by epitope mapping using resin-bound and soluble peptides. Methods Mol. Biol. 2015, 1348, 229–239. [Google Scholar]
- Tronstrom, J.; Draborg, A.H.; Hansen, P.R.; Houen, G.; Trier, N.H. Identification of a linear epitope recognized by a monoclonal antibody directed to the heterogeneous nucleoriboprotein A2. Protein Pept. Lett. 2014, 21, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Petersen, N.H.; Hansen, P.R.; Houen, G. Fast and efficient characterization of an anti-gliadin monoclonal antibody epitope related to celiac disease using resin-bound peptides. J. Immunol. Methods 2011, 365, 174–182. [Google Scholar] [CrossRef]
- Trier, N.H.; Dam, C.E.; Olsen, D.T.; Hansen, P.R.; Houen, G. Contribution of peptide backbone to anti-citrullinated peptide antibody reactivity. PLoS ONE 2015, 10, e0144707. [Google Scholar] [CrossRef]
- Trier, N.H.; Leth, M.L.; Hansen, P.R.; Houen, G. Cross-reactivity of a human IgG1 anticitrullinated fibrinogen monoclonal antibody to a citrullinated profilaggrin peptide. Protein Sci. 2012, 21, 1929–1941. [Google Scholar] [CrossRef]
- Amrutkar, S.D.; Trier, N.H.; Hansen, P.R.; Houen, G. Fine mapping of a monoclonal antibody to the n-methyl d-aspartate receptor reveals a short linear epitope. Biopolymers 2012, 98, 567–575. [Google Scholar] [CrossRef]
- Behan, K.A.; Johnston, P.G.; Allegra, C.J. Epitope mapping of a series of human thymidylate synthase monoclonal antibodies. Cancer Res. 1998, 58, 2606–2611. [Google Scholar]
- Thornton, J.M.; Edwards, M.S.; Taylor, W.R.; Barlow, D.J. Location of ‘continuous’ antigenic determinants in the protruding regions of proteins. EMBO J. 1986, 5, 409–413. [Google Scholar] [CrossRef]
- Trier, N.H.; Hansen, P.R.; Vedeler, C.A.; Somnier, F.E.; Houen, G. Identification of continuous epitopes of hud antibodies related to paraneoplastic diseases/small cell lung cancer. J. Neuroimmunol. 2012, 243, 25–33. [Google Scholar] [CrossRef]
- Ofran, Y.; Schlessinger, A.; Rost, B. Automated identification of complementarity determining regions (cdrs) reveals peculiar characteristics of cdrs and b cell epitopes. J. Immunol. 2008, 181, 6230–6235. [Google Scholar] [CrossRef]
- David, M.; Borza, D.B.; Leinonen, A.; Belmont, J.M.; Hudson, B.G. Hydrophobic amino acid residues are critical for the immunodominant epitope of the goodpasture autoantigen. A molecular basis for the cryptic nature of the epitope. J. Biol. Chem. 2001, 276, 6370–6377. [Google Scholar] [CrossRef] [PubMed]
- Kringelum, J.V.; Nielsen, M.; Padkjaer, S.B.; Lund, O. Structural analysis of b-cell epitopes in antibody: Protein complexes. Mol. Immunol. 2013, 53, 24–34. [Google Scholar] [CrossRef]
- Bogan, A.A.; Thorn, K.S. Anatomy of hot spots in protein interfaces. J. Mol. Biol. 1998, 280, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Uysal, H.; Bockermann, R.; Nandakumar, K.S.; Sehnert, B.; Bajtner, E.; Engstrom, A.; Serre, G.; Burkhardt, H.; Thunnissen, M.M.; Holmdahl, R. Structure and pathogenicity of antibodies specific for citrullinated collagen type ii in experimental arthritis. J. Exp. Med. 2009, 206, 449–462. [Google Scholar] [CrossRef]
- Welner, S.; Trier, N.H.; Houen, G.; Hansen, P.R. Identification and mapping of a linear epitope of centromere protein f using monoclonal antibodies. J. Pept. Sci. 2013, 19, 95–101. [Google Scholar] [CrossRef]
- Trier, N.H.; Holm, B.E.; Slot, O.; Locht, H.; Lindegaard, H.; Svendsen, A.; Houen, G. Physical characteristics of a citrullinated pro-filaggrin epitope recognized by anti-citrullinated protein antibodies in rheumatoid arthritis sera. PLoS ONE 2016, 11, e0168542. [Google Scholar] [CrossRef]
- Acharya, K.R.; Lloyd, M.D. The advantages and limitations of protein crystal structures. Trends Pharmacol. Sci. 2005, 26, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Duquerroy, S.; Stura, E.A.; Bressanelli, S.; Fabiane, S.M.; Vaney, M.C.; Beale, D.; Hamon, M.; Casali, P.; Rey, F.A.; Sutton, B.J.; et al. Crystal structure of a human autoimmune complex between igm rheumatoid factor rf61 and igg1 fc reveals a novel epitope and evidence for affinity maturation. J. Mol. Biol. 2007, 368, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.F.; Erlander, M.G.; Hitz, B.C.; Tillakaratne, N.J.; Kaufman, D.L.; Wagner-McPherson, C.B.; Evans, G.A.; Tobin, A.J. Two human glutamate decarboxylases, 65-kDa GAD and 67-kDa GAD, are each encoded by a single gene. Proc. Natl. Acad. Sci. USA 1992, 89, 2115–2119. [Google Scholar] [CrossRef] [PubMed]
- Fenalti, G.; Law, R.H.; Buckle, A.M.; Langendorf, C.; Tuck, K.; Rosado, C.J.; Faux, N.G.; Mahmood, K.; Hampe, C.S.; Banga, J.P.; et al. Gaba production by glutamic acid decarboxylase is regulated by a dynamic catalytic loop. Nat. Struct. Mol. Biol. 2007, 14, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Towns, R.; Pietropaolo, M. GAD65 autoantibodies and its role as biomarker of type 1 diabetes and latent autoimmune diabetes in adults (LADA). Drugs Future 2011, 36, 847. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Rowley, M.; Jayakrishnan, B.; Teuber, S.; Gershwin, M.E.; Mackay, I.R. Stiff-person syndrome (SPS) and anti-GAD-related CNS degenerations: Protean additions to the autoimmune central neuropathies. J. Autoimmun. 2011, 37, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Jayakrishnan, B.; Hoke, D.E.; Langendorf, C.G.; Buckle, A.M.; Rowley, M.J. An analysis of the cross-reactivity of autoantibodies to GAD65 and GAD67 in diabetes. PLoS ONE 2011, 6, e18411. [Google Scholar] [CrossRef]
- Luo, D.; Gilliam, L.K.; Greenbaum, C.; Bekris, L.; Hampe, C.S.; Daniels, T.; Richter, W.; Marcovina, S.M.; Rolandsson, O.; Landin-Olsson, M.; et al. Conformation-dependent GAD65 autoantibodies in diabetes. Diabetologia 2004, 47, 1581–1591. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, B.; Schlosser, M.; Luhder, F.; Strebelow, M.; Augstein, P.; Northemann, W.; Powers, A.C.; Ziegler, M. Murine monoclonal glutamic acid decarboxylase (GAD)65 antibodies recognize autoimmune-associated GAD epitope regions targeted in patients with type 1 diabetes mellitus and stiff-man syndrome. Acta Diabetol. 1996, 33, 225–231. [Google Scholar] [CrossRef]
- Fenalti, G.; Buckle, A.M. Structural biology of the gad autoantigen. Autoimmun. Rev. 2010, 9, 148–152. [Google Scholar] [CrossRef]
- Ronkainen, M.S.; Savola, K.; Knip, M. Antibodies to GAD65 epitopes at diagnosis and over the first 10 years of clinical type 1 diabetes mellitus. Scand. J. Immunol. 2004, 59, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Powers, A.C.; Bavik, K.; Tremble, J.; Daw, K.; Scherbaum, W.A.; Banga, J.P. Comparative analysis of epitope recognition of glutamic acid decarboxylase (GAD) by autoantibodies from different autoimmune disorders. Clin. Exp. Immunol. 1999, 118, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Tanaka, S.; Okubo, M.; Nakanishi, K.; Murase, T.; Lernmark, A. Unique epitopes of glutamic acid decarboxylase autoantibodies in slowly progressive type 1 diabetes. J. Clin. Endocrinol. Metab. 2003, 88, 4768–4775. [Google Scholar] [CrossRef] [PubMed]
- Trier, N.H.; Hansen, P.R.; Houen, G. Production and characterization of peptide antibodies. Methods 2012, 56, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, J. Mining for the antibody-antigen interacting associations that predict the b cell epitopes. BMC Struct. Biol. 2010, 10 (Suppl. 1), S6. [Google Scholar] [CrossRef]
- Kiener, T.K.; Jia, Q.; Lim, X.F.; He, F.; Meng, T.; Chow, V.T.; Kwang, J. Characterization and specificity of the linear epitope of the enterovirus 71 VP2 protein. Virol. J. 2012, 9, 55. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H. What is a b-cell epitope? Methods Mol. Biol. 2009, 524, 3–20. [Google Scholar] [PubMed]
- Schulz, G.E.; Schirmer, R.H. Principles of Protein Structure; Springer-Verlag: New York, NY, USA, 1979; p. 28. [Google Scholar]
- Liang, S.; Zheng, D.; Standley, D.M.; Yao, B.; Zacharias, M.; Zhang, C. Epsvr and epmeta: Prediction of antigenic epitopes using support vector regression and multiple server results. BMC Bioinforma. 2010, 11, 381. [Google Scholar] [CrossRef]
- Neuvirth, H.; Raz, R.; Schreiber, G. Promate: A structure based prediction program to identify the location of protein-protein binding sites. J. Mol. Biol. 2004, 338, 181–199. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdarnini, N.; Holm, B.; Hansen, P.; Rovero, P.; Houen, G.; Trier, N. Fine Mapping of Glutamate Decarboxylase 65 Epitopes Reveals Dependency on Hydrophobic Amino Acids for Specific Interactions. Int. J. Mol. Sci. 2019, 20, 2909. https://doi.org/10.3390/ijms20122909
Valdarnini N, Holm B, Hansen P, Rovero P, Houen G, Trier N. Fine Mapping of Glutamate Decarboxylase 65 Epitopes Reveals Dependency on Hydrophobic Amino Acids for Specific Interactions. International Journal of Molecular Sciences. 2019; 20(12):2909. https://doi.org/10.3390/ijms20122909
Chicago/Turabian StyleValdarnini, Niccolò, Bettina Holm, Paul Hansen, Paolo Rovero, Gunnar Houen, and Nicole Trier. 2019. "Fine Mapping of Glutamate Decarboxylase 65 Epitopes Reveals Dependency on Hydrophobic Amino Acids for Specific Interactions" International Journal of Molecular Sciences 20, no. 12: 2909. https://doi.org/10.3390/ijms20122909